
Events Leading to the Establishment of Pregnancy and Placental Formation: The Need to Fine-Tune the Nomenclature on Pregnancy and Gestation

Abstract
:1. Introduction
2. Methodology
3. Evidence of Early Embryonic Loss
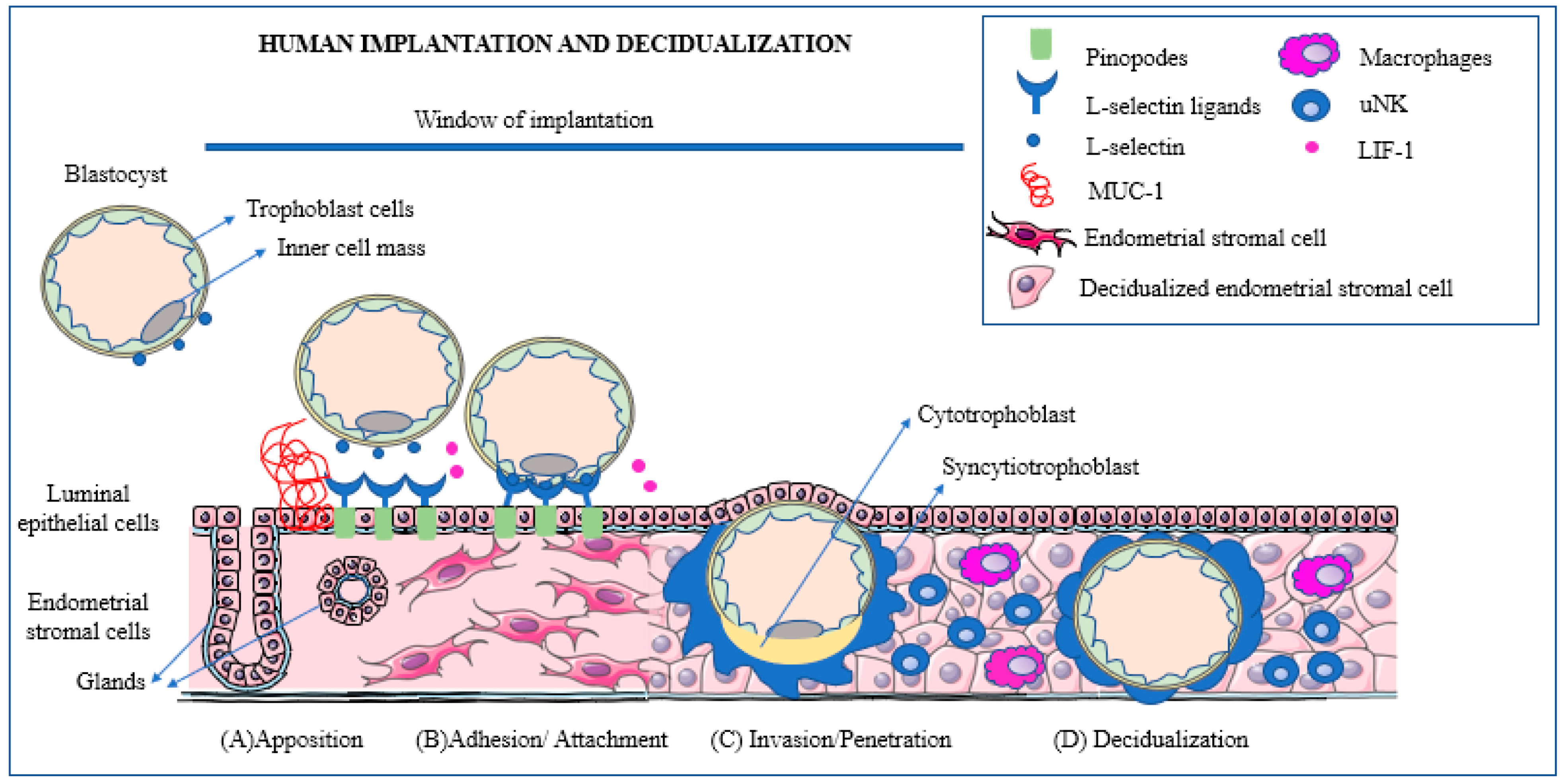
4. Mechanisms Involved in Early Embryo Selection
- The blastocyst moves towards the uterine cavity and contemporarily the decidualized endometrium evolves to a receptive phenotype, resulting in a biochemical cross-talk with the embryo.
