
Rab44 Deficiency Induces Impaired Immune Responses to Nickel Allergy

 ,
,  and
and 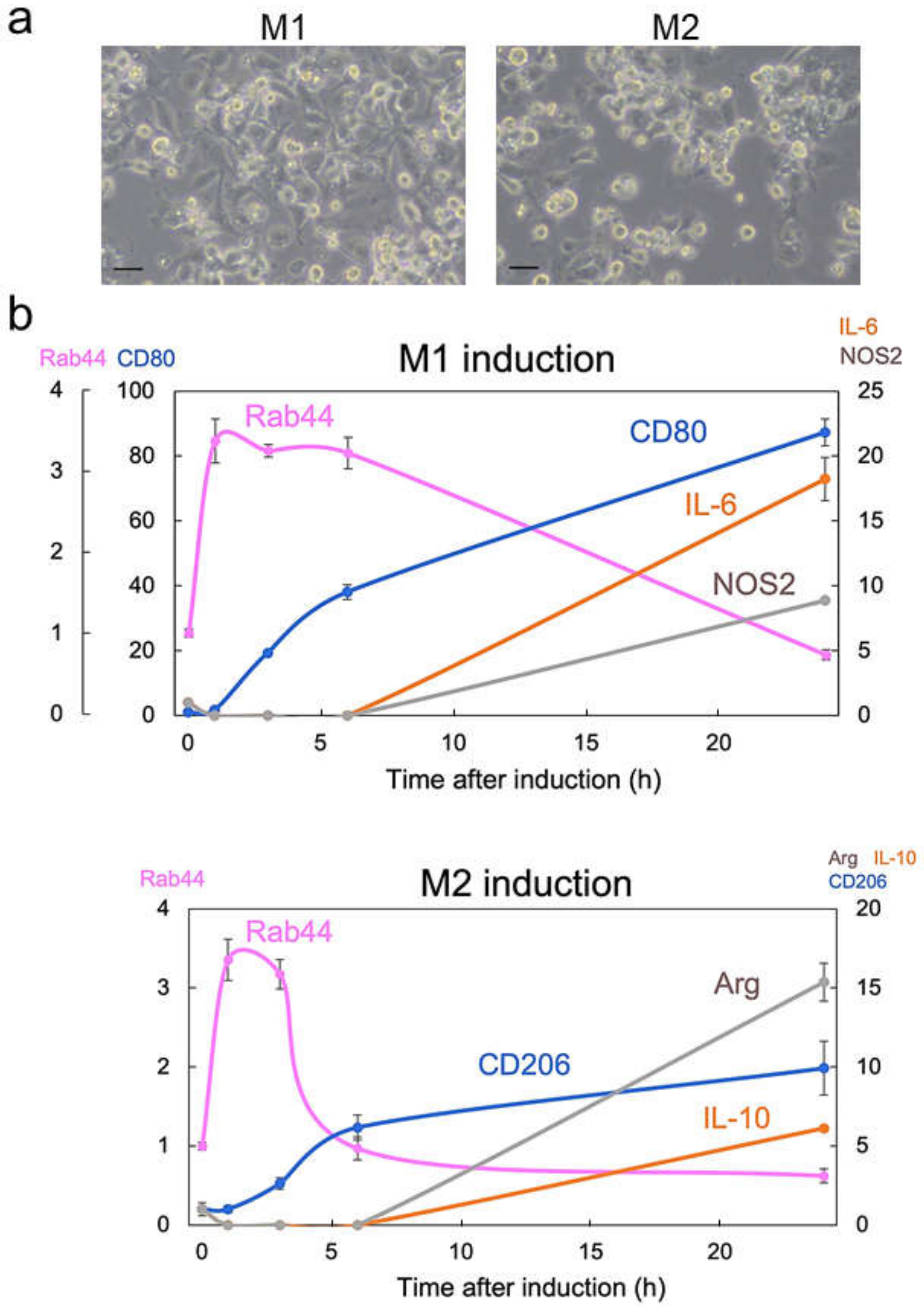{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Rab44 Is Upregulated at an Early Phase during Differentiation of M1- and M2-Type Macrophages
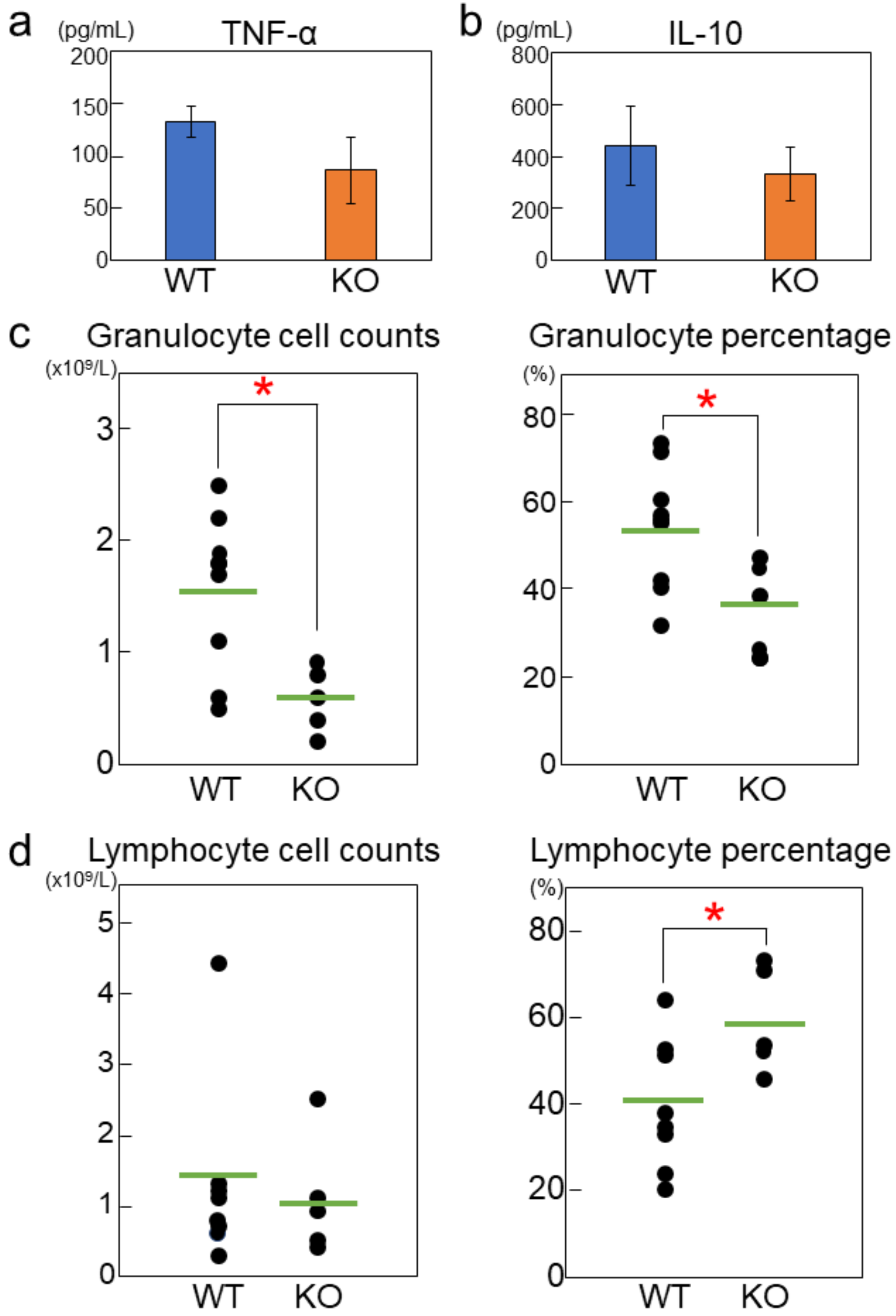
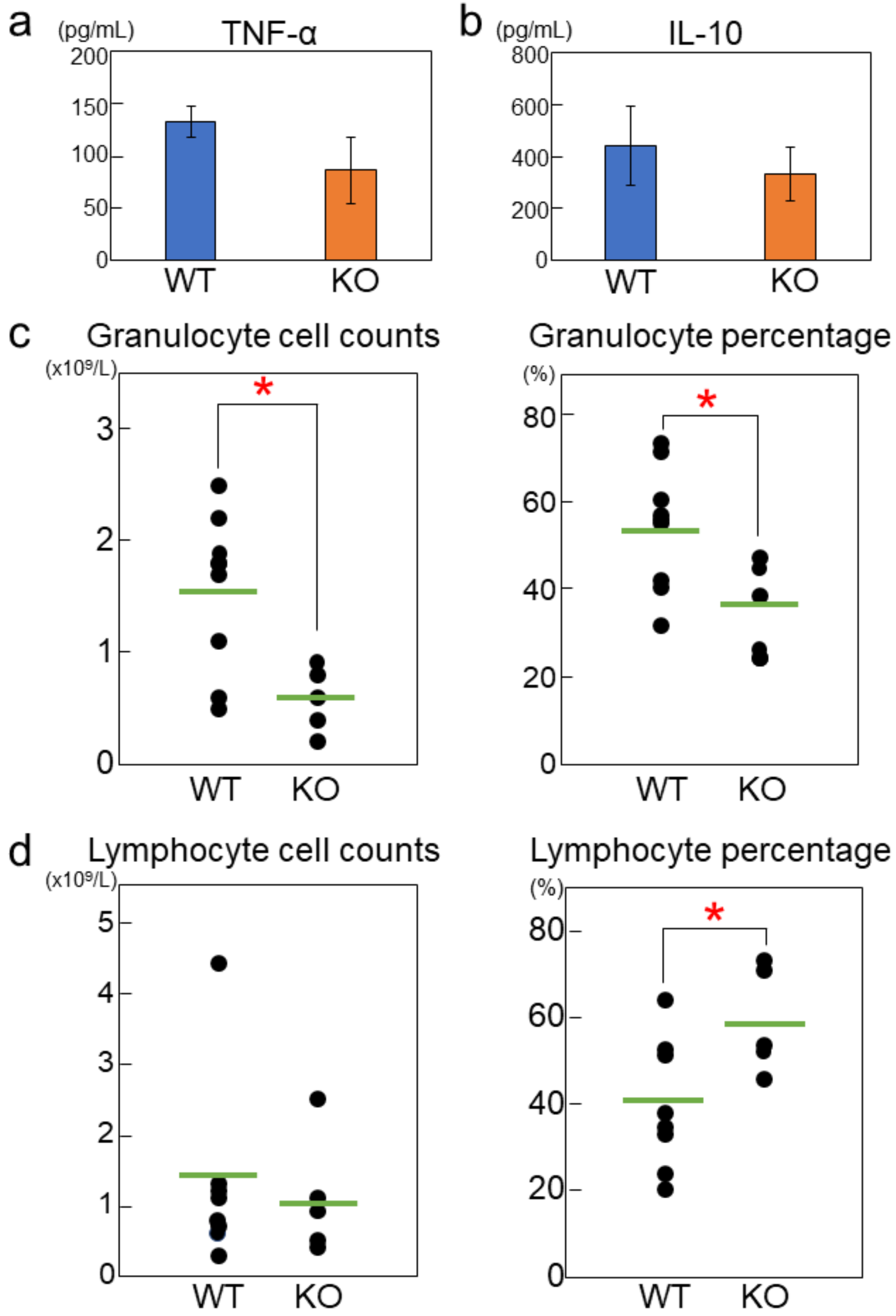
2.2. Rab44-Deficient Mice Exhibit Impaired Cytokine Production and Differential Immune Responses
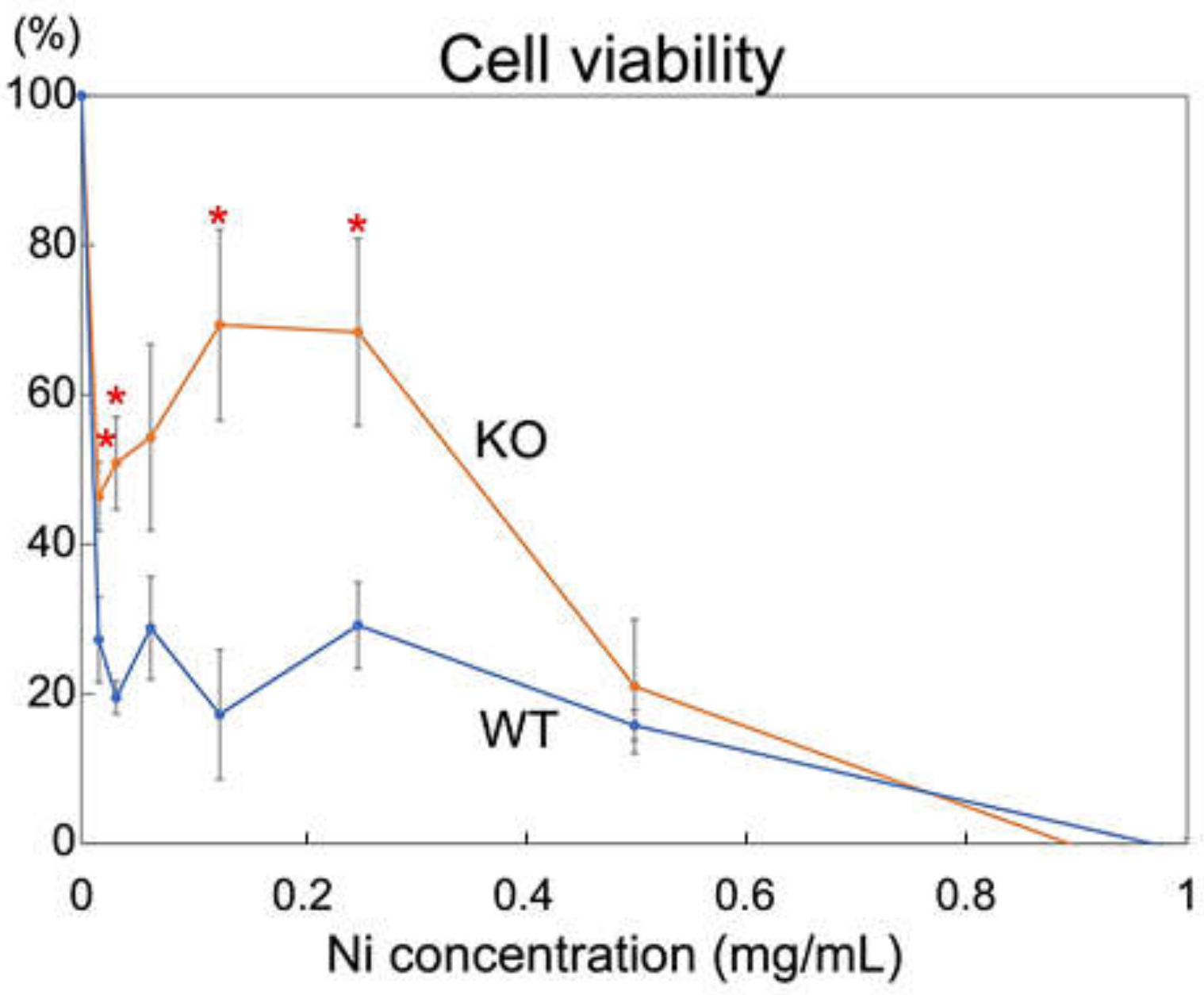
2.3. Rab44-Deficient Macrophages Show an Impaired Viability to Nickel Solution
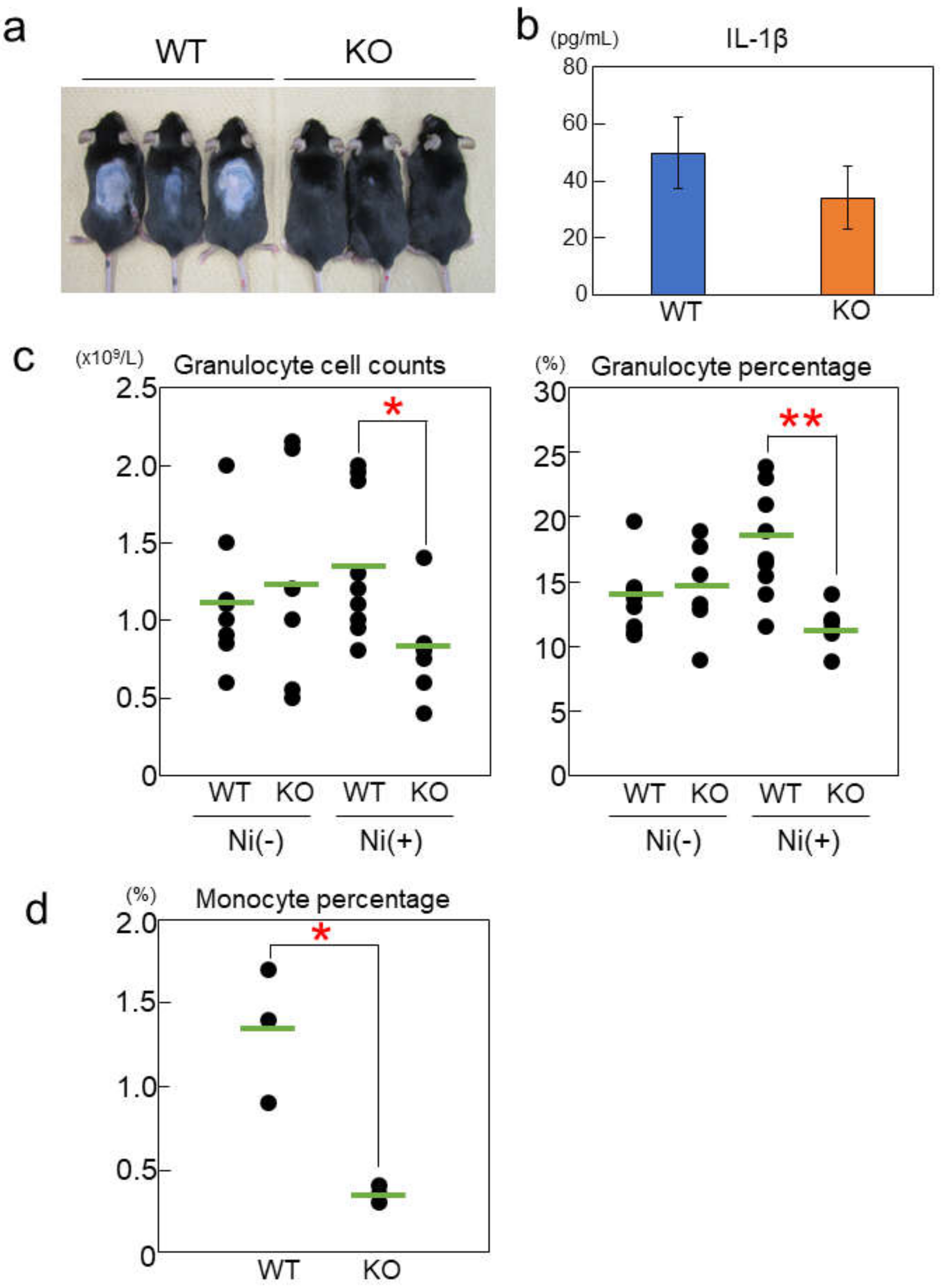
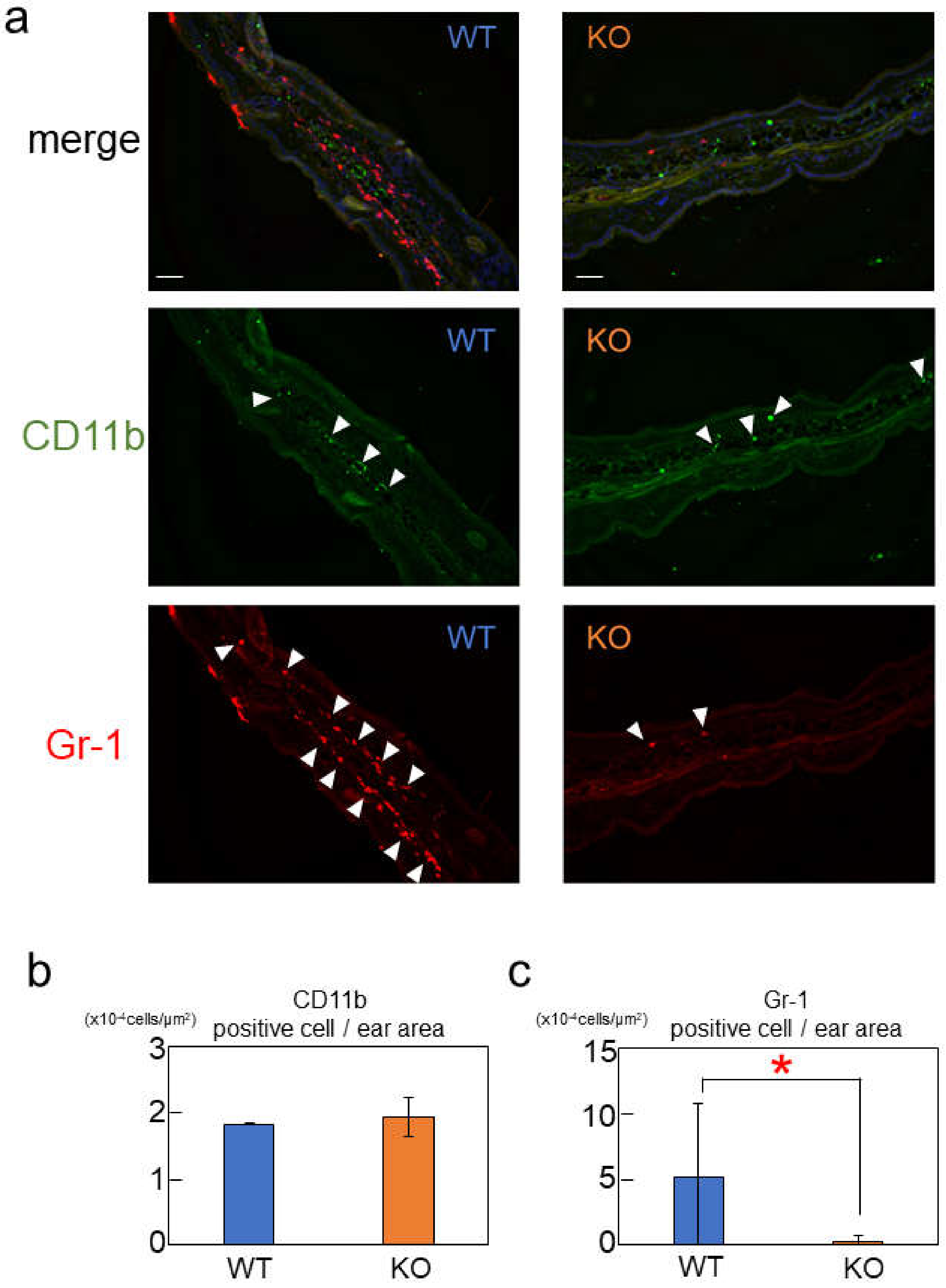
2.4. Rab44-Deficient Mice Show Different Frequencies of Immune Cells upon Nickel-Hypersensitivity Induction
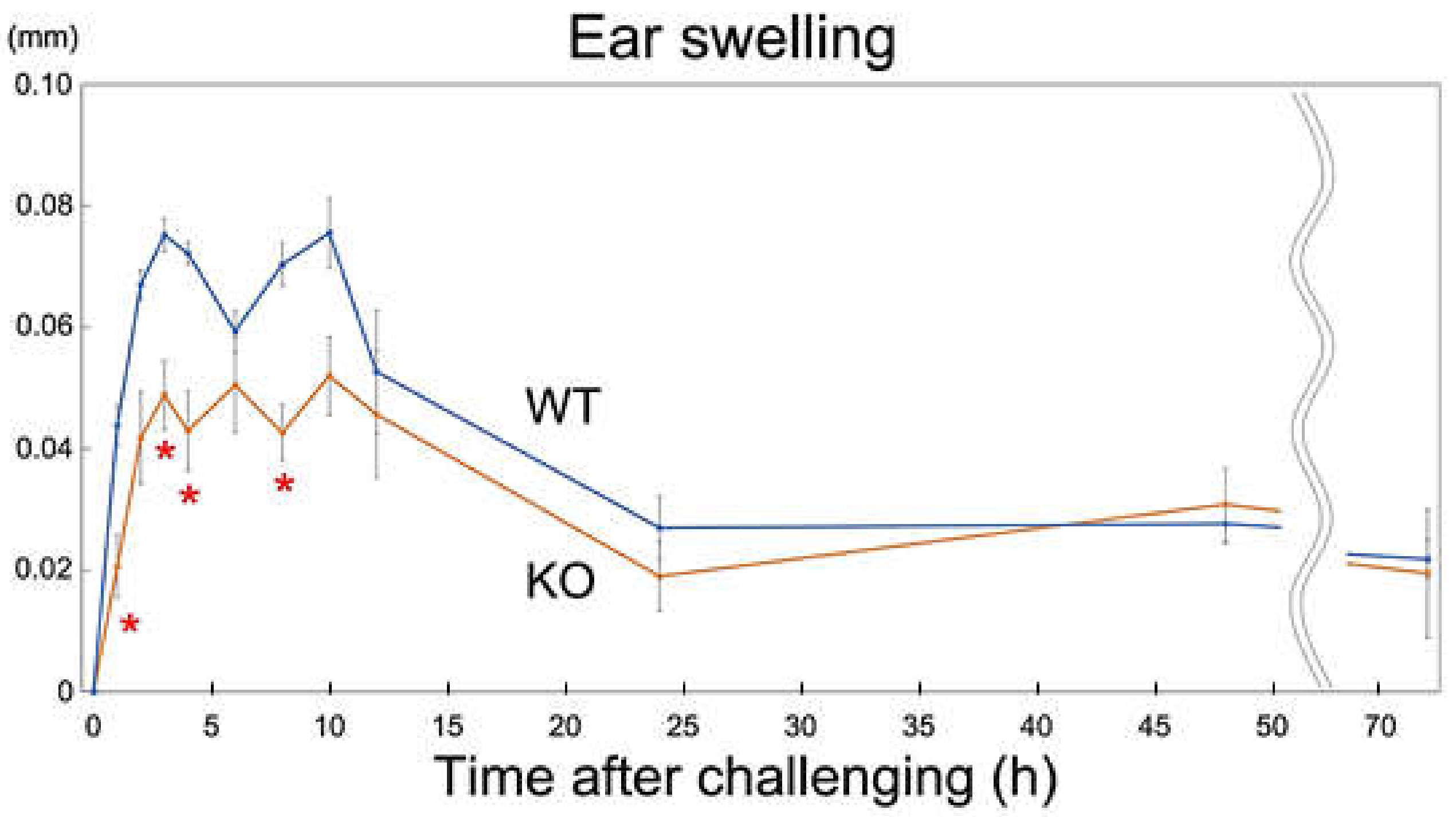
2.5. Rab44-Deficient Mice Show Reduced Nickel-Induced Inflammation
3. Discussion
4. Materials and Methods
4.1. Antibodies and Reagents
4.2. Animals
4.3. Cell Culture
4.4. Reverse Transcription-Polymerase Chain Reaction (RT-PCR) Analysis
4.5. Cell Viability to Nickel Solution
4.6. Nickel-Induced Allergic Contact Dermatitis
4.7. Immunofluorescence Microscopy
