
Upregulation of Peridinin-Chlorophyll A-Binding Protein in a Toxic Strain of Prorocentrum hoffmannianum under Normal and Phosphate-Depleted Conditions


Abstract
:1. Introduction
2. Results and Discussion
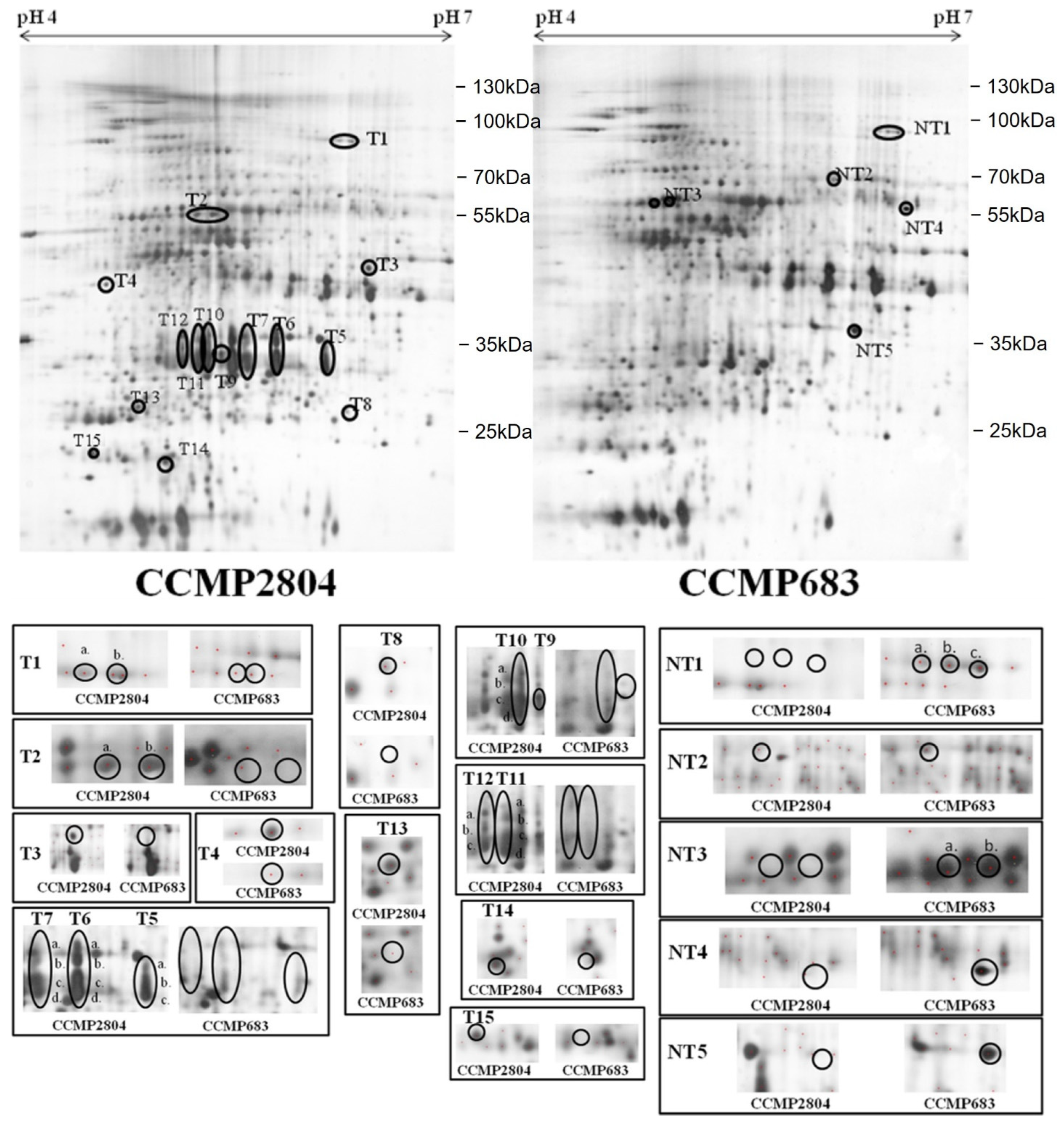
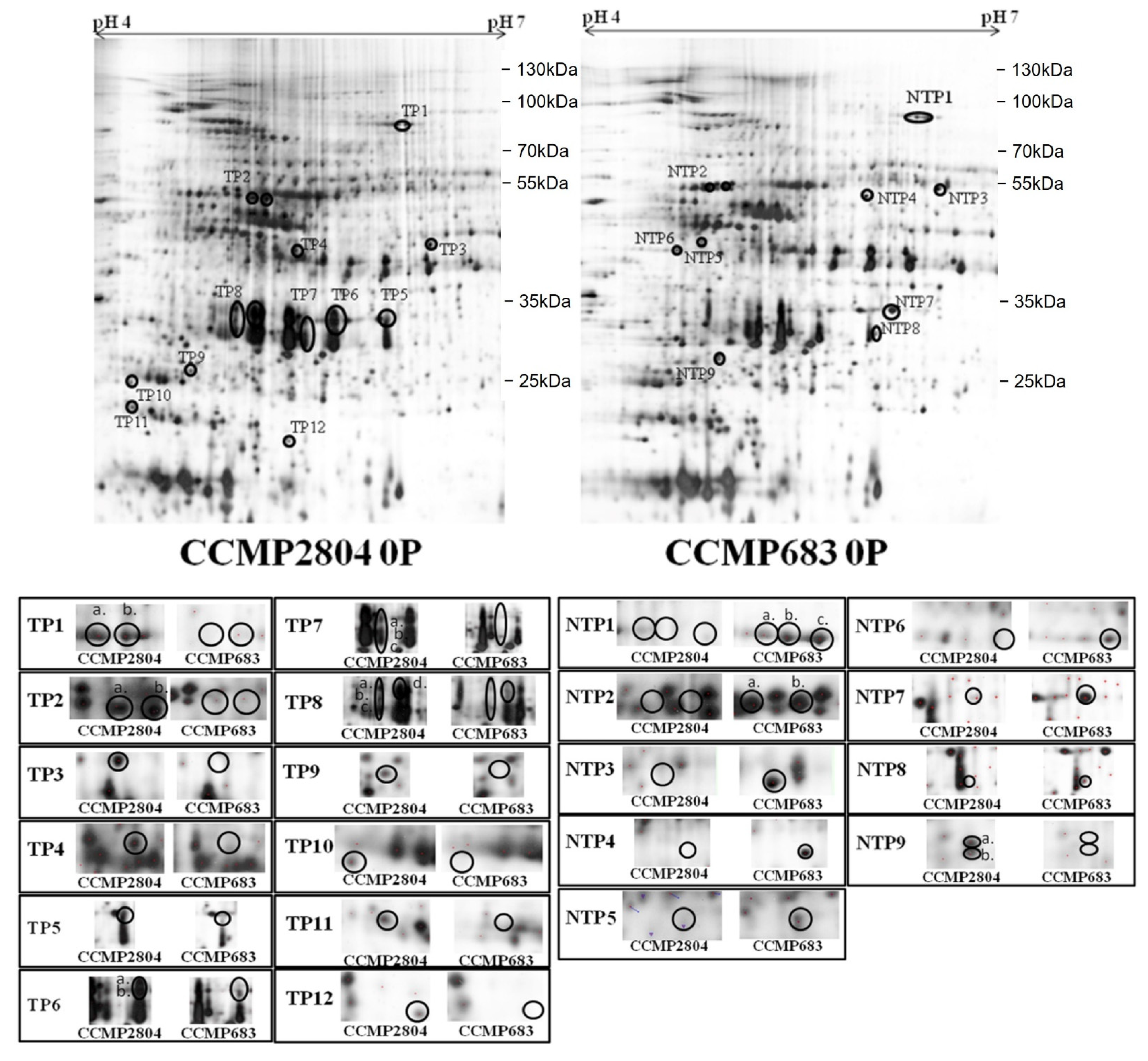
2.1. Differential Protein Expression between Toxic and Non-Toxic Strains
2.1.1. Normal Culture Condition
2.1.2. Phosphate Depletion
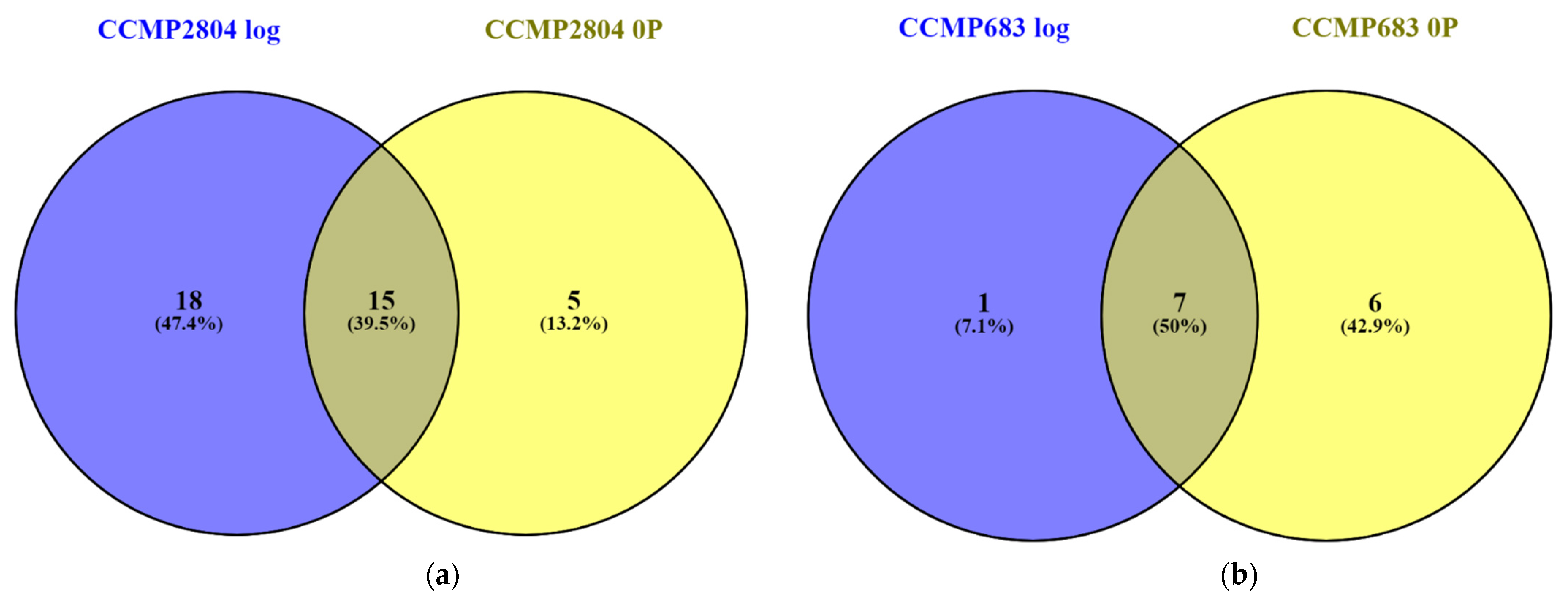
2.2. Shared Proteins between Both Strains during Log and under 0P
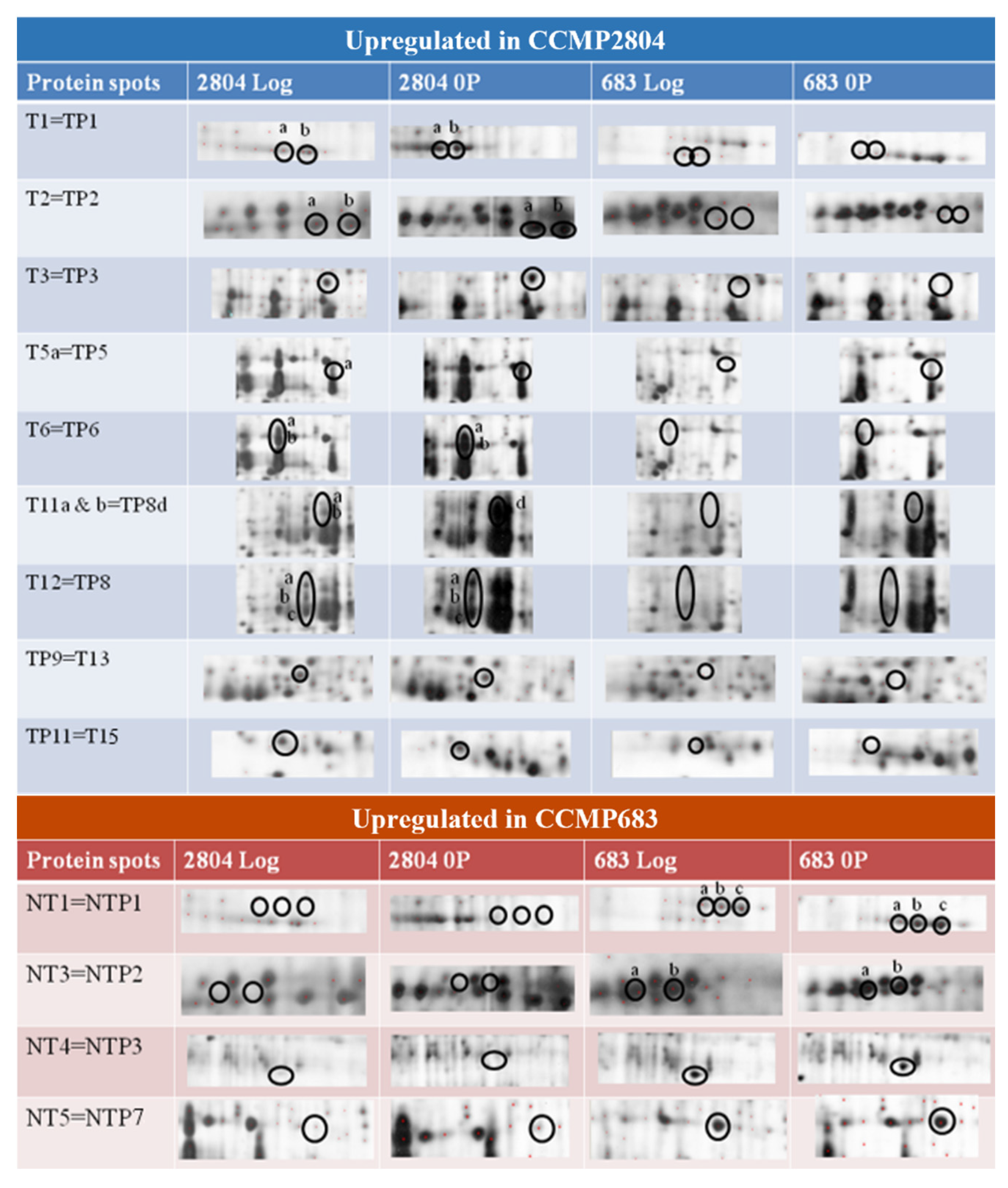
2.3. Identification of Differentially Expressed Proteins
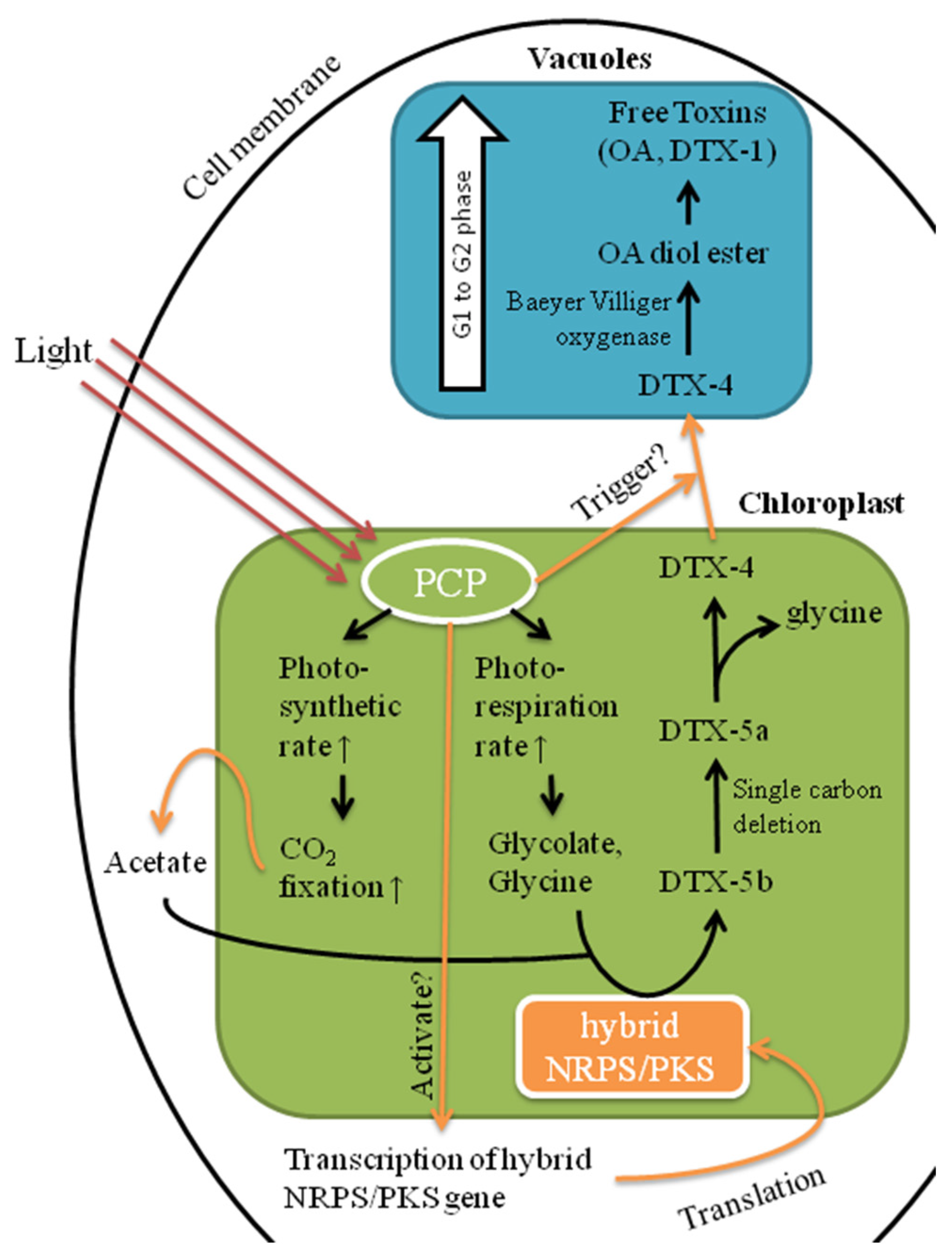
2.4. Possible Implications of Identified Proteins to DSP Toxin Production by P. hoffmannianum
3. Materials and Methods
3.1. Sources and Maintenance of Dinoflagellates
3.2. Cultivation Conditions and Cell Harvesting
3.3. Proteome Analysis
3.3.1. Protein Extraction and Quantification
3.3.2. Two-Dimensional Gel Electrophoresis and Image Analysis
3.3.3. In-Gel Digestion
3.3.4. MALDI-TOF Mass Spectrometry (MS) Analysis
3.3.5. De Novo Peptide Sequencing and N-Terminal Sulfonation
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Mok, J.-S.; Lee, T.-S.; Oh, E.-G.; Son, K.-T.; Hwang, H.-J.; Kim, J.-H. Stability of Domoic Acid at Different Temperature, pH and Light. Korean J. Fish. Aquat. Sci. 2009, 42, 8–14. [Google Scholar] [CrossRef] [Green Version]
- Indrasena, W. Storage stability of paralytic shellfish poisoning toxins. Food Chem. 2000, 71, 71–77. [Google Scholar] [CrossRef]
- Chen, T.; Xu, X.; Wei, J.; Chen, J.; Miu, R.; Huang, L.; Zhou, X.; Fu, Y.; Yan, R.; Wang, Z.; et al. Food-Borne Disease Outbreak of Diarrhetic Shellfish Poisoning Due to Toxic Mussel Consumption: The First Recorded Outbreak in China. PLoS ONE 2013, 8, e65049. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Altares, M.; Casanova, A.; Fernández-Tejedor, M.; Diogène, J.; de la Iglesia, P. Bloom of Dinophysis spp. dominated by D. sacculus and its related diarrhetic shellfish poisoning (DSP) outbreak in Alfacs Bay (Catalonia, NW Mediterranean Sea): Identification of DSP toxins in phytoplankton, shellfish and passive samplers. Reg. Stud. Mar. Sci. 2016, 6, 19–28. [Google Scholar] [CrossRef]
- Li, A.; Ma, J.; Cao, J.; McCarron, P. Toxins in mussels (Mytilus galloprovincialis) associated with diarrhetic shellfish poisoning episodes in China. Toxicon 2012, 60, 420–425. [Google Scholar] [CrossRef]
