
Mechanical Stretch Activates TRPV4 and Hemichannel Responses in the Nonpigmented Ciliary Epithelium


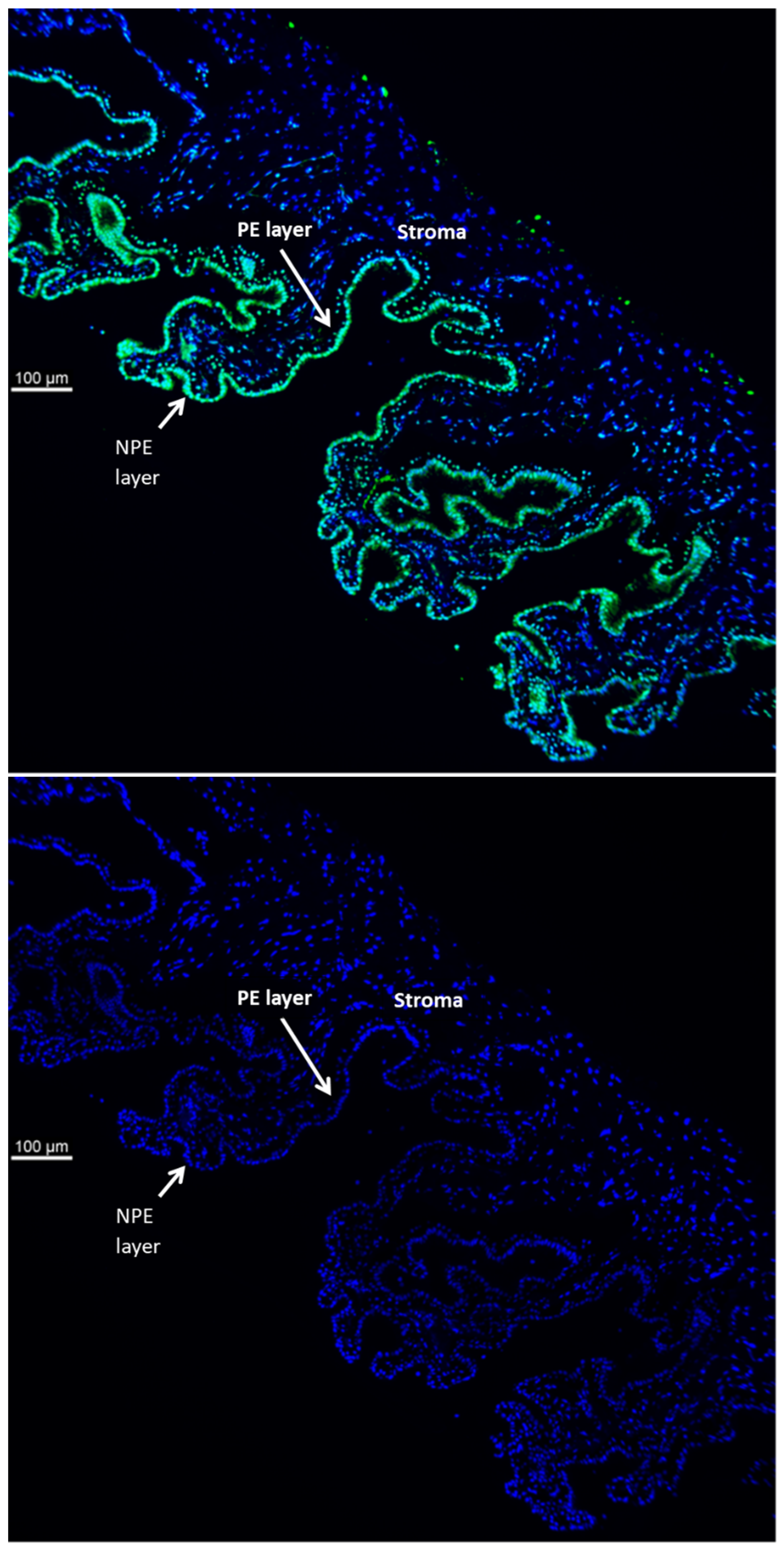{kind=link}
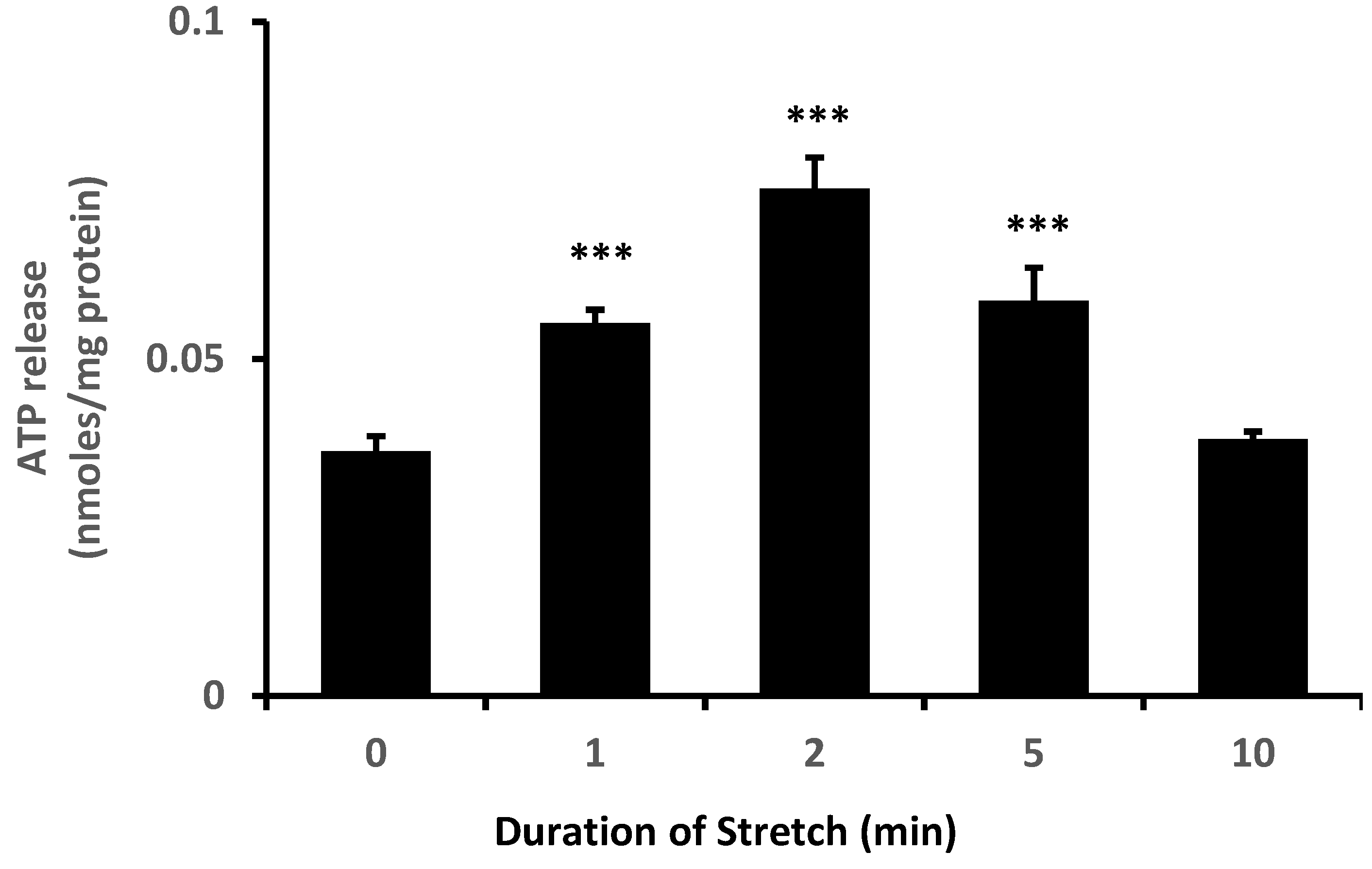{kind=link}
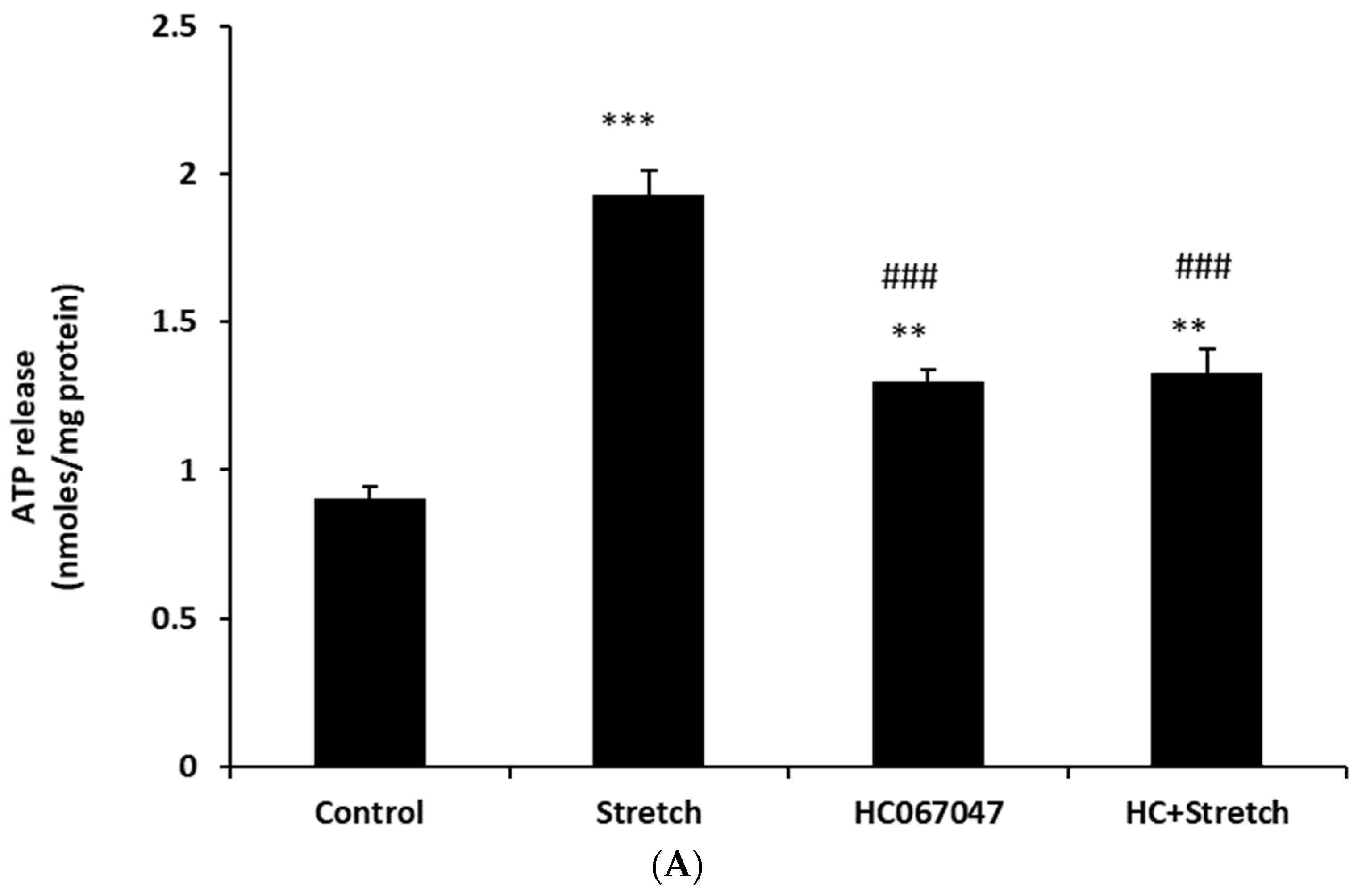{kind=link}
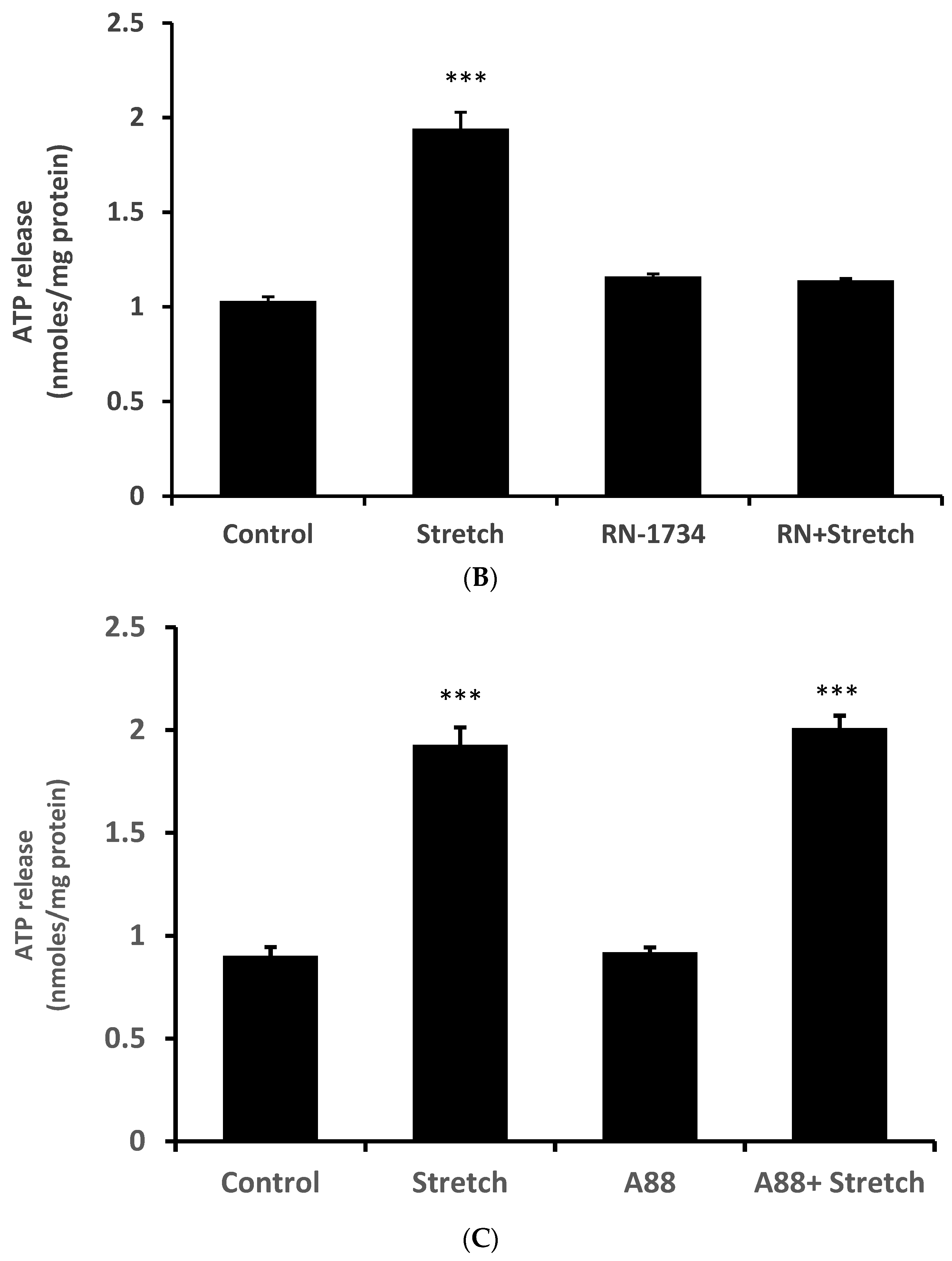{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
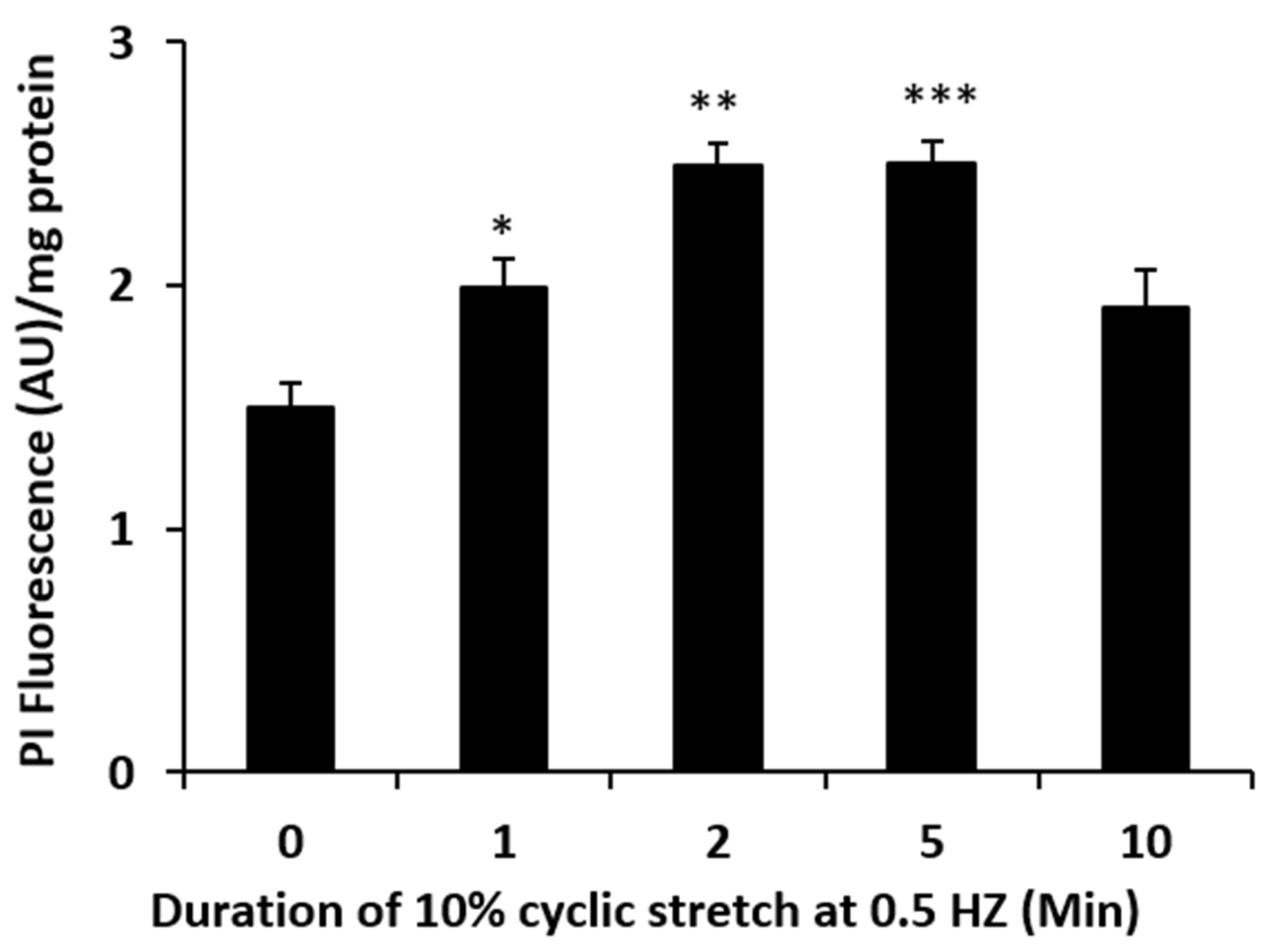
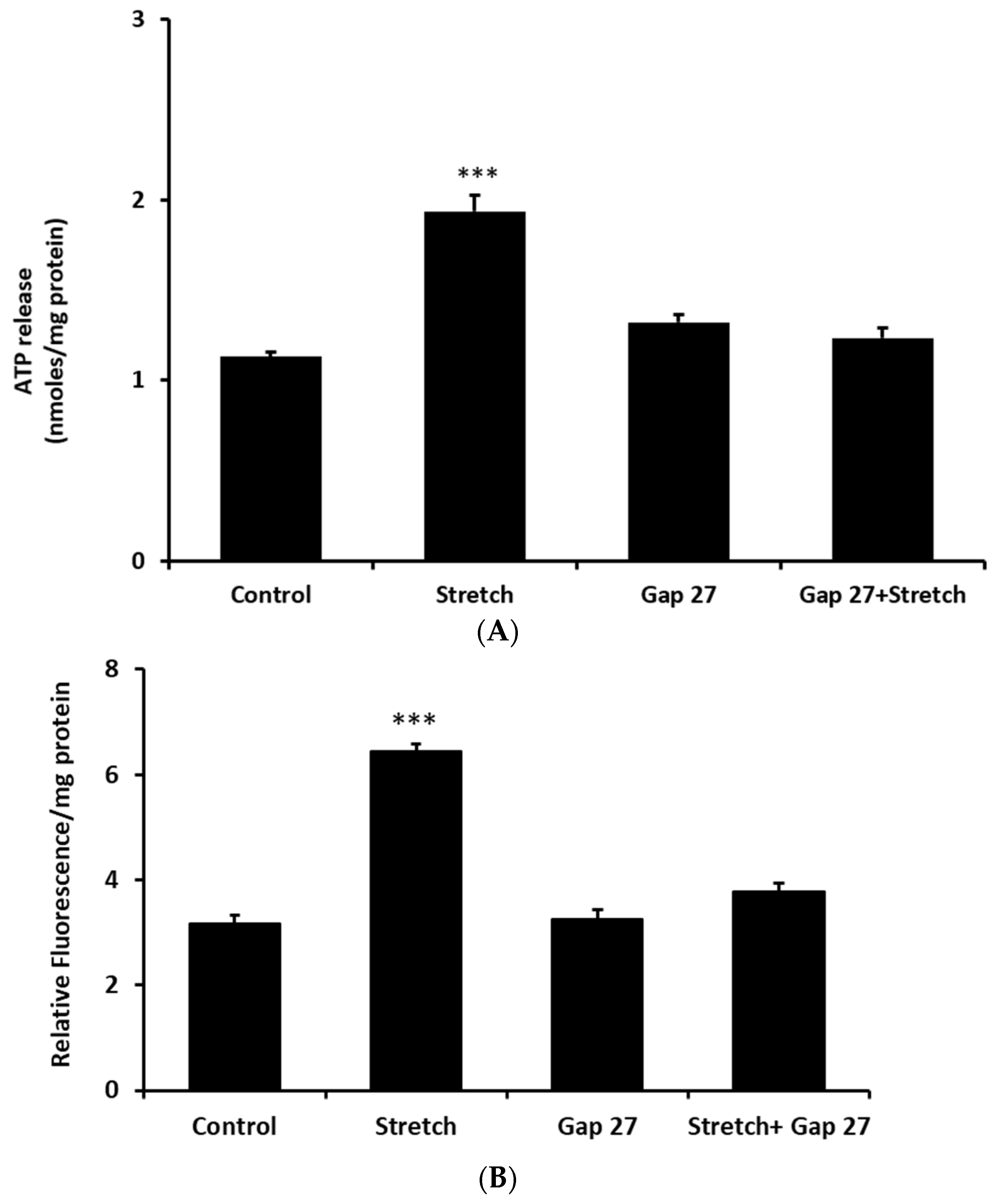
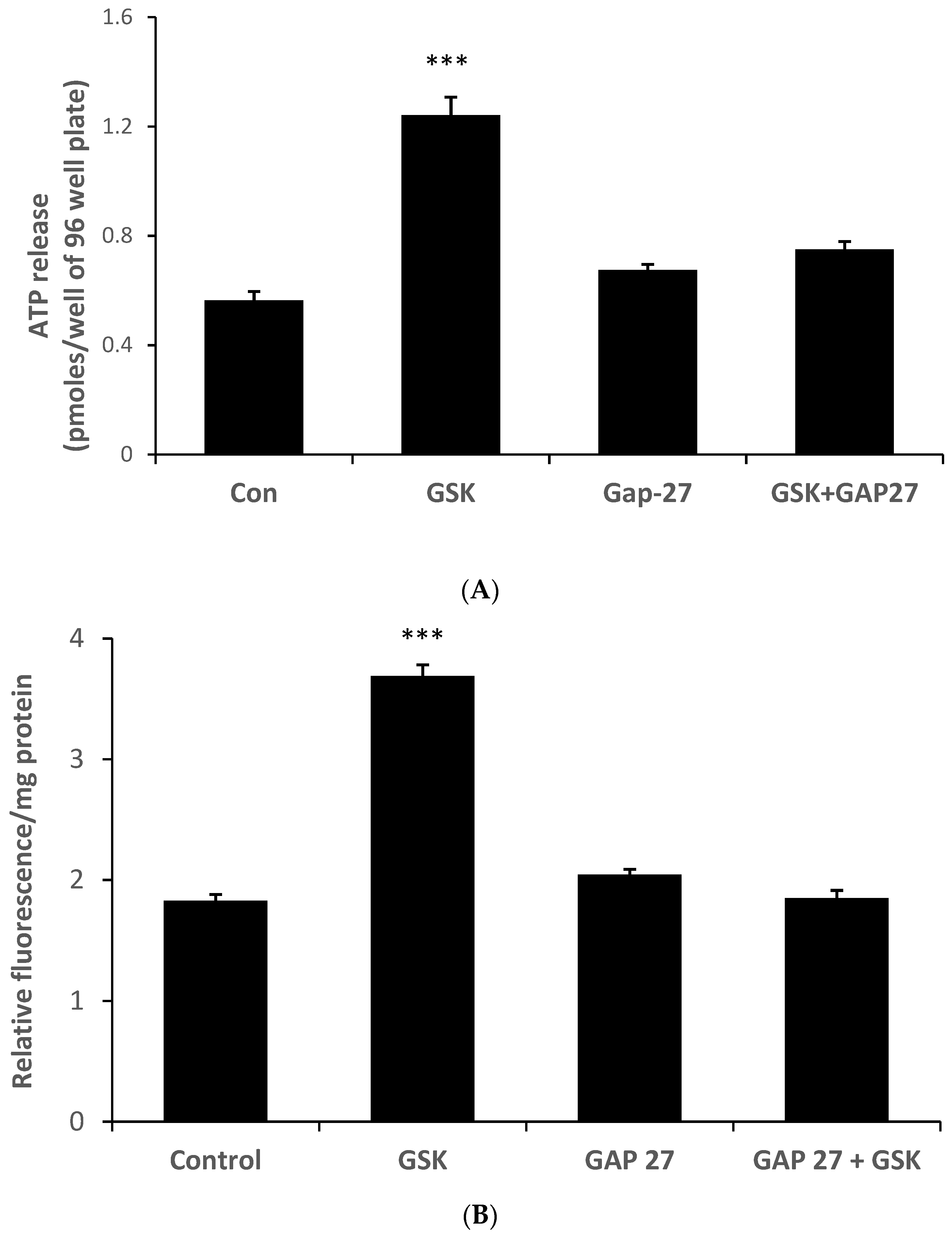
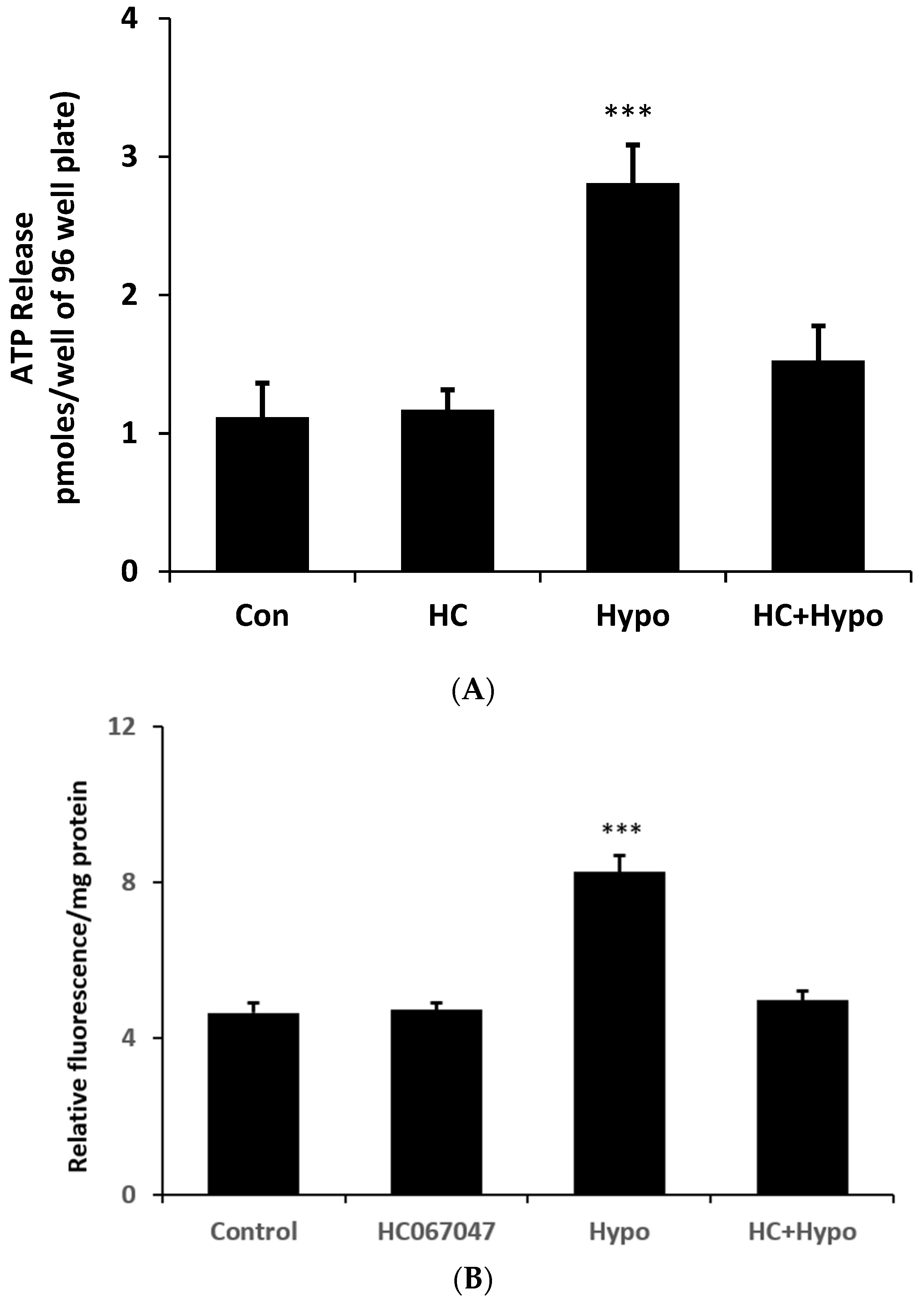
2. Results
3. Discussion
4. Methods
4.1. Reagents and Antibodies
4.2. Immunolocalization
4.3. Cell Culture
4.4. Stretch Stimulus
4.5. Measurement of ATP
4.6. Propidium Iodide (PI) Uptake
4.7. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Delamere, N.A. Ciliary Body and Ciliary Epithelium. Adv. Organ. Biol. 2005, 10, 127–148. [Google Scholar]
- Ghosh, S.; Hernando, N.; Martín-Alonso, J.M.; Martin-Vasallo, P.; Coca-Prados, M. Expression of multiple Na+, K(+)-ATPase genes reveals a gradient of isoforms along the nonpigmented ciliary epithelium: Functional implications in aqueous humor secretion. J. Cell Physiol. 1991, 149, 184–194. [Google Scholar] [CrossRef] [PubMed]