- The pre-implantation embryo begins to secrete factors capable of modulating the implantation site while, in turn, the decidua secretes cytokines and growth factors modulating embryonic differentiation and development.
- In the presence of a proper biochemical environment, the embryo and the decidua jointly promote trophoblast invasion. This process can be altered in a number of ways resulting in early embryonic loss.
4.1. Factors Secreted by the Pre-Implantation Embryo
4.1.1. The Early Pregnancy Factor
4.1.2. The Pre-Implantation Factor
4.1.3. The Embryo-Derived Platelet-Activating Factor
4.1.4. The Human Chorionic Gonadotrophin
4.2. Factors Produced by the Endometrium
4.3. The Role of the Immune System in Early Pregnancy Wastage
5. Factors Predictive of Embryonic Loss
The Embryo–Maternal Dialogue
6. The Need for New Terminology
7. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Benagiano, G.; Mancuso, S.; Gianaroli, L.; Di Renzo, G.C. Gestation vs. Pregnancy. Am. J. Obstet. Gynecol. 2023, 229, 91–92. [Google Scholar] [CrossRef] [PubMed]
- Green, R.M. Embryo as epiphenomenon: Some cultural, social and economic forces driving the stem cell debate. J. Med. Ethics 2008, 34, 840–844. [Google Scholar] [CrossRef] [PubMed]
- Macklon, N.S.; Geraedts, J.P.; Fauser, B.C. Conception to ongoing pregnancy: The ‘black box’ of early pregnancy loss. Hum. Reprod. Update 2002, 8, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Benagiano, G.; Farris, M.; Grudzinskas, G. Fate of fertilized human oocytes. Reprod. Biomed. Online 2010, 21, 732–741. [Google Scholar] [CrossRef] [PubMed]
- The Annual Capri Workshop Group. Early pregnancy loss: The default outcome for fertilized human oocytes. J. Assist. Reprod. Genet. 2020, 37, 1057–1063. [Google Scholar] [CrossRef]
- Steiner, A.Z.; Pritchard, D.A.; Young, S.L.; Herring, A.H. Peri-implantation intercourse lowers fecundability. Fertil. Steril. 2014, 102, 178–182. [Google Scholar] [CrossRef]
- World Health Organisation. Congenital Anomalies. Available online: https://www.who.int/health-topics/congenital-anomalies#tab=tab_1 (accessed on 22 July 2022).
- Nybo Andersen, A.M.; Wohlfahrt, J.; Christens, P.; Olsen, J.; Melbye, M. Maternal age and fetal loss: Population-based register linkage study. BMJ 2000, 320, 1708–1712. [Google Scholar] [CrossRef]
- Du Fosse, N.A.; van der Hoorn, M.P.; van Lith, J.M.M.; le Cessie, S.; Lashley, E. Advanced paternal age is associated with an increased risk of spontaneous miscarriage: A systematic review and meta-analysis. Hum. Reprod. Update 2020, 26, 650–669. [Google Scholar] [CrossRef]
- Hertig, A.T.; Rock, J.; Adams, E.C. A description of 34 human ova within the first 17 days of development. Am. J. Anat. 1956, 98, 435–493. [Google Scholar] [CrossRef]
- Hertig, A.T.; Rock, J.; Adams, E.C.; Menkin, M.C. Thirty-four fertilized human ova, good, bad and indifferent, recovered from 210 women of known fertility: A study of biologic wastage in early human pregnancy. Pediatrics 1959, 23 Pt 2, 202–211. [Google Scholar] [CrossRef]
- Hertig, A.T.; Rock, J. Searching for early fertilized human ova. Gynecol. Investig. 1973, 4, 121–139. [Google Scholar] [CrossRef] [PubMed]
- Roberts, C.J.; Lowe, C.R. Where have all the conceptions gone? Lancet 1975, 305, 498–499. [Google Scholar] [CrossRef]