- Taylor, M.; McIntyre, L.; Ritson, M.; Stone, J.; Bronson, R.; Bitzikos, O.; Rourke, W.; Galanis, E.; Team, O.I. Outbreak of Diarrhetic Shellfish Poisoning Associated with Mussels, British Columbia, Canada. Mar. Drugs 2013, 11, 1669–1676. [Google Scholar] [CrossRef] [Green Version]
- Pitcher, G.C.; Krock, B.; Cembella, A.D. Accumulation of diarrhetic shellfish poisoning toxins in the oyster Crassostrea gigas and the mussel Choromytilus meridionalis in the southern Benguela ecosystem. Afr. J. Mar. Sci. 2011, 33, 273–281. [Google Scholar] [CrossRef] [Green Version]
- Yasumoto, T.; Oshima, Y.; Sugawara, W.; Fukuyo, Y.; Oguri, H.; Igarashi, T.; Fujita, N. Identification of Dinophysis fortii as the causative organism of diarrhetic shellfish poisoning. Nippon Suisan Gakkaishi 1980, 46, 1405–1411. [Google Scholar] [CrossRef] [Green Version]
- Eberhart, B.-T.L.; Moore, L.K.; Harrington, N.; Adams, N.G.; Borchert, J.; Trainer, V.L. Screening Tests for the Rapid Detection of Diarrhetic Shellfish Toxins in Washington State. Mar. Drugs 2013, 11, 3718–3734. [Google Scholar] [CrossRef] [Green Version]
- Hattenrath-Lehmann, T.K.; Lusty, M.W.; Wallace, R.B.; Haynes, B.; Wang, Z.; Broadwater, M.; Deeds, J.R.; Morton, S.L.; Hastback, W.; Porter, L.; et al. Evaluation of Rapid, Early Warning Approaches to Track Shellfish Toxins Associated with Dinophysis and Alexandrium Blooms. Mar. Drugs 2018, 16, 28. [Google Scholar] [CrossRef]
- Holmes, M.J.; Teo, S.L.M.; Khoo, H.W. Detection of diarrhetic shellfish poisoning toxins from tropical shellfish using liquid chromatography-selected reaction monitoring mass spectrometry. Nat. Toxins 1999, 7, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Honkanen, R.E.; Mowdy, D.E.; Dickey, R.W. Detection of DSP-Toxins, Okadaic Acid, and Dinophysis Toxin-1 in Shellfish by Serine/Threonine Protein Phosphatase Assay. J. AOAC Int. 1996, 79, 1336–1343. [Google Scholar] [CrossRef] [Green Version]
- Imai, I.; Sugioka, H.; Nishitani, G.; Mitsuya, T.; Hamano, Y. Monitoring of DSP toxins in small-sized plankton fraction of seawater collected in Mutsu Bay, Japan, by ELISA method: Relation with toxin contamination of scallop. Mar. Pollut. Bull. 2003, 47, 114–117. [Google Scholar] [CrossRef]