- Riley, M.V.; Kishida, K. ATPases of ciliary epithelium: Cellular and subcellular distribution and probable role in secretion of aqueous humor. Exp. Eye Res. 1986, 42, 559–568. [Google Scholar] [CrossRef] [PubMed]
- Usukura, J.; Fain, G.L.; Bok, D. [3H]ouabain localization of Na-K ATPase in the epithelium of rabbit ciliary body pars plicata. Invest. Ophthalmol. Vis. Sci. 1988, 29, 606–614. [Google Scholar]
- Civan, M.M.; Macknight, A.D. The ins and outs of aqueous humour secretion. Exp. Eye Res. 2004, 78, 625–631. [Google Scholar] [CrossRef]
- Freddo, T.; Gong, H. Anatomy of the Ciliary Body and Outflow Pathways. Duanes Clin. Ophthalmol. William Tasman. Ed. Lipincott Williams Wilkins 2007, 3, 1–18. [Google Scholar]
- Freddo, T.F. A contemporary concept of the blood-aqueous barrier. Prog. Retin. Eye Res. 2013, 32, 181–195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brubaker, R.F. Flow of aqueous humor in humans The Friedenwald Lecture. Investig. Ophthalmol. Vis. Sci. 1991, 32, 3145–3166. [Google Scholar]
- Shahidullah, M.; Delamere, N.A. Connexins form functional hemichannels in porcine ciliary epithelium. Exp. Eye Res. 2014, 118, 20–29. [Google Scholar] [CrossRef] [Green Version]
