Abstract
As things stand in 2023, metastatic osteosarcoma commonly results in death. There has been little treatment progress in recent decades. To redress the poor prognosis of metastatic osteosarcoma, the present regimen, OSR9, uses nine already marketed drugs as adjuncts to current treatments. The nine drugs in OSR9 are: (1) the antinausea drug aprepitant, (2) the analgesic drug celecoxib, (3) the anti-malaria drug chloroquine, (4) the antibiotic dapsone, (5) the alcoholism treatment drug disulfiram, (6) the antifungal drug itraconazole, (7) the diabetes treatment drug linagliptin, (8) the hypertension drug propranolol, and (9) the psychiatric drug quetiapine. Although none are traditionally used to treat cancer, all nine have attributes that have been shown to inhibit growth-promoting physiological systems active in osteosarcoma. In their general medicinal uses, all nine drugs in OSR9 have low side-effect risks. The current paper reviews the collected data supporting the role of OSR9.
1. Introduction
Preface: In trying to hold onto everything, one holds onto nothing.King Fredrick the Great, 1712–1786
As things stand in 2023, metastatic osteosarcoma (OS) is often fatal within a few years of diagnosis. Extensive local bone destruction, aggressive surrounding soft tissue infiltration, osteoid deposition, and early metastases to the lungs are characteristic features of OS.
Predisposing factors are preexisting Paget’s disease or a prior cancer. The OS primary tumor is highly infiltrative. Pulmonary metastases are frequent and worsen prognosis. Although still not a common tumor, its incidence has been steadily increasing over recent decades. There has been little progress in improving survival in metastatic OS in the last 40 years, and five-year survival remains at ~30% [1]. OSR9 is a new approach to this problem. This paper presents the data outlining the rationale behind OSR9 treatment and for using a repurposed multidrug adjunctive regimen generally by adding it to current treatment options in metastatic OS.
The OSR9 regimen uses nine non-oncology drugs that have been repurposed as adjuncts to standard current OS treatments: (1) the anti-nausea drug aprepitant, (2) the analgesic drug celecoxib, (3) the anti-malaria drug chloroquine, (4) the anti-malaria drug dapsone, (5) the alcoholism treatment drug disulfiram, (6) the antifungal drug itraconazole, (7) the anti-diabetes drug linagliptin, (8) the antihypertension drug propranolol, and (9) the psychiatric drug quetiapine. These drugs and the data showing evidence of their ability to interfere with OS growth are discussed in detail below. See Table 1 for some of the core features of these OSR9 drugs’ attributes that are relevant in treating OS.

Table 1.
Overview of the nine OSR9 drugs (references are in the text). #, used in studies on the clinical treatment of glioblastoma; ALDH, aldehyde dehydrogenase; CA, carbonic anhydrase; COX, cyclooxygenase; GLP-1, glucagon-like peptide-1; DPP-4, dipeptidyl peptidase-4; Hh, Hedgehog; 5-LO, 5-lipoxygenase; RANK, receptor activator for nuclear factor-kappa B; RANKL, receptor activator for nuclear factor-kappa B ligand; TLR, toll-like receptor.
None of the OSR9 drugs have been proven to be of benefit during treatment for OS or any other cancer. However, since most physicians, regardless of specialty, are familiar with their use in their approved, non-oncology indications, all these OSR9 drugs are generally well-tolerated, and all nine drugs have preclinical evidence that they might retard OS growth, and so the risk/benefit ratio would favor a small study on this OSR9 regimen in metastatic OS patients for whom no other treatment options exist.
On X-ray plain films, OS displays the following features:
- Patchy cortical bone destruction
- Areas of sclerotic bone
- Wide and gradual transitions from normal-appearing trabecular bone to clearly abnormal, moth eaten, and irregularly mineralized bone
- Peritumoral soft tissue edema
- Codman’s triangle (irregular periosteum that has lifted off where advancing tumor margins destroy periosteal new bone formation faster than it can ossify)
OS is a tumor composed of mesenchymal cells held at an immature osteoblast differentiation stage. Local regions of genome hypermutations (kataegis) are present [2,3,4]. These regions of grossly disturbed genome show ploidy increases and decreases and chromosomal amplifications, losses, gains, and mutations. These genomic abnormalities are not stable either; they change rapidly. RUNX2, essential for the normal differentiation of bone and mammary tissues, have elevated copy numbers, RNA, and protein levels in OS tumors, but these are ineffective in driving differentiation as RUNX-2 would normally do in normal bone [2,3,4,5].
2. The Need for Both Repurposed and Multidrug Regimens in OS
There has been little progress in treating OS over the last 30+ years despite dozens of clinical trials of combinations of new and older cytotoxic drugs, tyrosine kinase receptor inhibitors, and monoclonal antibodies directed at putative growth factors [3,6,7]. A large peak OS incidence occurs between 5 and 25 years of age and a lesser peak incidence occurs between 75 and 85, with significantly poorer survival in the older group [6,7]. Overall, the five-year survival for OS in 0 to 25 year-olds was 36%; for 25 to 59 year-olds, it was 15%; and for >60 year-olds, it was 6% [6].
Twelve recent papers have presented the thinking about a new school of cancer treatment that espouses adding multiple repurposed non-oncology drugs to standard current cytotoxic chemotherapies. A series of recent papers from the IIAIGC Study Center [8,9,10,11] and from other centers [12,13,14,15,16,17,18,19] have outlined this reasoning for why such a multidrug regimen would be needed to treat common deadly metastatic cancers. Briefly summarized, the 12 papers outline these reasons as follows:
2.1. Why Many Drugs Are Needed
- Absent a “silver bullet”, many drugs may be needed to address tumors’ readily evolving and multiple resistance pathways and their ability to shift reliance to alternate growth drives when the one in use becomes blocked.
- The common metastatic deadly cancers have metabolic flexibility, shifting between aerobic oxphos, aerobic glycolysis, anaerobic glycolysis, beta-oxidation, autophagy, glutamine, and other energy sources as circumstances dictate.
- The common metastatic deadly cancers are hardy tissues with growth drive flexibility. The many alternate growth driving pathways can cross-cover for any one or two that become blocked.
- Inherent multiple subpopulations exist in these cancers, even before any treatment. Each subpopulation has its own particular set of growth drives and inhibition susceptibilities and resistances. Treatment-induced selection pressure favors the development of resistant clones.
- Ionizing irradiation and many common cytotoxic chemotherapies that are currently used to treat cancer are mutagens that hasten responses to selection pressure.
2.2. Why Repurposed Non-Oncology Drugs Are Needed
- We are constrained in treating today’s illnesses with today’s tools.