- Laycock, M.V.; Jellett, J.F.; Easy, D.J.; Donovan, M.A. First report of a new rapid assay for diarrhetic shellfish poisoning toxins. Harmful Algae 2006, 5, 74–78. [Google Scholar] [CrossRef]
- Mountfort, D.; Suzuki, T.; Truman, P. Protein phosphatase inhibition assay adapted for determination of total DSP in contaminated mussels. Toxicon 2001, 39, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Turrell, E.; Stobo, L. A comparison of the mouse bioassay with liquid chromatography–mass spectrometry for the detection of lipophilic toxins in shellfish from Scottish waters. Toxicon 2007, 50, 442–447. [Google Scholar] [CrossRef]
- Vale, P.; Botelho, M.J.; Rodrigues, S.M.; Gomes, S.S.; de Sampayo, M.A.M. Two decades of marine biotoxin monitoring in bivalves from Portugal (1986–2006): A review of exposure assessment. Harmful Algae 2008, 7, 11–25. [Google Scholar] [CrossRef]
- Daranas, A.H.; Fernández, J.J.; Norte, M.; Gavín, J.A.; Suárez-Gómez, B.; Souto, M.L. Biosynthetic studies of the DSP toxin skeleton. Chem. Rec. 2004, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Macpherson, G.R.; Burton, I.W.; LeBlanc, P.; Walter, J.A.; Wright, J.L.C. Studies of the Biosynthesis of DTX-5a and DTX-5b by the Dinoflagellate Prorocentrum maculosum: Regiospecificity of the Putative Baeyer−Villigerase and Insertion of a Single Amino Acid in a Polyketide Chain. J. Org. Chem. 2002, 68, 1659–1664. [Google Scholar] [CrossRef] [Green Version]
- Needham, J.; McLachlan, J.L.; Walter, J.A.; Wright, J.L.C. Biosynthetic origin of C-37 and C-38 in the polyether toxins okadaic acid and dinophysistoxin-1. J. Chem. Soc. Chem. Commun. 1994, 22, 2599–2600. [Google Scholar] [CrossRef]
- Norte, M.; Padilla, A.; Fernández, J.J. Studies on the biosynthesis of the polyether marine toxin dinophysistoxin-1 (DTX-1). Tetrahedron Lett. 1994, 35, 1441–1444. [Google Scholar] [CrossRef]
- Vilches, T.S.; Norte, M.; Daranas, A.H.; Fernández, J.J. Biosynthetic Studies on Water-Soluble Derivative 5c (DTX5c). Mar. Drugs 2012, 10, 2234–2245. [Google Scholar] [CrossRef] [PubMed]
- Hwang, D.F.; Lu, Y.H. Influence of environmental and nutritional factors on growth, toxicity, and toxin profile of dinoflagellate Alexandrium minutum. Toxicon 2000, 38, 1491–1503. [Google Scholar] [CrossRef] [PubMed]