- Delamere, N.A.; Shahidullah, M. Ion Transport Regulation by TRPV4 and TRPV1 in Lens and Ciliary Epithelium. Front. Physiol. 2021, 12, 834916. [Google Scholar] [CrossRef]
- Shahidullah, M.; Mandal, A.; Delamere, N.A. TRPV4 in porcine lens epithelium regulates hemichannel-mediated ATP release and Na-K-ATPase activity. Am. J. Physiol. Cell Physiol. 2012, 302, C1751–C1761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toft-Bertelsen, T.L.; MacAulay, N. TRPing on Cell Swelling—TRPV4 Senses It. Front. Immunol. 2021, 12, 730982. [Google Scholar] [CrossRef] [PubMed]
- Vincent, F.; Acevedo, A.; Nguyen, M.T.; Dourado, M.; DeFalco, J.; Gustafson, A.; Spiro, P.; Emerling, D.E.; Kelly, M.G.; Duncton, M.A. Identification and characterization of novel TRPV4 modulators. Biochem. Biophys. Res. Commun. 2009, 389, 490–494. [Google Scholar] [CrossRef]
- McGaraughty, S.; Segreti, J.A.; Fryer, R.M.; Brown, B.S.; Faltynek, C.R.; Kym, P.R. Antagonism of TRPV1 receptors indirectly modulates activity of thermoregulatory neurons in the medial preoptic area of rats. Brain Res. 2009, 1268, 58–67. [Google Scholar] [CrossRef]