- Directly cytotoxic and genotoxic drugs have limits on how many, how much, and how often they can be used without destroying bone marrow or other essential body systems.
- Cancers, generally, and OS, specifically, use normal physiological growth drive pathways, but they pathologically and exaggeratedly engage them. We have many drugs in general medical practice with established safety for influencing those core mammalian systems.
- Repurposed drugs from general medical practice are cheap, generic, readily available, and well-tolerated, and GPs worldwide are already familiar with their use.
3. The OSR9 Drugs
The drugs in the OSR9 regimen are reviewed below. It might be thought of as curious that two of these repurposed drugs—chloroquine and dapsone—have decades of use in treating malaria and a third drug, itraconazole, is cytotoxic to Plasmodium falciparum in vitro at 1 microg/mL [20].
That three of the nine OSR9 drugs are cytotoxic to a protozoan becomes less strange in light of 2.2 number 3 above. Cancers, generally, and OS, specifically, use the same physiological systems that non-malignant tissues use in their day-to-day functioning. There is great commonality in physiological systems among Eukaryotes. They generally use the same or similar physiological systems.
Also not strange is the fact that the individual OSR9 drugs each have several seemingly unrelated effects on different cell physiological systems. Such many-target attributes of common drugs are common.
3.1. The Anti-Nausea Drug Aprepitant
Aprepitant is a generic, anti-nausea drug approved for use in treating cancer chemotherapy-related nausea and vomiting. It inhibits the NK-1 receptor for substance P (also called NK-1), an 11 amino-acid peptide [21,22]. Hundreds of clinical studies have attested to the safety and effectiveness of aprepitant in treating chemotherapy-related nausea and vomiting. Aprepitant is experiencing a resurgence of interest as an adjunct to retard growth across a variety of cancers [23,24,25,26,27]. Aprepitant has an eminently benign side effect profile.
In the past decade, dozens of papers have attested to the expression of, and growth-metastasis drive contributions from, the NK-1 receptor across a variety of the common cancers [28,29,30,31,32,33,34,35,36,37,38,39]. In the past decade, dozens of papers have shown that aprepitant mediates the in vitro arrest in growth and migration across the common human cancers [23,25,40,41,42,43,44,45,46,47,48,49,50,51,52].
Specifically in OS, recent reviews have presented the rationale for using aprepitant to retard growth [53,54,55]. OS cells express NK-1 mRNA and the NK-1 receptor, suggesting an autocrine growth drive [53,54,55,56]. Aprepitant inhibited the growth of OS in a murine xenotransplant model [53].
Despite this wealth of evidence that NK-1/substance P signaling forms a growth-driving element across a wide variety of cancers, aprepitant’s only clinical use so far has been its use in treating glioblastoma where preliminary evidence indicates a survival advantage from its use in the ten-drug CUSP9v3 protocol for treating glioblastoma [9].
3.2. The Analgesic Celecoxib
Celecoxib is a cyclooxygenase (COX) inhibitor selective for the COX-2 isoform. Celecoxib is commonly used to treat pain of diverse origins [57,58]. It lacks the platelet aggregation inhibitory activity seen with some other COX inhibitors [59], and it does not have the potential to create the gastric ulcerations that are associated with other COX inhibitors [60,61].
Shortly after introduction to clinical practice in the symptomatic treatment of pain, anti-cancer effects were noted both empirically and by theoretical reasoning [62,63]. Celecoxib is now being widely studied and even used on and off cancer treatment protocols in a variety of different cancers [64,65,66,67,68,69,70,71]. Celecoxib has demonstrated preclinical synergy with a variety of other traditionally used cytotoxic drugs in suppressing OS growth [72,73,74,75].
COX is a sine qua non enzyme in the synthesis of prostaglandin E. The COX-2 isoform is widely recognized as being upregulated, forming one of the many upregulated and pathophysiological-driving elements across the common cancers [68,76,77,78,79,80]. Specifically in OS, COX-2 overactivity has been repeatedly demonstrated in both pediatric and adult cases [81,82,83,84,85,86,87]. The degree of COX-2 immunohistochemical expression is inversely associated with OS patient survival duration, implying a growth-promoting role for COX-2 [86].
Human OS tissues heavily immunostain positive for COX-2 [88]. COX-2 has been demonstrated to be an in vitro driver of OS proliferation, migration, and cell survival [89]. Of note, celecoxib must be dosed at a minimum of 600 mg p.o. bid for the adequate inhibition of COX-2 when treating any cancer [90]. Lower doses are effective when treating pain.
Carbonic anhydrase (CA) catalyzes the reaction as follows:
CO2 + H2O → H+ and bicarbonate.
CA has 12 isoforms, some are soluble in cytoplasm (CAII), while some are transmembrane (CAIX and CAXII. The net effect is the further acidification of the extracellular milieu. OS, similar to other cancers, grows with a disorganized architecture, abnormally functioning, malformed vasculature, and a stroma that impedes diffusion. CO2 (pKa 6.35) has less of an acidifying effect than lactic acid d (pKa 3.86) does. CO2 freely diffuses across outer cell membranes, bicarbonate and protons do not.
Celecoxib also inhibits CA [91,92,93]. There is a particularly important role for dual COX-2/CA inhibitors in the treatment of cancers generally [94,95]. CAIX participates in acidification of the extracellular tumor milieu, with maintenance of an alkaline intracellular milieu with an enhanced growth vigor consequent to this proton efflux across a wide variety of common human cancers [96,97,98,99,100]. Also in OS, CA-mediated cellular proton efflux is an important homeostasis-maintaining accommodation to the malignancy-characteristic aerobic glycolysis that is present in OS [101,102,103,104,105,106]. Celecoxib has the potential to undermine this malignancy-related homeostatic pathway. Figure 1 shows the basic pathway of how CAII and CAIX mediate proton export and the maintenance of relatively alkaline cytosol [107].
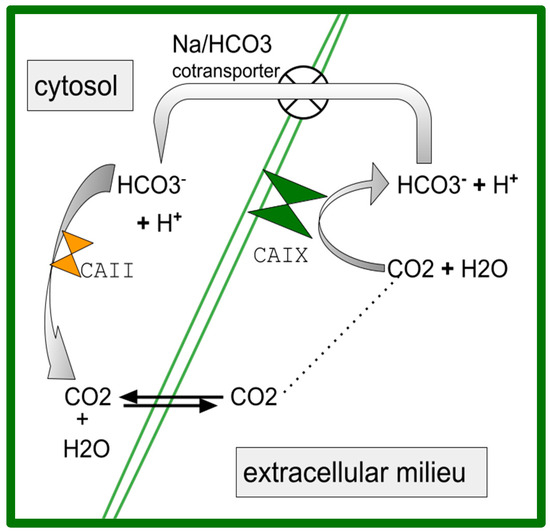
Figure 1.