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Stenmark, H. Rab GTPases as coordinators of vesicle traffic. Nat. Rev. Mol. Cell Biol. 2009, 10, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Zhen, Y.; Stenmark, H. Cellular functions of Rab GTPases at a glance. J. Cell Sci. 2015, 128, 3171–3176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mizuno-Yamasaki, E.; Rivera-Molina, F.; Novick, P. GTPase networks in membrane traffic. Annu. Rev. Biochem. 2012, 81, 637–659. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Homma, Y.; Hiragi, S.; Fukuda, M. Rab family of small GTPases: An updated view on their regulation and functions. FEBS J. 2021, 288, 36–55. [Google Scholar] [CrossRef] [PubMed]
- Hutagalung, A.H.; Novick, P.J. Role of Rab GTPases in membrane traffic and cell physiology. Physiol. Rev. 2011, 91, 119–149. [Google Scholar] [CrossRef] [Green Version]
- Mitra, S.; Cheng, K.W.; Mills, G.B. Rab GTPases implicated in inherited and acquired disorders. Semin. Cell Dev. Biol. 2011, 22, 57–68. [Google Scholar] [CrossRef] [Green Version]
- Prashar, A.; Schnettger, L.; Bernard, E.M.; Gutierrez, M.G. Rab GTPases in Immunity and Inflammation. Front. Cell. Infect. Microbiol. 2017, 7, 435. [Google Scholar] [CrossRef] [Green Version]