- Chan, L.L.; Hodgkiss, I.J.; Lam, P.K.S.; Wan, J.M.F.; Chou, H.N.; Lum, J.H.K.; Lo, M.G.Y.; Mak, A.S.C.; Sit, W.H.; Lo, S.C.L. Use of two-dimensional gel electrophoresis to differentiate morphospecies of Alexandrium minutum, a paralytic shellfish poisoning toxin-producing dinoflagellate of harmful algal blooms. Proteomics 2005, 5, 1580–1593. [Google Scholar] [CrossRef] [PubMed]
- Chan, L.L.; Sit, W.-H.; Lam, P.K.-S.; Hsieh, D.P.H.; Hodgkiss, I.J.; Wan, J.M.-F.; Ho, A.Y.-T.; Choi, N.M.-C.; Wang, D.-Z.; Dudgeon, D. Identification and characterization of a “biomarker of toxicity” from the proteome of the paralytic shellfish toxin-producing dinoflagellateAlexandrium tamarense (Dinophyceae). Proteomics 2006, 6, 654–666. [Google Scholar] [CrossRef] [PubMed]
- Lee, F.W.-F.; Morse, D.; Lo, S.C.-L. Identification of Two Plastid Proteins in the Dinoflagellate Alexandrium affine That Are Substantially down-Regulated by Nitrogen-Depletion. J. Proteome Res. 2009, 8, 5080–5092. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Wang, D.; Dong, H.; Xie, Z.; Hong, H. Proteomics of a toxic dinoflagellate Alexandrium catenella DH01: Detection and identification of cell surface proteins using fluorescent labeling. Chin. Sci. Bull. 2012, 57, 3320–3327. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zhang, Y.; Xie, Z.-X.; He, Z.-P.; Lin, L.; Wang, D.-Z. Quantitative proteomic analysis reveals evolutionary divergence and species-specific peptides in the Alexandrium tamarense complex (Dinophyceae). J. Proteom. 2013, 86, 85–96. [Google Scholar] [CrossRef]
- Wang, D.Z.; Li, C.; Zhang, Y.; Wang, Y.Y.; He, Z.P.; Lin, L.; Hong, H.S. Quantitative proteomic analysis of differentially expressed proteins in the toxicity-lost mutant of Alexandrium catenella (Dinophyceae) in the exponential phase. J. Proteom. 2012, 75, 5564–5577. [Google Scholar] [CrossRef]
- Wang, D.; Lin, L.; Wang, M.; Li, C.; Hong, H. Proteomic analysis of a toxic dinoflagellate Alexandrium catenella under different growth phases and conditions. Chin. Sci. Bull. 2012, 57, 3328–3341. [Google Scholar] [CrossRef]
- Wang, D.-Z.; Gao, Y.; Lin, L.; Hong, H.-S. Comparative Proteomic Analysis Reveals Proteins Putatively Involved in Toxin Biosynthesis in the Marine Dinoflagellate Alexandrium catenella. Mar. Drugs 2013, 11, 213–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mihali, T.K.; Kellmann, R.; Neilan, B.A. Characterisation of the paralytic shellfish toxin biosynthesis gene clusters in Anabaena circinalis AWQC131C and Aphanizomenon sp. NH-5. BMC Biochem. 2009, 10, 8. [Google Scholar] [CrossRef] [Green Version]