A simplified schematic of the role of carbonic anhydrase (CA) in acidifying tumor extracellular space and maintaining a relatively basic cytosol [107]. CAIX is outer membrane bound acting in the extracellular space while CAII is cytosolic. Both are inhibited by celecoxib (references in Section 3.2).
Celecoxib performs the nanoM inhibition of CAIX [103]. CAIX participates in creating the characteristic OS migration, survival, and acidification of the extracellular milieu [108,109,110,111]. Higher CAIX expression is associated with shorter OS survival [112], which we also see in pancreatic ductal adenocarcinoma [113], glioblastoma [114], breast cancer [115], and other cancers [100].
3.3. The Antimalarial Drug Chloroquine
Chloroquine is a drug that was introduced to treat malaria in the mid-1940s, and it has been in continuous use for this since then. The mechanism of action in killing Plasmodia is by chlororoquine’s accumulation in the trophozoite’s acidic food vacuole, thereby preventing ingested hemoglobin degradation and/or by inhibiting the trophozoite’s heme polymerase [116,117,118]. Hydroxychloroquine and chloroquine are closely related and have similar uses in treating rheumatoid diseases such as rheumatoid arthritis, lupus, Sjögren’s, etc., as well as malaria. They both reduce mammalian lysosome function by increasing intralysosomal pH. They both result in clinically reduced inflammation-related cytokine production, inhibit cytokine production, and have a half-life of 40–60 days.
Their primary serious potential side effect of retinal injury is derived from their strong binding to melanin. For linguistic simplicity, the following discussion should be understood to refer to both when “chloroquine” is mentioned.
Much of the previous work on chloroquine as a cancer treatment adjunct generally [119,120,121,122,123], and specifically in OS [124], has been focused on chloroquine’s inhibition of autophagy. Indeed, chloroquine does this well and by known mechanisms [125]. This may be an important mode of OS growth inhibition, as these works outline. However, here, the focus is on chloroquine’s inhibition of inflammation-related cytokines and on its toll-like receptor-9 (TLR-9) inhibition.
TLR-9 is one of the many innate pathogen-recognizing systems cells have. It is activated by CpG DNA [126]. CpG DNA sites are cytosine-phosphate-guanine DNA sequence pairs that tend to occur grouped in localized DNA areas. Chloroquine inhibits TLR-9 signaling, as has been shown in diverse settings [127,128,129,130,131,132]. TLR-9 activation on normal osteoblasts by CpG DNA triggers, through intermediaries, nuclear NFkappaB activation and the expression of tumor necrosis factor-alpha (TNF) and macrophage-colony-stimulating factor (M-CSF) [133].
TLR-9 expression is increased in OS compared to normal osteoblasts [134]. The knockdown of TLR-9 inhibited OS growth and forced overexpression of TLR-9 increased OS growth in experimental models [135].
A similar pattern was seen in cholangiocarcinoma, where TLR-9 was overexpressed, yet it was absent in normal biliary tracts [136]. TLR-9 inhibition with chloroquine inhibited in vitro growth of cholangiocarcinoma and in xenografts [136]. The growth drive contributions of TLR-9 agonism have also been seen in the cells of breast [137] and esophagus [138] cancers.
3.4. The Antibiotic Dapsone
Dapsone is a sulfone antibiotic used to treat malaria, Hansen’s disease, tuberculosis, and other microbiological diseases. Similar to chloroquine, dapsone is effective for and used to treat rheumatological disease, particularly, rheumatoid arthritis [139,140,141].
IL-8 (also called CXCL8) is a neutrophil-attractant chemokine that signals via CXCR1 or CXCR2 receptors [142]. IL-8 is stored in neutrophils and the vascular endothelium.
Dapsone suppresses IL-8 synthesis [143,144,145]. Dapsone also suppresses neutrophil chemotaxis to IL-8 gradients [14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52,53,54,55,56,57,58,59,60,61,62,63,64,65,66,67,68,69,70,71,72,73,74,75,76,77,78,79,80,81,82,83,84,85,86,87,88,89,90,91,92,93,94,95,96,97,98,99,100,101,102,103,104,105,106,107,108,109,110,111,112,113,114,115,116,117,118,119,120,121,122,123,124,125,126,127,128,129,130,131,132,133,134,135,136,137,138,139,140,141,142,143,144,145,146,147,148]. Il-8 is an active participant in the joint-destructive inflammation of rheumatoid arthritis [149]. It is thought that dapsone reduces rheumatoid arthritis activity by reducing Il-8 and Il-8-mediated neutrophil accumulations. Dapsone also reduces mortality associated with COVID-19 and acute adult respiratory syndromes of any other origin by this same mechanism of action [150,151]. Neutrophil function is essential for life, but overly exuberant neutrophil function potentially creates tissue destruction and tumor trophic functions.
Exosomes from OS cells signal otherwise normal lung resident fibroblasts to secrete IL-8, facilitating the formation of the pulmonary metastases characteristic of OS [152,153]. IL-8 also forms one of the factors that trigger single OS cells to avoid anoikis [152,154]. OS patients have increased circulating IL-8 levels proportional to increased tumor burden [155,156,157,158,159]. Dogs with OS also have shown increased circulating IL-8 compared to normal dogs [160].
An interesting point about metastasis in cancer generally, and in OS specifically, is the extreme rarity of metastasis considering the millions of viable circulating tumor cells. There is experimental evidence in OS that the subset of circulating OS cells with greater IL-8 synthesis are the subset that can establish metastasis [152,155,159,161]. IL-8 signaling at CXCR1 is a cisplatin cytotoxicity resistance factor in OS [161].
In 2018, Kawano et al. provided evidence that an amplifying IL-8 feedback loop exists between OS cells and stroma, where the OS release of IL-8 triggers stroma cells’ synthesis of IL-8, which then triggers the OS cell to make more IL-8 [162]. This positive feedback loop is diagrammed in Figure 2. This work has been supported by the work of others [163]. Of great interest is that this same IL-8-amplifying feedback loop has been described as occurring in the generation of the erlotinib skin rash and its inhibition by dapsone [164].
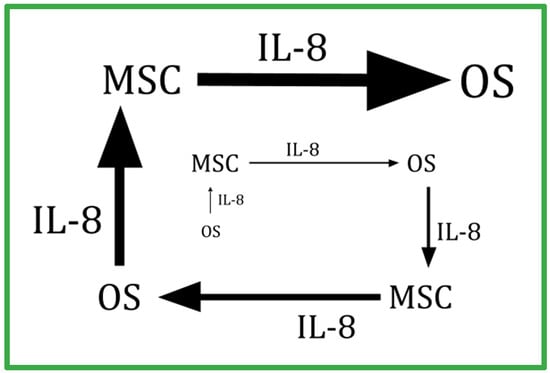
Figure 2.