- Banworth, M.J.; Li, G. Consequences of Rab GTPase dysfunction in genetic or acquired human diseases. Small GTPases 2018, 9, 158–181. [Google Scholar] [CrossRef] [Green Version]
- Ohishi, Y.; Ammann, S.; Ziaee, V.; Strege, K.; Groß, M.; Amos, C.V.; Shahrooei, M.; Ashournia, P.; Razaghian, A.; Griffiths, G.M.; et al. Griscelli Syndrome Type 2 Sine Albinism: Unraveling Differential RAB27A Effector Engagement. Front. Immunol. 2020, 11, 612977. [Google Scholar] [CrossRef]
- Fukuda, M. Rab GTPases: Key players in melanosome biogenesis, transport, and transfer. Pigment Cell Melanoma Res. 2021, 34, 222–235. [Google Scholar] [CrossRef]
- Ohira, M.; Oshitani, N.; Hosomi, S.; Watanabe, K.; Yamagami, H.; Tominaga, K.; Watanabe, T.; Fujiwara, Y.; Maeda, K.; Hirakawa, K.; et al. Dislocation of Rab13 and vasodilator-stimulated phosphoprotein in inactive colon epithelium in patients with Crohn’s disease. Int. J. Mol. Med. 2009, 24, 829–835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gerondopoulos, A.; Langemeyer, L.; Liang, J.R.; Linford, A.; Barr, F.A. BLOC-3 mutated in Hermansky-Pudlak syndrome is a Rab32/38 guanine nucleotide exchange factor. Curr. Biol. 2012, 22, 