- Lee, T.; Chan, P.; Xu, S.; Lee, F. Comparison of growth and toxicity responses between non-toxic and toxic strains of Prorocentrum hoffmannianum. Aquat. Biol. 2020, 29, 59–70. [Google Scholar] [CrossRef]
- Chan, L.L.; Hodgkiss, I.J.; Lu, S.; Lo, S.C.-L. Use of two-dimensional gel electrophoresis proteome reference maps of dinoflagellates for species recognition of causative agents of harmful algal blooms. Proteomics 2004, 4, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Lee, F.; Lo, S. Proteomic study of micro-algae: Sample preparation for two-dimensional gel electrophoresis and de novo peptide sequencing using MALDI-TOF MS. Curr. Proteom. 2007, 4, 67–78. [Google Scholar] [CrossRef]
- Chudnovsky, Y.; Li, J.F.; Rizzo, P.J.; Hastings, J.W.; Fagan, T.F. Cloning, expression, and characterization of a histone-like protein from the marine dinoflagellate Lingulodinium polyedrum (Dinophyceae). J. Phycol. 2002, 38, 543–550. [Google Scholar] [CrossRef]
- Liu, B.; Lo, S.C.-L.; Matton, D.P.; Lang, B.F.; Morse, D. Daily Changes in the Phosphoproteome of the Dinoflagellate Lingulodinium. Protist 2012, 163, 746–754. [Google Scholar] [CrossRef]
- Roy, S.; Morse, D. A Full Suite of Histone and Histone Modifying Genes Are Transcribed in the Dinoflagellate Lingulodinium. PLoS ONE 2012, 7, e34340. [Google Scholar] [CrossRef] [PubMed]
- Larkum, T.; Howe, C. Molecular Aspects of Light-harvesting Processes in Algae. In Advances in Botanical Research; Academic Press: Cambridge, MA, USA, 1997; Volume 27, pp. 257–330. [Google Scholar] [CrossRef]
- Norris, B.J.; Miller, D.J. Nucleotide sequence of a cDNA clone encoding the precursor of the peridinin-chlorophyll a-binding protein from the dinoflagellate Symbiodinium sp. Plant Mol. Biol. 1994, 24, 673–677. [Google Scholar] [CrossRef]
- Schulte, T.; Johanning, S.; Hofmann, E. Structure and function of native and refolded peridinin-chlorophyll-proteins from dinoflagellates. Eur. J. Cell Biol. 2010, 89, 990–997. [Google Scholar] [CrossRef]
- Iglesias-Prieto, R.; Govind, N.S.; Trench, R.K. Apoprotein composition and spectroscopic characterization of the water-soluble peridinin—Chlorophyll a —Proteins from three symbiotic dinoflagellates. Proc. R. Soc. B Boil. Sci. 1991, 246, 275–283. [Google Scholar] [CrossRef]