Schematic of the amplifying feedback loop between nonmalignant mesenchymal cells and malignant osteosarcoma (OS) cells showing the OS-secreted IL-8 that stimulates stromal mesenchymal stem cells (MSC) that also synthesize IL-8 and are stimulated to make more IL-8 that, in turn, stimulates OS cells to make more IL-8. A similar IL-8-centered feedback loop exists between circulating neutrophils and a tumor’s endothelium has been described in other cancers (refs. [162,163,164]). Dapsone, by decreasing IL-8 and blunting cells’ responses to IL-8, interrupts this amplifying feedback loop.
There are, however, dozens or hundreds of additional attributes beyond IL-8 that favor or do not favor an OS cell’s ability or inability to establish metastasis.
3.5. The Alcoholism Treatment Drug Disulfiram
Disulfiram is a drug that has been used since the 1950s to treat alcoholism by making ethanol ingestion highly unpleasant. It is a potent inhibitor of all the isoforms of aldehyde dehydrogenase (ALDH). Disulfiram thereby stops ethanol metabolism at the acetaldehyde stage. Flushing, hypotension, malaise, nausea, and vomiting ensue due to acetaldehyde accumulation if ethanol is consumed.
The basic rationale for adding disulfiram to OS treatment is based on the association between high ALDH expression in individual cells and those cells having stem cell attributes. This holds true for the stem cell subpopulations within common cancers [165,166], including throughout the sarcomas [167] and, specifically, in OS [168,169,170,171,172,173]. There are currently over 30 open studies on disulfiram as an adjuvant treatment in various cancers that were all based on that association (clinicaltrials.gov).
Recent works have continued to support an adjunctive cancer growth suppression role for disulfiram [174,175,176,177,178].
3.6. The Antifungal Drug Itraconazole
Itraconazole is a broad-spectrum antifungal agent that has been used clinically since the late 1980s. It is commonly used today for minor illnesses such as onychomycosis [179], and for serious fungal infections [180], or as an antifungal prophylaxis for the immunosuppressed [181,182,183].
Itraconazole inhibits the function of several cell physiological systems that are known to facilitate or drive growth across a variety of cancers [184]. Specifically, itraconazole has inhibited Hedgehog (Hh) signaling [185], O-glycosylation [186], the drug efflux pump P-gp [187], and 5-lipoxygenase 5-LO [188].
Specifically in OS, Hh signaling drives increased growth vigor. Hh inhibition retards growth, and higher Hh expression is associated with shorter OS survival, increased radioresistance, and chemotherapy cytotoxicity resistance [189,190,191,192,193,194,195,196,197,198,199,200,201].
Clinically, survival benefit from adjunctive use of itraconazole, has been shown in several human cancers, including colon cancer [202], pancreatic ductal adenocarcinoma [203], gastric cancer [204], non-small-cell lung cancer [205], and epithelial ovarian cancer [206]. A retrospective analysis of patients with acute lymphoblastic and acute myelogenous leukemia receiving daunorubicin where itraconazole was used as antifungal prophylaxis during the neutropenic nadir showed better remission rates than those not receiving itraconazole [207]. In vitro, intracellular levels of daunorubicin have increased proportionately as itraconazole levels rose from 0.5 to 5.0 microg/mL [208]. Experimental (non-marketed) Hh inhibitors have impaired OS growth in preclinical OS model systems [209].
Clinically achieved levels of itraconazole block VEGF binding to VEGFR2. This was traced to defective receptor trafficking, which, in turn, was secondary to the itraconazole-mediated defective glycosylation of VEGFR2. Itraconazole disrupts the orderly formation of mammalian outer cell surface lipid rafts and, therefore, tends to inhibit cytotoxic mechanisms that depend on target molecule expression on/in lipid raft concentrated domains [210,211]. Itraconazole has significantly inhibited breast cancer resistance protein (BCRP), P-gp, and other drug efflux pumps [212,213,214,215]. By this mechanism, itraconazole has lowered resistance to topotecan [216], doxorubicin, and etoposide [217].
Recent works have continued to support the potential for itraconazole to inhibit cancer growth [218,219,220,221].
3.7. The Anti-Diabetes Drug Linagliptin
The gliptins are a group of related drugs marketed to treat type 2 diabetes, and they include alogliptin, anagliptin, evogliptin, gemigliptin, linagliptin, omarigliptin, sitagliptin, teneligliptin, trelagliptin, and vildagliptin. They all inhibit dipeptidyl peptidase-4 (DPP4, also termed CD26) [222]. DPP4 is a dimeric serine exopeptidase that cleaves at the penultimate proline or alanine from the N-terminus of a polypeptide. The cleaved polypeptide usually, but not always, loses signaling activity. DPP-4 is found in both soluble and membrane-bound forms. The DPP-4 inhibitor linagliptin, clinically marketed to treat type 2 diabetes, has inhibited two OS cell lines in vitro [223]. However, a concentration of 50 microM was used to show that. This raises uncertainty about the clinical potential of linagliptin in OS. Increased DPP4 activity has indirectly promoted bone resorption and inhibits bone formation [224].
RUNX2 is a transcription factor crucial for multipotent mesenchymal cells’ differentiation into immature osteoblasts. OS cases with greater RUNX2 overexpression have worse prognosis [225,226,227]. Runx2 must form a heterodimer, thereby to acquire DNA binding and to mediate osteoblast maturation. DPP4 is overexpressed in OS tissues compared to corresponding normal bone tissues, and greater overexpression is associated with shorter survival [228,229].
As diagrammed in Figure 3, gliptin DPP-4 inhibitors result in increasing AMPK phosphorylation. This is one of the osteoblast maturation pathways of gliptins. Gemigliptin has increased the level of P-AMPK and Akt phosphorylation in human umbilical vein endothelial cells and in macrophages [230]. Consequent to that, gemigliptin has also reduced the levels of LPS-induced vascular cell adhesion molecule-1 (VCAM-1), E-selectin, TNF, monocyte chemoattractant protein-1 (MCP-1), interleukin-1 beta, and IL-6 [230]. In mice fed a high fat diet, linagliptin plus metformin reversed the elevated TNF and IL-1 beta levels that are otherwise generated by this diet [231]. Other DPP-4-inhibiting gliptins, such as omarigliptin [232], anagliptin [233], and trelagliptin [234], have promoted the in vitro maturation of osteoblasts. Omarigliptin has also enhanced the conversion of AMPK to phosphorylated AMPK (P-AMPK) in vitro in renal glomerular cells [235]. Other gliptins have also mediated AMPK phosphorylation [236,237,238,239,240].