2135–2139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bowman, S.L.; Bi-Karchin, J.; Le, L.; Marks, M.S. The road to lysosome-related organelles: Insights from Hermansky-Pudlak syndrome and other rare diseases. Traffic 2019, 20, 404–435. [Google Scholar] [CrossRef] [Green Version]
- Tsukuba, T.; Yamaguchi, Y.; Kadowaki, T. Large Rab GTPases: Novel Membrane Trafficking Regulators with a Calcium Sensor and Functional Domains. Int. J. Mol. Sci. 2021, 22, 7691. [Google Scholar] [CrossRef] [PubMed]
- Ogawa, K.; Kadowaki, T.; Tokuhisa, M.; Yamaguchi, Y.; Umeda, M.; Tsukuba, T. Role of the EF-hand and coiled-coil domains of human Rab44 in localisation and organelle formation. Sci. Rep. 2020, 10, 19149. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Sakai, E.; Okamoto, K.; Kajiya, H.; Okabe, K.; Naito, M.; Kadowaki, T.; Tsukuba, T. Rab44, a novel large Rab GTPase, negatively regulates osteoclast differentiation by modulating intracellular calcium levels followed by NFATc1 activation. Cell. Mol. Life Sci. 2018, 75, 33–48. [Google Scholar] [CrossRef]
- Tokuhisa, M.; Kadowaki, T.; Ogawa, K.; Yamaguchi, Y.; Kido, M.A.; Gao, W.; Umeda, M.; Tsukuba, T. Expression and localisation of Rab44 in immune-related cells change during cell differentiation and stimulation. Sci. Rep. 2020, 10, 10728. [Google Scholar] [CrossRef]