- Shepard, T.H.; Fantel, A.G. Embryonic and early fetal loss. Clin. Perinatol. 1979, 6, 219–243. [Google Scholar] [CrossRef]
- Morton, H.; Tinneberg, H.R.; Rolfe, B.; Wolf, M.; Mettler, L. Rosette inhibition test: A multicentre investigation of early pregnancy factor in humans. J. Reprod. Immunol. 1982, 4, 251–261. [Google Scholar] [CrossRef]
- Rolfe, B.E. Detection of fetal wastage. Fertil. Steril. 1982, 37, 655–660. [Google Scholar] [CrossRef] [PubMed]
- Smart, Y.C.; Fraser, I.S.; Roberts, T.K.; Clancy, R.L.; Cripps, A.W. Fertilization and early pregnancy loss in healthy women attempting conception. Clin. Reprod. Fertil. 1982, 1, 177–184. [Google Scholar]
- Wilcox, A.J.; Weinberg, C.R.; O’Connor, J.F.; Baird, D.D.; Schlatterer, J.P.; Canfield, R.E.; Armstrong, E.G.; Nisula, B.C. Incidence of early loss of pregnancy. N. Engl. J. Med. 1988, 319, 189–194. [Google Scholar] [CrossRef]
- Olsen, J.; Rachootin, P. Invited commentary: Monitoring fecundity over time—If we do it, then let’s do it right. Am. J. Epidemiol. 2003, 157, 94–97. [Google Scholar] [CrossRef]
- Wood, J.W. Fecundity and natural fertility in humans. Oxf. Rev. Reprod. Biol. 1989, 11, 61–109. [Google Scholar]
- Konishi, S.; Kariya, F.; Hamasaki, K.; Takayasu, L.; Ohtsuki, H. Fecundability and Sterility by Age: Estimates Using Time to Pregnancy Data of Japanese Couples Trying to Conceive Their First Child with and without Fertility Treatment. Int. J. Environ. Res. Public. Health. 2021, 18, 5486. [Google Scholar] [CrossRef]
- Henripin, J. La Population Canadienne au début du XVIIIème Siècle [Canadian Population at the Beginning of the XVIII Century]; Presses Universitaires de France: Paris, France, 1954. [Google Scholar]
- Wang, X.; Chen, C.; Wang, L.; Chen, D.; Guang, W.; French, J. Conception, early pregnancy loss, and time to clinical pregnancy: A population-based prospective study. Fertil. Steril. 2003, 79, 1517–1521. [Google Scholar] [CrossRef] [PubMed]
- Charbonneaux, P. Tourouvre-au-Perche au XVIIème et XVIIIème Siècle [Tourouvre-au-Perche during the XVII and XVIII Centuries]; Presses Universitaires de France: Paris, France, 1970. [Google Scholar]
- Vessey, M.; Doll, R.; Peto, R.; Johnson, B.; Wiggins, P. A long-term follow-up study of women using different methods of contraception. An interim Report. J. Biosoc. Sci. 1976, 8, 373–427. [Google Scholar] [CrossRef] [PubMed]
- Balakrishnan, T.R. Probability of conception, conception delay and estimates of fecundability in rural and semi-urban areas of certain Latin-American countries. Soc. Biol. 1979, 26, 226–231. [Google Scholar] [CrossRef] [PubMed]
- van Noord-Zaadstra, B.M.; Looman, C.W.; Alsbach, H.; Habbema, J.D.; te Velde, E.R.; Karbaat, J. Delaying childbearing: Effect of age on fecundity and outcome of pregnancy. Br. Med. J. 1991, 302, 1361–1365. [Google Scholar] [CrossRef] [PubMed]
- Sheps, M.C. An analysis of reproductive patterns in an American isolate. Popul. Stud. 1965, 19, 65–77. [Google Scholar] [CrossRef]
- Zinaman, M.J.; Clegg, E.D.; Brown, C.C.; O’Connor, J.; Selevan, S.G. Estimates of human fertility and pregnancy loss. Fertil. Steril. 1996, 65, 503–509. [Google Scholar] [CrossRef]