- Taruno, A. ATP Release Channels. Int. J. Mol. Sci. 2018, 19, 808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Figueroa, V.A.; Jara, O.; Oliva, C.A.; Ezquer, M.; Ezquer, F.; Retamal, M.A.; Martínez, A.D.; Altenberg, G.A.; Vargas, A.A. Contribution of Connexin Hemichannels to the Decreases in Cell Viability Induced by Linoleic Acid in the Human Lens Epithelial Cells (HLE-B3). Front. Physiol. 2020, 10, 1574. [Google Scholar] [CrossRef] [Green Version]
- Rosenbaum, T.; Benítez-Angeles, M.; Sánchez-Hernández, R.; Morales-Lázaro, S.L.; Hiriart, M.; Morales-Buenrostro, L.E.; Torres-Quiroz, F. TRPV4: A Physio and Pathophysiologically Significant Ion Channel. Int. J. Mol. Sci. 2020, 21, 3837. [Google Scholar] [CrossRef]
- White, J.P.; Cibelli, M.; Urban, L.; Nilius, B.; McGeown, J.G.; Nagy, I. TRPV4: Molecular Conductor of a Diverse Orchestra. Physiol. Rev. 2016, 96, 911–973. [Google Scholar] [CrossRef] [Green Version]
- Delamere, N.A.; Shahidullah, M.; Mathias, R.T.; Gao, J.; Sun, X.; Sellitto, C.; White, T.W. Signaling Between TRPV1/TRPV4 and Intracellular Hydrostatic Pressure in the Mouse Lens. Invest. Ophthalmol. Vis. Sci. 2020, 61, 58. [Google Scholar] [CrossRef]
- Shahidullah, M.; Mandal, A.; Delamere, N.A. Activation of TRPV1 channels leads to stimulation of NKCC1 cotransport in the lens. Am. J. Physiol. Cell Physiol. 2018, 315, C793–C802. [Google Scholar] [CrossRef]