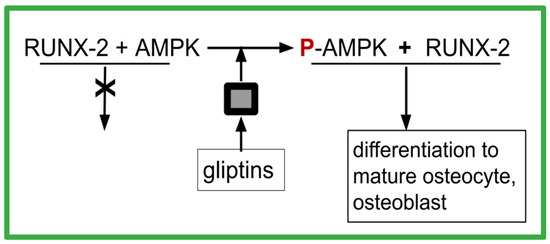
Figure 3.
How gliptins allow RUNX-2 to function as differentiation factor. The black box indicates missing steps between the presence of a gliptin and the phosphorylation of AMPK. The intermediates are unknown.
P-AMPK is required for RUNX-2 to function as a differentiation transcription factor, as depicted in Figure 3. This might account for the seemingly paradoxical situation where OS has excess RUNX-2 yet is deficient in differentiation to mature well-behaved osteoblasts. The excess DPP-4 also characteristic of OS prevents the differentiation function of RUNX-2 by preventing the P-AMPK necessary for RUNX-2 to function properly. A second indicator supporting such a conjecture was the observation from large meta-analyses of clinical trials of DPP-4 inhibitors that showed 40% reductions in fractures in those on DPP-4 inhibitors, which was indicative of improved osteoblast function [241].
MC3T3-E1 cells are a commercial murine cell line that can differentiate into osteoblasts and osteocytes. The addition of saxagliptin in vitro increased osteopontin (OPN), RUNX-2, and type I collagen in MC3T3-E1 cells and enhanced their differentiation into mature osteoblasts [242]. This differentiation mediation was dependent on RUNX-2 and on saxagliptin’s mediation of the conversion of AMPK to phosphorylated AMPK (P-AMPK) [232,242]. Another gliptin DPP-4 inhibitor, anagliptin, has also increased the differentiation of MC3T3-E1 into mineralized osteoblasts via the activation of RUNX2 [233].
The above references can be summarized as follows:
- OS overexpress RUNX-2, yet the malignant clone fails to mature
- OS overexpress DPP-4
- P-AMPK is required for RUNX-2 to function in osteoblast maturation
- gliptins promote AMPK phosphorylation
From this, we can then hypothesize that a core feature of OS is differentiation (maturation) failure due to RUNX-2 function failure due to DPP-4-mediated local unavailability of the required co-factor P-AMPK. This is schematically depicted in Figure 3.
If linagliptin or any other DPP-4 inhibitor is used during the course of OS, an appropriate PET scan should be completed before starting treatment, and then again after 20–40 days on the DPP-4 inhibitor. This requirement is due to elements of uncertainty about the role of DPP-4, its inhibition, and malignant growth [243].
3.8. The Antihypertension Drug Propranolol
Propranolol is the first beta adrenergic receptor blocking drug to be marketed. Introduced to clinical practice in the 1960s, it continues to be in common use for treating hypertension, migraines, or pediatric hemangioma [244].
Although little has been published on this aspect of propranolol, a pre-performance beta-adrenergic blockade with propranolol is in wide use in musicians, actors, public speakers, etc. [245,246,247].
Propranolol reduces the physical impediments to music performances (e.g., sweating, dry mouth, and hand tremors) without reducing mental acuity, performance capability, or emotional components. This reduction in the physical signs of sympathetic overdrive is of relevance to propranolol’s intended use during cancer and, specifically, OS treatment.
Propranolol’s sympathetic drive reduction by blocking the beta-adrenergic receptor as a cancer treatment adjunct, including in sarcomas and OS specifically, occurs through several putative mechanisms [247,248,249,250,251,252,253,254]. The primacy of any one of these paths has not been established.
Beta-adrenergic signaling is one of the cancer-related drivers of the elevated numbers of myeloid derived suppressor cells characteristically seen in most of the common cancers [255,256,257,258,259]. Beta-adrenergic signaling particularly contributes to conditioning lung tissues to facilitate metastasis establishment [260,261].
Normal bone is innervated by the sympathetic nervous system acting on beta-adrenergic receptors [262,263]. OS tissues overexpress the beta-adrenergic receptor [264]. Trabecular and cortical bone density is increased by a beta-adrenergic blockade and conversely decreased by an unchecked beta-adrenergic drive [265,266,267,268]. Specifically in OS, propranolol has retarded growth and increased cisplatin’s growth retardation in murine and canine OS models [269,270,271].
3.9. The Psychiatric Drug Quetiapine
Quetiapine is a drug used in psychiatry to reduce the signs and symptoms of psychosis, to augment an inadequate response to antidepressant drugs, and to stop disrupted sleep–wake cycles. It is also eminently safe to use in non-psychiatric populations.
Normal mammalian bones are constantly being absorbed by osteoclasts and resynthesized by osteoblasts throughout life, a process called remodeling [272]. Osteoclasts differentiate from hematopoietic progenitors, and osteoblasts differentiate from mesenchymal stem cells.
Receptor activator for nuclear factor-kappa B ligand (RANKL) and its decoy receptor osteoprotegerin (OPG), a soluble glycoprotein 60-kDa monomer, are expressed on osteoblasts while RANK is expressed on osteoclasts. OPG, the soluble RANKL decoy receptor, reduces RANKL-driven osteoclastogenesis by binding RANKL, limiting RANKL signaling to RANK.
The RANK/RANKL/OPG system lies at the core of osteosarcoma. In OS, this RANK–RANKL process participates in creating the characteristic patchy areas of bone resorption. Bone metastasis is common in other cancers where the RANK–RANKL system also is a core mediating system in perimetastasis bone destruction [273,274,275].
The immunohistochemical localization of RANK-positive cells in human OS biopsy tissues tend to be seen in osteoclasts at the tumor–bone interface and in intratumoral osteoclasts and in the myeloid osteoclast precursors more than in the OS cells themselves. OS cells themselves express RANKL [276]. RANK–RANKL signaling mediates the trophic function of OS stromal cells, facilitating or permitting the malignant cells themselves to thrive [276,277,278,279,280,281]. Larger total OS tumor tissue RANK expression has been associated with shorter OS survival.
RANKL binds to osteoclast the outer membrane receptor RANK on immature osteoclast precursors. This triggers widespread changes, including NFkB nuclear translocation, mitogen-activated protein kinase activations, activating protein 1 (AP-1), activating the nuclear factor of activated T cells 1 (NFAT-1), and activating phosphatidylinositol 3-kinase (PI3K)/Akt, ending in multinucleated, active osteoclasts. This is a key factor in immature preosteoclast cell differentiation and activation.