- Smarr, M.M.; Sapra, K.J.; Gemmill, A.; Kahn, L.G.; Wise, L.A.; Lynch, C.D.; Factor-Litvak, P.; Mumford, S.L.; Skakkebaek, N.E.; Slama, R.; et al. Is human fecundity changing? A discussion of research and data gaps precluding us from having an answer. Hum. Reprod. 2017, 32, 499–504. [Google Scholar] [CrossRef]
- Ellish, N.J.; Saboda, K.; O’Connor, J.; Nasca, P.C.; Stanek, E.J.; Boyle, C. A prospective study of early pregnancy loss. Hum. Reprod. 1996, 11, 406–412. [Google Scholar] [CrossRef]
- Leridon, H. Human fecundity: Situation and outlook. Popul. Soc. 2010, 471, 1–4. [Google Scholar]
- Simon, C.; Landeras, J.; Zuzuarregui, J.L.; Martin, J.C.; Remohi, J.; Pellicer, A. Early pregnancy losses in in vitro fertilization and oocyte donation. Fertil. Steril. 1999, 72, 1061–1065. [Google Scholar] [CrossRef]
- Boomsma, C.M.; Kavelaars, A.; Eijkemans, M.J.; Lentjes, E.G.; Fauser, B.C.; Heijnen, C.J.; Macklon, N.S. Endometrial secretion analysis identifies a cytokine profile predictive of pregnancy in IVF. Hum. Reprod. 2009, 24, 1427–1435. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, G.E. Misjudging early embryo mortality in natural human reproduction. F1000Res. 2020, 9, 702. [Google Scholar] [CrossRef] [PubMed]
- Kennedy, T.G. Physiology of implantation. In In Vitro Fertilization and Assisted Reproduction; Gomel, V., Leung, P.C.K., Eds.; Monduzzi Editore: Bologna, Italy, 1997; pp. 729–735. [Google Scholar]
- Jones, R.E.; Lopez, K.H. Pregnancy. In Human Reproductive Biology, 4th ed.; Jones, R.E., Lopez, K.H., Eds.; Elsevier: Amsterdam, The Netherlands, 2014; pp. 175–204. [Google Scholar]
- Sharkey, A.M.; Macklon, N.S. The science of implantation emerges blinking into the light. Reprod. Biomed. Online 2013, 27, 453–460. [Google Scholar] [CrossRef]
- Short, R.V. When a conception fails to become a pregnancy. Ciba Found. Symp. 1978, 64, 377–394. [Google Scholar]
- Wilcox, A.J.; Harmon, Q.; Doody, K.; Wolf, D.P.; Adashi, E.Y. Preimplantation loss of fertilized human ova: Estimating the unobservable. Hum. Reprod. 2020, 35, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Makrigiannakis, A.; Vrekoussis, T.; Zoumakis, E.; Kalantaridou, S.N.; Jeschke, U. The Role of HCG in Implantation: A Mini-Review of Molecular and Clinical Evidence. Int. J. Mol. Sci. 2017, 18, 1305. [Google Scholar] [CrossRef]
- Kim, S.M.; Kim, J.S. A Review of Mechanisms of Implantation. Dev. Reprod. 2017, 21, 351–359. [Google Scholar] [CrossRef]
- Macklon, N.S.; Brosens, J.J. The human endometrium as a sensor of embryo quality. Biol. Reprod. 2014, 91, 98. [Google Scholar] [CrossRef]
- Brosens, J.J.; Bennett, P.R.; Abrahams, V.M.; Ramhorst, R.; Coomarasamy, A.; Quenby, S.; Lucas, E.S.; McCoy, R.C. Maternal selection of human embryos in early gestation: Insights from recurrent miscarriage. Semin. Cell Dev. Biol. 2022, 131, 14–24. [Google Scholar] [CrossRef]
- Brosens, J.J.; Salker, M.S.; Teklenburg, G.; Nautiyal, J.; Salter, S.; Lucas, E.S.; Steel, J.H.; Christian, M.; Chan, Y.-W.; Boomsma, C.M.; et al. Uterine selection of human embryos at implantation. Sci. Rep. 2014, 4, 3894. [Google Scholar] [CrossRef]
- Shutt, D.A.; Lopata, A. The secretion of hormones during the culture of human preimplantation embryos with corona cells. Fertil. Steril. 1981, 35, 413–416. [Google Scholar] [CrossRef]