- Weis, V.; Verde, E.A.; Reynolds, W.S. Characterization of a short form peridinin-chlorophyll-protein (PCP) cDNA and protein from the symbiotic dinoflagellate Symbiodinium muscatinei (Dinophyceae) from the sea anemone Anthopleura elegantissima (Cnidaria). J. Phycol. 2002, 38, 157–163. [Google Scholar] [CrossRef]
- Sharples, F.P.; Wrench, P.M.; Ou, K.; Hiller, R.G. Two distinct forms of the peridinin-chlorophyll a-protein from Amphidinium carterae. Biochim. Biophys. Acta (BBA)-Bioenerg. 1996, 1276, 117–123. [Google Scholar] [CrossRef] [Green Version]
- Ilagan, R.P.; Shima, S.; Melkozernov, A.; Lin, S.; Blankenship, R.E.; Sharples, F.P.; Hiller, R.G.; Birge, R.R.; Frank, H.A. Spectroscopic Properties of the Main-Form and High-Salt Peridinin−Chlorophyll a Proteins from Amphidinium carterae. Biochemistry 2004, 43, 1478–1487. [Google Scholar] [CrossRef] [PubMed]
- Haxo, F.T.; Kycia, J.H.; Somers, G.F.; Bennett, A.; Siegelman, H.W. Peridinin-Chlorophyll a Proteins of the Dinoflagellate Amphidinium carterae (Plymouth 450). Plant Physiol. 1976, 57, 297–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jean, N.; Dumont, E.; Herzi, F.; Balliau, T.; Laabir, M.; Masseret, E.; Mounier, S. Modifications of the soluble proteome of a mediterranean strain of the invasive neurotoxic dinoflagellate Alexandrium catenella under metal stress conditions. Aquat. Toxicol. 2017, 188, 80–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prézelin, B.B. The role of peridinin-chlorophyll a-proteins in the photosynthetic light adaption of the marine dinoflagellate, Glenodinium sp. Planta 1976, 130, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Cembella, A.D.; Quilliam, M.A. Cell cycle and toxin production in the benthic dinoflagellate Prorocentrum lima. Mar. Biol. 1999, 134, 541–549. [Google Scholar] [CrossRef]
- Lee, T.C.-H.; Fong, F.L.-Y.; Ho, K.-C.; Lee, F.W.-F. The Mechanism of Diarrhetic Shellfish Poisoning Toxin Production in Prorocentrum spp.: Physiological and Molecular Perspectives. Toxins 2016, 8, 272. [Google Scholar] [CrossRef]
- Rein, K.S.; Snyder, R.V. The Biosynthesis of Polyketide Metabolites by Dinoflagellates. Adv. Appl. Microbiol. 2006, 59, 93–125. [Google Scholar] [CrossRef] [Green Version]