- Shahidullah, M.; Mandal, A.; Mathias, R.T.; Gao, J.; Križaj, D.; Redmon, S.; Delamere, N.A. TRPV1 activation stimulates NKCC1 and increases hydrostatic pressure in the mouse lens. Am. J. Physiol. Cell Physiol. 2020, 318, C969–C980. [Google Scholar] [CrossRef]
- Grygorczyk, R.; Hanrahan, J.W. CFTR-independent ATP release from epithelial cells triggered by mechanical stimuli. Am. J. Physiol. 1997, 272 Pt 1, C1058–C1066. [Google Scholar] [CrossRef]
- Burnstock, G. Introduction to Purinergic Signaling. Methods Mol. Biol. 2020, 2041, 1–15. [Google Scholar] [PubMed]
- Sanderson, J.; Dartt, D.A.; Trinkaus-Randall, V.; Pintor, J.; Civan, M.M.; Delamere, N.A.; Fletcher, E.L.; Salt, T.E.; Grosche, A.; Mitchell, C.H. Purines in the eye: Recent evidence for the physiological and pathological role of purines in the RPE, retinal neurons, astrocytes, Müller cells, lens, trabecular meshwork, cornea and lacrimal gland. Exp. Eye Res. 2014, 127, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Mitchell, C.H.; Carré, D.A.; McGlinn, A.M.; Stone, R.A.; Civan, M.M. A release mechanism for stored ATP in ocular ciliary epithelial cells. Proc. Natl. Acad. Sci. USA 1998, 95, 7174–7178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edelman, J.L.; Sachs, G.; Adorante, J.S. Ion transport asymmetry and functional coupling in bovine pigmented and nonpigmented ciliary epithelial cells. Am. J. Physiol. 