High-dose quetiapine (100 microM) is nontoxic to osteoclasts in vitro, yet even lower doses (25 microM) can suppress RANK–RANKL driven in vitro osteoclast maturation and function [282]. In a breast cancer metastasis murine model, quetiapine inhibited perimetastasis bone destruction [282]. Quietapine also inhibited RANKL-induced osteoclast differentiation in a mouse macrophage cell line with no sign of general cytotoxicity [282].
Mature oligodendrocytes make myelin and oligodendrocyte progenitor cells do not. A series of preclinical studies have shown that quetiapine stimulates the maturation of oligodendrocyte progenitor cells and their myelin synthesis [283,284,285,286,287,288]. Reflecting this improved myelination and oligodendrocyte maturation, a human study on mania showed that a year of quetiapine treatment repaired the impaired functional connectivity of brain regions characteristic of untreated mania [289].
Heterotopic hepatocellular carcinoma tumors grew more slowly in quetiapine treated mice, but the proximate mediating event of the growth inhibition was unknown [290].
We have no direct evidence that quetiapine will enhance malignant OS cell maturation or slow OS growth, but the above evidence allows a plausible pathway by which quetiapine might do this by depriving OS of the RANK–RANKL signaling that is active in the trophic nonmalignant cells within a tumor mass.
4. TICO
The neutrophil-to-lymphocyte ratio (NLR) is the ratio of the number of neutrophils compared to the number of lymphocytes in peripheral blood. A higher than normal NLR is a common finding across the common cancers [291,292,293,294]. The consensus finding across these studies is that as this ratio progressively exceeds 3:1, the prognosis decreases.
Fourteen independent clinical studies conducted specifically on OS have shown that the higher an NLR becomes, the shorter the survival becomes [294,295,296,297,298,299,300,301,302,303,304,305,306]. The extraordinary nature of this association of a higher NLR with a shorter survival in adult and pediatric OS should be fully appreciated. There is no other finding, in the entire body of oncology knowledge, that is so uniformly found across all the common cancers, generally, and as above, in OS, specifically.
TICO is a repurposed drug regimen designed to lower the NLR [44]. Given the strong association of a higher NLR with shorter survival in OS, the TICO regimen should be considered as a treatment adjunct in OS.
The TICO regimen has marshaled past research showing NLR reductions after individual exposure to tadalafil, isotretinoin, colchicine, and omega-3. Therefore, TICO proposes using all four during cancer treatment to improve prognosis by pharmacologically lowering NLRs. However, TICO has not yet been proven to be safe or effective.
5. Discussion and Conclusions
This paper discusses several pathogenic mediators in OS and potential already marketed repurposed drugs to inhibit these systems, but it is important to remember that many more pathogenic physiological derangements are present in OS beyond those discussed here [196,307,308,309,310]. As with other deadly cancers, the malignant behavior of OS cells involves deranged activation and/or pathological drop-out and/or the pathologically active participation of dozens (or hundreds) of cell-signaling systems. The drugs and the pathology-driving systems they inhibit, as mentioned in this paper on OSR9, are the systems we might currently be able to easily inhibit. How many pathology-driving systems must we inhibit to stop OS growth? We do not know, but treatment with OSR9 is a start.
There have been no controlled trials of the OSR9 drugs. For the reasons listed in Section 2.1 and Section 2.2 above, we should not expect meaningful effects from any one or two drug additions to the current treatments. Hence, CUSP9v3 and the current OSR9 have nine or more drugs.
A current trial of adjuvant hydroxychloroquine with docetaxel and gemcitabine (NCT03598595) was started in June of 2023. We may not achieve positive results in this trial. The need for a many-drug regimen and the reasons why these many drugs must be taken from general medicine, as outlined in Section 2 of this paper, represent the thinking of a new school of oncology. Although currently still a minority among oncologists and researchers, this movement is gaining adherents. OSR9 is a further contribution to this trend.
As things currently stand in 2023, and given the likely outcome of metastatic OS and the likely benign side effects expected from the OSR9 medicines, this regimen might be worth a small pilot study.
Funding
This was unfunded research carried out under the aegis of the IIAIGC Study Center.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
All data has been presented in the paper.
Conflicts of Interest
The author declares no conflict of interest.
References
- Panez-Toro, I.; Muñoz-García, J.; Vargas-Franco, J.W.; Renodon-Cornière, A.; Heymann, M.-F.; Lézot, F.; Heymann, D. Advances in Osteosarcoma. Curr. Osteoporos. Rep. 2023, 21, 330–343. [Google Scholar] [CrossRef]
- Mortus, J.R.; Zhang, Y.; Hughes, D.P.M. Developmental Pathways Hijacked by Osteosarcoma. Adv. Exp. Med. Biol. 2014, 804, 93–118. [Google Scholar] [CrossRef]
- Lindsey, B.A.; Markel, J.E.; Kleinerman, E.S. Osteosarcoma Overview. Rheumatol. Ther. 2017, 4, 25–43. [Google Scholar] [CrossRef]
- Thomas, D.M.; Johnson, S.A.; Sims, N.A.; Trivett, M.K.; Slavin, J.L.; Rubin, B.P.; Waring, P.; McArthur, G.A.; Walkley, C.R.; Holloway, A.J. Terminal osteoblast differentiation, mediated by runx2 and p27KIP1, is disrupted in osteosarcoma. J. Cell Biol. 2004, 167, 925–934. [Google Scholar] [CrossRef]