- Kaebernick, M.; Neilan, B.A.; Boörner, T.; Dittmann, E. Light and the Transcriptional Response of the Microcystin Biosynthesis Gene Cluster. Appl. Environ. Microbiol. 2000, 66, 3387–3392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, J.; Zhang, H.; Kang, Y.; Bina, D.; Lo, C.S.; Blankenship, R.E. Characterization of the peridinin–chlorophyll a-protein complex in the dinoflagellate Symbiodinium. Biochim. Biophys. Acta (BBA)-Bioenerg. 2012, 1817, 983–989. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbier, M.; Amzil, Z.; Mondeguer, F.; Bhaud, Y.; Soyer-Gobillard, M.-O.; Lassus, P. Okadaic acid and PP2A cellular immunolocalization in Prorocentrum lima (Dinophyceae). Phycologia 1999, 38, 41–46. [Google Scholar] [CrossRef]
- Zhou, J.; Fritz, L. Okadaic acid antibody localizes to chloroplasts in the DSP-toxin-producing dinoflagellates Prorocentrum lima and Prorocentrum maculosum. Phycologia 1994, 33, 455–461. [Google Scholar] [CrossRef]
- Needham, J.; Hu, T.; McLachlan, J.L.; Walter, J.A.; Wright, J.L.C. Biosynthetic studies of the DSP toxin DTX-4 and an okadaic acid diol ester. J. Chem. Soc. Chem. Commun. 1995, 16, 1623–1624. [Google Scholar] [CrossRef]
- Wright, J.L.; Hu, T.; McLachlan, J.L.; Needham, J.; Walter, J.A. Biosynthesis of DTX-4: Confirmation of a polyketide pathway, proof of a Baeyer−Villiger oxidation step, and evidence for an unusual carbon deletion process. J. Am. Chem. Soc. 1996, 118, 8757–8758. [Google Scholar] [CrossRef]
- Wood, H.G. Life with CO or CO2 and H2 as a source of carbon and energy. FASEB J. 1991, 5, 156–163. [Google Scholar] [CrossRef]
- Burris, J.E. Photosynthesis, photorespiration, and dark respiration in eight species of algae. Mar. Biol. 1977, 39, 371–379. [Google Scholar] [CrossRef]
- Lloyd, N.D.H.; Canvin, D.T.; Culver, D.A. Photosynthesis and Photorespiration in Algae. Plant Physiol. 1977, 59, 936–940. [Google Scholar] [CrossRef]
- Guillard, R.R.L.; E Hargraves, P. Stichochrysis immobilis is a diatom, not a chrysophyte. Phycologia 1993, 32, 234–236. [Google Scholar] [CrossRef]
- Lee, T.C.-H.; Lai, K.K.-Y.; Kwok, C.S.-N.; Xu, S.J.-L.; Lee, F.W.-F. Comparison of Five TRIzol-Based Protein Preparation Methods for 2-DE Production from Challenging Marine Dinoflagellate Samples: A Case Study on Two Benthic Prorocentrum Species. J. Mar. Sci. Eng. 2020, 8, 363. [Google Scholar] [CrossRef]