1994, 266 Pt 1, C1210–C1221. [Google Scholar] [CrossRef]
- Edelman, J.L.; Loo, D.D.; Sachs, G. Characterization of potassium and chloride channels in the basolateral membrane of bovine nonpigmented ciliary epithelial cells. Investig. Ophthalmol. Vis. Sci. 1995, 36, 2706–2716. [Google Scholar]
- Walker, V.E.; Stelling, J.W.; Miley, H.E.; Jacob, T.J. Effect of coupling on volume-regulatory response of ciliary epithelial cells suggests mechanism for secretion. Am. J. Physiol. 1999, 276, C1432–C1438. [Google Scholar] [CrossRef]
- Martin-Vasallo, P.; Ghosh, S.; Coca-Prados, M. Expression of Na, K-ATPase alpha subunit isoforms in the human ciliary body and cultured ciliary epithelial cells. J. Cell Physiol. 1989, 141, 243–252. [Google Scholar] [CrossRef]
- Wetzel, R.K.; Sweadner, K.J. Immunocytochemical localization of NaK-ATPase isoforms in the rat and mouse ocular ciliary epithelium. Invest. Ophthalmol. Vis. Sci. 2001, 42, 763–769. [Google Scholar]
- Kiel, J.W.; Hollingsworth, M.; Rao, R.; Chen, M.; Reitsamer, H.A. Ciliary blood flow and aqueous humor production. Prog. Retin. Eye Res. 2011, 30, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahidullah, M.; Wilson, W.S. Atriopeptin, sodium azide and cyclic GMP reduce secretion of aqueous humour and inhibit intracellular calcium release in bovine cultured ciliary epithelium. Br. J. Pharmacol. 1999, 127, 1438–1446. [Google Scholar] [CrossRef] [Green Version]
- Shearer, T.W.; Crosson, C.E. Adenosine A1 receptor modulation of MMP-2 secretion by trabecular meshwork cells. Invest. Ophthalmol. Vis. Sci. 2002, 43, 3016–3020. [Google Scholar] [PubMed]
- Shahidullah, M.; To, C.-H.; Pelis, R.M.; Delamere, N.A. Studies on Bicarbonate Transporters and Carbonic Anhydrase in Porcine Nonpigmented Ciliary Epithelium. Invest. Ophthalmol. Vis. Sci. 2009, 50, 1791–1800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahidullah, M.; Rosales, J.L.; Delamere, N. Activation of Piezo1 Increases Na, K-ATPase-Mediated Ion Transport in Mouse Lens. Int. J. Mol. Sci. 2022, 23, 12870. [Google Scholar] [CrossRef]
- Shahidullah, M.; Tamiya, S.; Delamere, N.A. Primary culture of porcine nonpigmented ciliary epithelium. Curr. Eye Res. 2007, 32, 511–522. [Google Scholar] [CrossRef]