- Martin, J.W.; Zielenska, M.; Stein, G.S.; van Wijnen, A.J.; Squire, J.A. The Role of RUNX2 in Osteosarcoma Oncogenesis. Sarcoma 2011, 2011, 282745. [Google Scholar] [CrossRef]
- Cole, S.; Gianferante, D.M.; Zhu, B.; Mirabello, L. Osteosarcoma: A Surveillance, Epidemiology, and End Results program-based analysis from 1975 to 2017. Cancer 2022, 128, 2107–2118. [Google Scholar] [CrossRef]
- Mirabello, L.; Zhu, B.; Koster, R.; Karlins, E.; Dean, M.; Yeager, M.; Gianferante, M.; Spector, L.G.; Morton, L.M.; Karyadi, D.; et al. Frequency of Pathogenic Germline Variants in Cancer-Susceptibility Genes in Patients With Osteosarcoma. JAMA Oncol. 2020, 6, 724–734. [Google Scholar] [CrossRef]
- Kast, R.E.; Alfieri, A.; Assi, H.I.; Burns, T.C.; Elyamany, A.M.; Gonzalez-Cao, M.; Karpel-Massler, G.; Marosi, C.; Salacz, M.E.; Sardi, I.; et al. MDACT: A New Principle of Adjunctive Cancer Treatment Using Combinations of Multiple Repurposed Drugs, with an Example Regimen. Cancers 2022, 14, 2563. [Google Scholar] [CrossRef]
- Halatsch, M.-E.; Kast, R.E.; Karpel-Massler, G.; Mayer, B.; Zolk, O.; Schmitz, B.; Scheuerle, A.; Maier, L.; Bullinger, L.; Mayer-Steinacker, R.; et al. A phase Ib/IIa trial of 9 repurposed drugs combined with temozolomide for the treatment of recurrent glioblastoma: CUSP9v3. Neuro-Oncol. Adv. 2021, 3, vdab075. [Google Scholar] [CrossRef]
- Kast, R.E.; Karpel-Massler, G.; Halatsch, M.-E. CUSP9* treatment protocol for recurrent glioblastoma: Aprepitant, artesunate, auranofin, captopril, celecoxib, disulfiram, itraconazole, ritonavir, sertraline augmenting continuous low dose temozolomide. Oncotarget 2014, 5, 8052–8082. [Google Scholar] [CrossRef]
- Kast, R.E.; Boockvar, J.A.; Brüning, A.; Cappello, F.; Chang, W.-W.; Cvek, B.; Dou, Q.P.; Duenas-Gonzalez, A.; Efferth, T.; Focosi, D.; et al. A conceptually new treatment approach for relapsed glioblastoma: Coordinated undermining of survival paths with nine repurposed drugs (CUSP9) by the International Initiative for Accelerated Improvement of Glioblastoma Care. Oncotarget 2013, 4, 502–530. [Google Scholar] [CrossRef]
- Asija, S.; Chatterjee, A.; Yadav, S.; Chekuri, G.; Karulkar, A.; Jaiswal, A.K.; Goda, J.S.; Purwar, R. Combinatorial approaches to effective therapy in glioblastoma (GBM): Current status and what the future holds. Int. Rev. Immunol. 2022, 41, 582–605. [Google Scholar] [CrossRef]
- Dahlberg, D.; Rummel, J.; Distante, S.; De Souza, G.A.; Stensland, M.E.; Mariussen, E.; Rootwelt, H.; Voie, Ø.; Hassel, B. Glioblastoma microenvironment contains multiple hormonal and non-hormonal growth-stimulating factors. Fluids Barriers CNS 2022, 19, 45. [Google Scholar] [CrossRef]
- Kilmister, E.J.; Koh, S.P.; Weth, F.R.; Gray, C.; Tan, S.T. Cancer Metastasis and Treatment Resistance: Mechanistic Insights and Therapeutic Targeting of Cancer Stem Cells and the Tumor Microenvironment. Biomedicines 2022, 10, 2988. [Google Scholar] [CrossRef]
- Martell, E.; Kuzmychova, H.; Senthil, H.; Kaul, E.; Chokshi, C.R.; Venugopal, C.; Anderson, C.M.; Singh, S.K.; Sharif, T. Compensatory cross-talk between autophagy and glycolysis regulates senescence and stemness in heterogeneous glioblastoma tumor subpopulations. Acta Neuropathol. Commun. 2023, 11, 110. [Google Scholar] [CrossRef]
- Nguyen, H.S.; Shabani, S.; Awad, A.J.; Kaushal, M.; Doan, N. Molecular Markers of Therapy-Resistant Glioblastoma and Potential Strategy to Combat Resistance. Int. J. Mol. Sci. 2018, 19, 1765. [Google Scholar] [CrossRef]
- Palmer, A.C.; Chidley, C.; Sorger, P.K. A curative combination cancer therapy achieves high fractional cell killing through low cross-resistance and drug additivity. eLife 2019, 8, e50036. [Google Scholar] [CrossRef]
- Shafi, O.; Siddiqui, G. Tracing the origins of glioblastoma by investigating the role of gliogenic and related neurogenic genes/signaling pathways in GBM development: A systematic review. World J. Surg. Oncol. 2022, 20, 146. [Google Scholar] [CrossRef]
- Westermarck, J. Inhibition of adaptive therapy tolerance in cancer: Is triplet mitochondrial targeting the key? Mol. Oncol. 2023, 17, 537–540. [Google Scholar] [CrossRef]
- Pongratz, P.; Kurth, F.; Ngoma, G.M.; Basra, A.; Ramharter, M. In vitro activity of antifungal drugs against Plasmodium falciparum field isolates. Wien. Klin. Wochenschr. 2011, 123 (Suppl. S1), 26–30. [Google Scholar] [CrossRef]
- Aapro, M.; Carides, A.; Rapoport, B.L.; Schmoll, H.-J.; Zhang, L.; Warr, D. Aprepitant and Fosaprepitant: A 10-Year Review of Efficacy and Safety. Oncologist 2015, 20, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Padilla, A.; Habib, A.S. A pharmacological overview of aprepitant for the prevention of postoperative nausea and vomiting. Expert Rev. Clin. Pharmacol. 2023, 16, 491–505. [Google Scholar] [CrossRef] [PubMed]
- Kast, R.E.; Ramiro, S.; Lladó, S.; Toro, S.; Coveñas, R.; Muñoz, M. Antitumor action of temozolomide, ritonavir and aprepitant against human glioma cells. J. Neuro-Oncol. 2016, 126, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Kast, R.E. Glioblastoma: Synergy of growth promotion between CCL5 and NK-1R can be thwarted by blocking CCL5 with miraviroc, an FDA approved anti-HIV drug and blocking NK-1R with aprepitant, an FDA approved anti-nausea drug. J. Clin. Pharm. Ther. 2010, 35, 657–663. [Google Scholar] [CrossRef] [PubMed]