- Lee, F.W.-F.; Lo, S.C.-L. The use of Trizol reagent (phenol/guanidine isothiocyanate) for producing high quality two-dimensional gel electrophoretograms (2-DE) of dinoflagellates. J. Microbiol. Methods 2008, 73, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Chan, L.L.; Lo, S.C.-L.; Hodgkiss, I.J. Proteomic study of a model causative agent of harmful red tide, Prorocentrum triestinum I: Optimization of sample preparation methodologies for analyzing with two-dimensional electrophoresis. Proteomics 2002, 2, 1169–1186. [Google Scholar] [CrossRef]
- Chan, L.L.; Hodgkiss, I.J.; Wan, J.M.-F.; Lum, J.H.-K.; Mak, A.S.-C.; Sit, W.-H.; Lo, S.C.L. Proteomic study of a model causative agent of harmful algal blooms, Prorocentrum triestinum II: The use of differentially expressed protein profiles under different growth phases and growth conditions for bloom prediction. Proteomics 2004, 4, 3214–3226. [Google Scholar] [CrossRef] [PubMed]
- Perkins, D.N.; Pappin, D.J.; Creasy, D.M.; Cottrell, J.S. Probability-based protein identification by searching sequence databases using mass spectrometry data. Electrophoresis 1999, 20, 3551–3567. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spots | Fragment Peaks (m/z) | Protein Identity | Differential Expression |
|---|---|---|---|
| T2b | 1229.6 | Putative uncharacterized protein | Only expressed in CCMP2804 compared to CCMP683 under normal condition |
| T5b,c T6a–d T7a–d T10a–d T11a–d T12a–c TP5 TP6a,b TP7a–c TP8a–d | 1258.6 2393.2 | Peridinin-chlorophyll a-binding protein | Two- to four-fold upregulation or only expressed in CCMP2804 compared to CCMP683 under normal condition or phosphate depletion |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, T.C.-H.; Lai, K.K.-Y.; Xu, S.J.-L.; Lee, F.W.-F. Upregulation of Peridinin-Chlorophyll A-Binding Protein in a Toxic Strain of Prorocentrum hoffmannianum under Normal and Phosphate-Depleted Conditions. Int. J. Mol. Sci. 2023, 24, 1735. https://doi.org/10.3390/ijms24021735
Lee TC-H, Lai KK-Y, Xu SJ-L, Lee FW-F. Upregulation of Peridinin-Chlorophyll A-Binding Protein in a Toxic Strain of Prorocentrum hoffmannianum under Normal and Phosphate-Depleted Conditions. International Journal of Molecular Sciences. 2023; 24(2):1735. https://doi.org/10.3390/ijms24021735
Chicago/Turabian StyleLee, Thomas Chun-Hung, Kaze King-Yip Lai, Steven Jing-Liang Xu, and Fred Wang-Fat Lee. 2023. "Upregulation of Peridinin-Chlorophyll A-Binding Protein in a Toxic Strain of Prorocentrum hoffmannianum under Normal and Phosphate-Depleted Conditions" International Journal of Molecular Sciences 24, no. 2: 1735. https://doi.org/10.3390/ijms24021735