- Morton, H.; Rolfe, B.; Clunie, G.J. An early pregnancy factor detected in human serum by the rosette inhibition test. Lancet 1977, 1, 394–397. [Google Scholar] [CrossRef]
- Morton, H. Early pregnancy factor: An extracellular chaperonin 10 homologue. Immunol. Cell Biol. 1998, 76, 483–496. [Google Scholar] [CrossRef]
- Nahhas, F.; Barnea, E. Human embryonic origin early pregnancy factor before and after implantation. Am. J. Reprod. Immunol. 1990, 22, 105–108. [Google Scholar] [CrossRef]
- Barnea, E.R.; Lahijani, K.I.; Roussev, R.; Barnea, J.D.; Coulam, C.B. Use of lymphocyte platelet bindingassay for detecting a preimplantation factor: A quantitative assay. Am. J. Reprod. Immunol. 1994, 32, 133–138. [Google Scholar] [CrossRef]
- Barnea, E.R.; Simon, J.; Levine, S.P.; Coulam, C.B.; Taliadouros, G.S.; Leavis, P.C. Progress in characterization of pre-implantation factor in embryo cultures and in vivo. Am. J. Reprod. Immunol. 1999, 42, 95–99. [Google Scholar]
- Yang, M.; Yang, Y.; She, S.; Li, S. Proteomic investigation of the effects of preimplantation factor on human embryo implantation. Mol. Med. Rep. 2018, 17, 3481–3488. [Google Scholar] [CrossRef]
- Zare, F.; Seifati, S.M.; Mahdi Dehghan-Manshadi, M.; Fesahat, F. Preimplantation Factor (PIF): A peptide with various functions. JBRA Assist. Reprod. 2020, 24, 214–218. [Google Scholar] [CrossRef]
- Dos Santos, E.; Moindjie, H.; Sérazin, V.; Arnould, L.; Rodriguez, Y.; Fathallah, K.; Barnea, E.R.; Vialard, F.; Dieudonné, M.-N. Preimplantation factor modulates trophoblastic invasion throughout the decidualization of human endometrial stromal cells. Reprod. Biol. Endocrinol. 2021, 19, 96. [Google Scholar] [CrossRef]
- O’Neill, C.; Collier, M.; Ryan, J.P.; Spinks, N.R. Embryo-derived platelet-activating factor. J. Reprod. Fertil. Suppl. 1989, 37, 19–27. [Google Scholar]
- Roudebush, W.E.; Wininger, J.D.; Jones, A.E.; Wright, G.; Toledo, A.A.; Kort, H.I.; Massey, J.B.; Shapiro, D.B. Embryonic platelet-activating factor: An indicator of embryo viability. Hum. Reprod. 2002, 17, 1306–1310. [Google Scholar] [CrossRef] [PubMed]
- Velasquez, L.A.; Maisey, K.; Fernandez, R.; Valdes, D.; Cardenas, H.; Imarai, M.; Delgado, J.; Aguilera, J.; Croxatto, H.B. PAF receptor and PAF acetylhydrolase expression in the endosalpinx of the human Fallopian tube: Possible role of embryo-derived PAF in the control of embryo transport to the uterus. Hum. Reprod. 2001, 16, 1583–1587. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, C. The role of PAF in embryo physiology. Hum. Reprod. Update. 2005, 11, 215–228. [Google Scholar] [CrossRef] [PubMed]
- Sato, Y.; Fujiwara, H.; Konishi, I. Role of platelets in placentation. Med. Mol. Morphol. 2010, 43, 129–133. [Google Scholar] [CrossRef]
- Palomares, K.T.; Parobchak, N.; Ithier, M.C.; Aleksunes, L.M.; Castaño, P.M.; So, M.; Faro, R.; Heller, D.; Wang, B.; Rosen, T. Fetal Exosomal Platelet-activating Factor Triggers Functional Progesterone Withdrawal in Human Placenta. Reprod. Sci. 2021, 28, 252–262. [Google Scholar] [CrossRef]
- Fishel, S.B.; Edwards, R.G.; Evans, C.J. Human chorionic gonadotropin secreted by preimplantation embryos cultured in vitro. Science 1984, 223, 816–818. [Google Scholar] [CrossRef]