- Kadowaki, T.; Yamaguchi, Y.; Ogawa, K.; Tokuhisa, M.; Okamoto, K.; Tsukuba, T. Rab44 isoforms similarly promote lysosomal exocytosis, but exhibit differential localization in mast cells. FEBS Open Bio 2021, 11, 1165–1185. [Google Scholar] [CrossRef]
- Kadowaki, T.; Yamaguchi, Y.; Kido, M.A.; Abe, T.; Ogawa, K.; Tokuhisa, M.; Gao, W.; Okamoto, K.; Kiyonari, H.; Tsukuba, T. The large GTPase Rab44 regulates granule exocytosis in mast cells and IgE-mediated anaphylaxis. Cell. Mol. Immunol. 2020, 17, 1287–1289. [Google Scholar] [CrossRef] [Green Version]
- Kinbara, M.; Bando, K.; Shiraishi, D.; Kuroishi, T.; Nagai, Y.; Ohtsu, H.; Takano-Yamamoto, T.; Sugawara, S.; Endo, Y. Mast cell histamine-mediated transient inflammation following exposure to nickel promotes nickel allergy in mice. Exp. Dermatol. 2016, 25, 466–471. [Google Scholar] [CrossRef]
- Banu, Y.; Watanabe, T. Augmentation of antigen receptor-mediated responses by histamine H1 receptor signaling. J. Exp. Med. 1999, 189, 673–682. [Google Scholar] [CrossRef] [Green Version]
- Jutel, M.; Watanabe, T.; Akdis, M.; Blaser, K.; Akdis, C.A. Immune regulation by histamine. Curr. Opin. Immunol. 2002, 14, 735–740. [Google Scholar] [CrossRef]