- Smith, P.K.; Krohn, R.I.; Hermanson, G.T.; Mallia, A.K.; Gartner, F.H.; Provenzano, M.D.; Fujimoto, E.K.; Goeke, N.M.; Olson, B.J.; Klenk, D.C. Measurement of protein using bicinchoninic acid. Anal. Biochem. 1985, 150, 76–85. [Google Scholar] [CrossRef] [PubMed]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shahidullah, M.; Delamere, N.A. Mechanical Stretch Activates TRPV4 and Hemichannel Responses in the Nonpigmented Ciliary Epithelium. Int. J. Mol. Sci. 2023, 24, 1673. https://doi.org/10.3390/ijms24021673
Shahidullah M, Delamere NA. Mechanical Stretch Activates TRPV4 and Hemichannel Responses in the Nonpigmented Ciliary Epithelium. International Journal of Molecular Sciences. 2023; 24(2):1673. https://doi.org/10.3390/ijms24021673
Chicago/Turabian StyleShahidullah, Mohammad, and Nicholas A. Delamere. 2023. "Mechanical Stretch Activates TRPV4 and Hemichannel Responses in the Nonpigmented Ciliary Epithelium" International Journal of Molecular Sciences 24, no. 2: 1673. https://doi.org/10.3390/ijms24021673
APA StyleShahidullah, M., & Delamere, N. A. (2023). Mechanical Stretch Activates TRPV4 and Hemichannel Responses in the Nonpigmented Ciliary Epithelium. International Journal of Molecular Sciences, 24(2), 1673. https://doi.org/10.3390/ijms24021673