- Bonduelle, M.L.; Dodd, R.; Liebaers, I.; Van Steirteghem, A.; Williamson, R.; Akhurst, R. Chorionic gonadotrophin-beta mRNA, a trophoblast marker, is expressed in human 8-cell embryos derived from tripronucleate zygotes. Hum. Reprod. 1988, 3, 909–914. [Google Scholar] [CrossRef]
- Ochoa-Bernal, M.A.; Fazleabas, A.T. Physiologic Events of Embryo Implantation and Decidualization in Human and Non-Human Primates. Int. J. Mol. Sci. 2020, 21, 1973. [Google Scholar] [CrossRef]
- You, Y.; Stelzl, P.; Joseph, D.N.; Aldo, P.B.; Maxwell, A.J.; Dekel, N.; Liao, A.; Whirledge, S.; Mor, G. TNF-alpha Regulated Endometrial Stroma Secretome Promotes Trophoblast Invasion. Front. Immunol. 2021, 12, 737401. [Google Scholar] [CrossRef]
- Zhang, Z.; Yang, Y.; Lv, X.; Liu, H. Interleukin-17 promotes proliferation, migration, and invasion of trophoblasts via regulating PPAR-γ/RXR-α/Wnt signaling. Bioengineered 2022, 13, 1224–1234. [Google Scholar] [CrossRef]
- Chi, F.; Sharpley, M.S.; Nagaraj, R.; Sen Roy, S.; Banerjee, U. Glycolysis-Independent Glucose Metabolism Distinguishes TE from ICM Fate during Mammalian Embryogenesis. Dev. Cell 2020, 53, 9–26.e4. [Google Scholar] [CrossRef] [PubMed]
- Clark, D.A. Is there any evidence for immunologically mediated or immunologically modifiable early pregnancy failure? J. Assist. Reprod. Genet. 2003, 20, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Clark, D.A. Immunological factors in pregnancy wastage: Fact or fiction. Am. J. Reprod. Immunol. 2008, 59, 277–300. [Google Scholar] [CrossRef] [PubMed]
- Haddad, E.K.; Duclos, A.J.; Antecka, E.; Lapp, W.S.; Baines, M.G. Role of interferon-gamma in the priming of decidual macrophages for nitric oxide production and early pregnancy loss. Cell Immunol. 1997, 181, 68–75. [Google Scholar] [CrossRef] [PubMed]
- Haddad, E.K.; Duclos, A.J.; Lapp, W.S.; Baines, M.G. Early embryo loss is associated with the prior expression of macrophage activation markers in the decidua. J. Immunol. 1997, 158, 4886–4892. [Google Scholar] [CrossRef] [PubMed]
- Vento-Tormo, R.; Efremova, M.; Botting, R.A.; Turco, M.Y.; Vento-Tormo, M.; Meyer, K.B.; Park, J.-E.; Stephenson, E.; Polański, K.; Goncalves, A.; et al. Single-cell reconstruction of the early maternal-fetal interface in humans. Nature 2018, 563, 347–353. [Google Scholar] [CrossRef] [PubMed]
- Nancy, P.; Tagliani, E.; Tay, C.S.; Asp, P.; Levy, D.E.; Erlebacher, A. Chemokine gene silencing in decidual stromal cells limits T cell access to the maternal-fetal interface. Science 2012, 336, 1317–1321. [Google Scholar] [CrossRef]
- Ticconi, C.; Pietropolli, A.; Di Simone, N.; Piccione, E.; Fazleabas, A. Endometrial Immune Dysfunction in Recurrent Pregnancy Loss. Int. J. Mol. Sci. 2019, 20, 5332. [Google Scholar] [CrossRef]
- Robertson, S.A.; Care, A.S.; Moldenhauer, L.M. Regulatory T cells in embryo implantation and the immune response to pregnancy. J. Clin. Investig. 2018, 128, 4224–4235. [Google Scholar] [CrossRef]
- Kanamori, M.; Nakatsukasa, H.; Okada, M.; Lu, Q.; Yoshimura, A. Induced Regulatory T Cells: Their Development, Stability, and Applications. Trends Immunol. 2016, 37, 803–811. [Google Scholar] [CrossRef]
- Josefowicz, S.Z.; Rudensky, A. Control of regulatory T cell lineage commitment and maintenance. Immunity 2009, 30, 616–625. [Google Scholar] [CrossRef] [PubMed]