- Martinez, F.O.; Gordon, S. The M1 and M2 paradigm of macrophage activation: Time for reassessment. F1000Prime Rep. 2014, 6, 13. [Google Scholar] [CrossRef] [Green Version]
- Yunna, C.; Mengru, H.; Lei, W.; Weidong, C. Macrophage M1/M2 polarization. Eur. J. Pharmacol. 2020, 877, 173090. [Google Scholar] [CrossRef]
- Mohd Yasin, Z.N.; Mohd Idrus, F.N.; Hoe, C.H.; Yvonne-Tee, G.B. Macrophage polarization in THP-1 cell line and primary monocytes: A systematic review. Differ. Res. Biol. Divers. 2022, 128, 67–82. [Google Scholar] [CrossRef]
- Takemoto, R.; Kamiya, T.; Atobe, T.; Hara, H.; Adachi, T. Regulation of lysyl oxidase expression in THP-1 cell-derived M2-like macrophages. J. Cell. Biochem. 2021, 122, 777–786. [Google Scholar] [CrossRef]
- De Graaf, N.P.J.; Roffel, S.; Gibbs, S.; Kleverlaan, C.J.; Lopez Gonzalez, M.; Rustemeyer, T.; Feilzer, A.J.; Bontkes, H.J. Nickel allergy is associated with a broad spectrum cytokine response. Contact Dermat. 2023, 88, 10–17. [Google Scholar] [CrossRef]