- Vent-Schmidt, J.; Han, J.M.; MacDonald, K.G.; Levings, M.K. The role of FOXP3 in regulating immune responses. Int. Rev. Immunol. 2014, 33, 110–128. [Google Scholar] [CrossRef] [PubMed]
- Sadlon, T.; Brown, C.Y.; Bandara, V.; Hope, C.M.; Schjenken, J.E.; Pederson, S.M.; Breen, J.; Forrest, A.; Beyer, M.; Robertson, S.; et al. Unravelling the molecular basis for regulatory T-cell plasticity and loss of function in disease. Clin. Transl. Immunol. 2018, 7, e1011. [Google Scholar] [CrossRef] [PubMed]
- Barnea, E.R. Embryo maternal dialogue: From pregnancy recognition to proliferation control. Early Pregnancy 2001, 5, 65–66. [Google Scholar]
- Csapo, A.I.; Pulkkinen, M.O.; Ruttner, B.; Sauvage, J.P.; Wiest, W.G. The significance of the human corpus luteum in pregnancy maintenance. I. Preliminary studies. Am. J. Obstet. Gynecol. 1972, 112, 1061–1067. [Google Scholar] [CrossRef]
- Jurisicova, A.; Varmuza, S.; Casper, R.F. Programmed cell death and human embryo fragmentation. Mol. Hum. Reprod. 1996, 2, 93–98. [Google Scholar] [CrossRef]
- Cecchele, A.; Cermisoni, G.C.; Giacomini, E.; Pinna, M.; Vigano, P. Cellular and Molecular Nature of Fragmentation of Human Embryos. Int. J. Mol. Sci. 2022, 23, 1349. [Google Scholar] [CrossRef]
- Hardy, K. Apoptosis in the human embryo. Rev. Reprod. 1999, 4, 125–134. [Google Scholar] [CrossRef]
- Leidenfrost, S.; Boelhauve, M.; Reichenbach, M.; Gungor, T.; Reichenbach, H.D.; Sinowatz, F.; Wolf, E.; Habermann, F.A. Cell arrest and cell death in mammalian preimplantation development: Lessons from the bovine model. PLoS ONE 2011, 6, e22121. [Google Scholar] [CrossRef]
- Liu, H.C.; He, Z.Y.; Mele, C.A.; Veeck, L.L.; Davis, O.; Rosenwaks, Z. Expression of apoptosis-related genes in human oocytes and embryos. J. Assist. Reprod. Genet. 2000, 17, 521–533. [Google Scholar] [CrossRef]
- Jurisicova, A.; Antenos, M.; Varmuza, S.; Tilly, J.L.; Casper, R.F. Expression of apoptosis-related genes during human preimplantation embryo development: Potential roles for the Harakiri gene product and Caspase-3 in blastomere fragmentation. Mol. Hum. Reprod. 2003, 9, 133–141. [Google Scholar] [CrossRef] [PubMed]
- Jurisicova, A.; Acton, B.M. Deadly decisions: The role of genes regulating programmed cell death in human preimplantation embryo development. Reproduction 2004, 128, 281–291. [Google Scholar] [CrossRef] [PubMed]
- Acton, B.M.; Jurisicova, A.; Jurisica, I.; Casper, R.F. Alterations in mitochondrial membrane potential during preimplantation stages of mouse and human embryo development. Mol. Hum. Reprod. 2004, 10, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Komatsu, K.; Iwase, A.; Mawatari, M.; Wang, J.; Yamashita, M.; Kikkawa, F. Mitochondrial membrane potential in 2-cell stage embryos correlates with the success of preimplantation development. Reproduction 2014, 147, 627–638. [Google Scholar] [CrossRef]
- Zhao, N.; Zhang, Y.; Liu, Q.; Xiang, W. Mfn2 Affects Embryo Development via Mitochondrial Dysfunction and Apoptosis. PLoS ONE 2015, 10, e0125680. [Google Scholar] [CrossRef]
- Filadi, R.; Pendin, D.; Pizzo, P. Mitofusin 2: From functions to disease. Cell Death Dis. 2018, 9, 330. [Google Scholar] [CrossRef]
- Haouzi, D.; Boumela, I.; Chebli, K.; Hamamah, S. Global, Survival, and Apoptotic Transcriptome during Mouse and Human Early Embryonic Development. Biomed. Res. Int. 2018, 2018, 5895628. [Google Scholar] [CrossRef]
- Amitai, T.; Kan-Tor, Y.; Or, Y.; Shoham, Z.; Shofaro, Y.; Richter, D.; Har-Vardi, I.; Ben-Meir, A.; Srebnik, N.; Buxboim, A. Embryo classification beyond pregnancy: Early prediction of first trimester miscarriage using machine learning. J. Assist. Reprod. Genet. 2023, 40, 309–322. [Google Scholar] [CrossRef]
- Orvieto, R.; Shimon, C.; Rienstein, S.; Jonish-Grossman, A.; Shani, H.; Aizer, A. Do human embryos have the ability of self-correction? Reprod. Biol. Endocrinol. 2020, 18, 98. [Google Scholar] [CrossRef]
- Monsivais, D.; Clementi, C.; Peng, J.; Titus, M.M.; Barrish, J.P.; Creighton, C.J.; Lydon, J.P.; DeMayo, F.J.; Matzuk, M.M. Uterine ALK3 is essential during the window of implantation. Proc. Natl. Acad. Sci. USA 2016, 113, E387–E395. [Google Scholar] [CrossRef]
- Chung, K.; Sammel, M.D.; Coutifaris, C.; Chalian, R.; Lin, K.; Castelbaum, A.J.; Freedman, M.F.; Barnhart, K.T. Defining the rise of serum HCG in viable pregnancies achieved through use of IVF. Hum. Reprod. 2006, 21, 823–828. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO Model List of Essential Medicines—22nd List. 2021. Available online: https://www.who.int/publications/i/item/WHO-MHP-HPS-EML-2021.02 (accessed on 15 September 2023).
- American College of Obstetricians, Gynecologists. Committee on Practice B-G. ACOG Practice Bulletin No. 200: Early Pregnancy Loss. Obstet. Gynecol. 2018, 132, e197–e207. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Population Studied | Publication | Fecundity Index |
|---|---|---|
| Canada (Québec), 18th century | Henrypin (1954) [22] | 0.31 |
| China 20th century | Wang et al (2003) [23] | 0.40 |
| France, 17th and 18th centuries | Charbonneaux (1970) [24] | 0.21 |
| Great Britain, 20th century | Vessey et al. (1976) [25] | 0.21 |
| Mexico, 20th century | Balakrishnan (1979) [26] | 0.21 |
| Peru, 20th century | Balakrishnan (1979) [26] | 0.17 |
| The Netherlands, 20th century * | van Noord Zaadstra et al. (1991) [27] | 0.54 |
| USA (Hutterite sect) 20th century | Sheps (1965) [28] | 0.28 |
| USA, 20th century | Zinamen et al. (1996) [29] | 0.30 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benagiano, G.; Mancuso, S.; Guo, S.-W.; Di Renzo, G.C. Events Leading to the Establishment of Pregnancy and Placental Formation: The Need to Fine-Tune the Nomenclature on Pregnancy and Gestation. Int. J. Mol. Sci. 2023, 24, 15420. https://doi.org/10.3390/ijms242015420
Benagiano G, Mancuso S, Guo S-W, Di Renzo GC. Events Leading to the Establishment of Pregnancy and Placental Formation: The Need to Fine-Tune the Nomenclature on Pregnancy and Gestation. International Journal of Molecular Sciences. 2023; 24(20):15420. https://doi.org/10.3390/ijms242015420
Chicago/Turabian StyleBenagiano, Giuseppe, Salvatore Mancuso, Sun-Wei Guo, and Gian Carlo Di Renzo. 2023. "Events Leading to the Establishment of Pregnancy and Placental Formation: The Need to Fine-Tune the Nomenclature on Pregnancy and Gestation" International Journal of Molecular Sciences 24, no. 20: 15420. https://doi.org/10.3390/ijms242015420