- Sato, N.; Kinbara, M.; Kuroishi, T.; Kimura, K.; Iwakura, Y.; Ohtsu, H.; Sugawara, S.; Endo, Y. Lipopolysaccharide promotes and augments metal allergies in mice, dependent on innate immunity and histidine decarboxylase. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2007, 37, 743–751. [Google Scholar] [CrossRef]
- Bando, K.; Kuroishi, T.; Sugawara, S.; Endo, Y. Interleukin-1 and histamine are essential for inducing nickel allergy in mice. Clin. Exp. Allergy J. Br. Soc. Allergy Clin. Immunol. 2019, 49, 1362–1373. [Google Scholar] [CrossRef]
- Krayem, I.; Sohrabi, Y.; Javorková, E.; Volkova, V.; Strnad, H.; Havelková, H.; Vojtíšková, J.; Aidarova, A.; Holáň, V.; Demant, P.; et al. Genetic Influence on Frequencies of Myeloid-Derived Cell Subpopulations in Mouse. Front. Immunol. 2021, 12, 760881. [Google Scholar] [CrossRef]
- Sakai, E.; Shimada-Sugawara, M.; Nishishita, K.; Fukuma, Y.; Naito, M.; Okamoto, K.; Nakayama, K.; Tsukuba, T. Suppression of RANKL-dependent heme oxygenase-1 is required for high mobility group box 1 release and osteoclastogenesis. J. Cell. Biochem. 2012, 113, 486–498. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, Y.; Kadowaki, T.; Aibara, N.; Ohyama, K.; Okamoto, K.; Sakai, E.; Tsukuba, T. Coronin1C Is a GDP-Specific Rab44 Effector That Controls Osteoclast Formation by Regulating Cell Motility in Macrophages. Int. J. Mol. Sci. 2022, 23, 6619. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Noguromi, M.; Yamaguchi, Y.; Sato, K.; Oyakawa, S.; Okamoto, K.; Murata, H.; Tsukuba, T.; Kadowaki, T. Rab44 Deficiency Induces Impaired Immune Responses to Nickel Allergy. Int. J. Mol. Sci. 2023, 24, 994. https://doi.org/10.3390/ijms24020994
Noguromi M, Yamaguchi Y, Sato K, Oyakawa S, Okamoto K, Murata H, Tsukuba T, Kadowaki T. Rab44 Deficiency Induces Impaired Immune Responses to Nickel Allergy. International Journal of Molecular Sciences. 2023; 24(2):994. https://doi.org/10.3390/ijms24020994
Chicago/Turabian StyleNoguromi, Mayuko, Yu Yamaguchi, Keiko Sato, Shun Oyakawa, Kuniaki Okamoto, Hiroshi Murata, Takayuki Tsukuba, and Tomoko Kadowaki. 2023. "Rab44 Deficiency Induces Impaired Immune Responses to Nickel Allergy" International Journal of Molecular Sciences 24, no. 2: 994. https://doi.org/10.3390/ijms24020994
APA StyleNoguromi, M., Yamaguchi, Y., Sato, K., Oyakawa, S., Okamoto, K., Murata, H., Tsukuba, T., & Kadowaki, T. (2023). Rab44 Deficiency Induces Impaired Immune Responses to Nickel Allergy. International Journal of Molecular Sciences, 24(2), 994. https://doi.org/10.3390/ijms24020994