- Kast, R.E. Why cerebellar glioblastoma is rare and how that indicates adjunctive use of the FDA-approved anti-emetic aprepitant might retard cerebral glioblastoma growth: A new hypothesis to an old question. Clin. Transl. Oncol. 2009, 11, 408–410. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, M.; Coveñas, R. The Neurokinin-1 Receptor Antagonist Aprepitant: An Intelligent Bullet against Cancer? Cancers 2020, 12, 2682. [Google Scholar] [CrossRef]
- Muñoz, M.; Parrilla, J.; Rosso, M.; Coveñas, R. Antipruritic vs. Antitumour Action of Aprepitant: A Question of Dose. Acta Derm. Venereol. 2019, 99, 620–621. [Google Scholar] [CrossRef]
- Beirith, I.; Renz, B.W.; Mudusetti, S.; Ring, N.S.; Kolorz, J.; Koch, D.; Bazhin, A.V.; Berger, M.; Wang, J.; Angele, M.K.; et al. Identification of the Neurokinin-1 Receptor as Targetable Stratification Factor for Drug Repurposing in Pancreatic Cancer. Cancers 2021, 13, 2703. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, L.; Dong, D.; Wang, Z.; Ji, W.; Yu, M.; Zhang, F.; Niu, R.; Zhou, Y. MiR-34b/c-5p and the neurokinin-1 receptor regulate breast cancer cell proliferation and apoptosis. Cell Prolif. 2019, 52, e12527. [Google Scholar] [CrossRef]
- Garnier, A.; Ilmer, M.; Becker, K.; Häberle, B.; VON Schweinitz, D.; Kappler, R.; Berger, M. Truncated neurokinin-1 receptor is an ubiquitous antitumor target in hepatoblastoma, and its expression is independent of tumor biology and stage. Oncol. Lett. 2016, 11, 870–878. [Google Scholar] [CrossRef][Green Version]
- Ebrahimi, S.; Erfani, B.; Alalikhan, A.; Ghorbani, H.; Farzadnia, M.; Afshari, A.R.; Mashkani, B.; Hashemy, S.I. The In Vitro Pro-inflammatory Functions of the SP/NK1R System in Prostate Cancer: A Focus on Nuclear Factor-Kappa B (NF-κB) and Its Pro-inflammatory Target Genes. Appl. Biochem. Biotechnol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Korfi, F.; Javid, H.; Darban, R.A.; Hashemy, S.I. The Effect of SP/NK1R on the Expression and Activity of Catalase and Superoxide Dismutase in Glioblastoma Cancer Cells. Biochem. Res. Int. 2021, 2021, 6620708. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.-W.; Li, L.; Hu, W.-Q.; Hu, M.-N.; Tao, Y.; Hu, H.; Miao, X.-K.; Yang, W.-L.; Zhu, Q.; Mou, L.-Y. Neurokinin-1 receptor promotes non-small cell lung cancer progression through transactivation of EGFR. Cell Death Dis. 2022, 13, 6620708. [Google Scholar] [CrossRef] [PubMed]
- Javid, H.; Asadi, J.; Avval, F.Z.; Afshari, A.R.; Hashemy, S.I. The role of substance P/neurokinin 1 receptor in the pathogenesis of esophageal squamous cell carcinoma through constitutively active PI3K/Akt/NF-κB signal transduction pathways. Mol. Biol. Rep. 2020, 47, 2253–2263. [Google Scholar] [CrossRef] [PubMed]
- Ilmer, M.; Garnier, A.; Vykoukal, J.; Alt, E.; von Schweinitz, D.; Kappler, R.; Berger, M. Targeting the Neurokinin-1 Receptor Compromises Canonical Wnt Signaling in Hepatoblastoma. Mol. Cancer Ther. 2015, 14, 2712–2721. [Google Scholar] [CrossRef]
- Golestaneh, M.; Firoozrai, M.; Javid, H.; Hashemy, S.I. The substance P/neurokinin-1 receptor signaling pathway mediates metastasis in human colorectal SW480 cancer cells. Mol. Biol. Rep. 2022, 49, 4893–4900. [Google Scholar] [CrossRef]
- Henssen, A.G.; Odersky, A.; Szymansky, A.; Seiler, M.; Althoff, K.; Beckers, A.; Speleman, F.; Schäfers, S.; De Preter, K.; Astrahanseff, K.; et al. Targeting tachykinin receptors in neuroblastoma. Oncotarget 2017, 8, 430–443. [Google Scholar] [CrossRef]
- Kolorz, J.; Demir, S.; Gottschlich, A.; Beirith, I.; Ilmer, M.; Lüthy, D.; Walz, C.; Dorostkar, M.M.; Magg, T.; Hauck, F.; et al. The Neurokinin-1 Receptor Is a Target in Pediatric Rhabdoid Tumors. Curr. Oncol. 2021, 29, 94–110. [Google Scholar] [CrossRef]
- Mohammadi, F.; Javid, H.; Afshari, A.R.; Mashkani, B.; Hashemy, S.I. Substance P accelerates the progression of human esophageal squamous cell carcinoma via MMP-2, MMP-9, VEGF-A, and VEGFR1 overexpression. Mol. Biol. Rep. 2020, 47, 4263–4272. [Google Scholar] [CrossRef]
- Berger, M.; Neth, O.; Ilmer, M.; Garnier, A.; Salinas-Martín, M.V.; de Agustín Asencio, J.C.; von Schweinitz, D.; Kappler, R.; Muñoz, M. Hepatoblastoma cells express truncated neurokinin-1 receptor and can be growth inhibited by aprepitant in vitro and in vivo. J. Hepatol. 2014, 60, 985–994. [Google Scholar] [CrossRef]
- Akbari, S.; Darban, R.A.; Javid, H.; Esparham, A.; Hashemy, S.I. The anti-tumoral role of Hesperidin and Aprepitant on prostate cancer cells through redox modifications. Naunyn-Schmiedeberg’s Arch. Pharmacol. 2023, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dikmen, M.; Gökhaner, G.; Cantürk, Z. Evaluation of the antileukemic effects of neurokinin-1 receptor antagonists, aprepitant, and L-733,060, in chronic and acute myeloid leukemic cells. Anti-Cancer Drugs 2019, 30, 693–705. [Google Scholar] [CrossRef]
- Muñoz, M.; Rosso, M.; Robles-Frias, M.J.; Salinas-Martín, M.V.; Rosso, R.; González-Ortega, A.; Coveñas, R. The NK-1 receptor is expressed in human melanoma and is involved in the antitumor action of the NK-1 receptor antagonist aprepitant on melanoma cell lines. Lab. Investig. 2010, 90, 1259–1269. [Google Scholar] [CrossRef] [PubMed]
- Kast, R.E. High Neutrophil-to-Lymphocyte Ratio Facilitates Cancer Growth—Currently Marketed Drugs Tadalafil, Isotretinoin, Colchicine, and Omega-3 to Reduce It: The TICO Regimen. Cancers 2022, 14, 4965. [Google Scholar] [CrossRef]
- Bayati, S.; Bashash, D.; Ahmadian, S.; Safaroghli-Azar, A.; Alimoghaddam, K.; Ghavamzadeh, A.; Ghaffari, S.H. Inhibition of tachykinin NK1 receptor using aprepitant induces apoptotic cell death and G1 arrest through Akt/p53 axis in pre-B acute lymphoblastic leukemia cells. Eur. J. Pharmacol. 2016, 791, 274–283. [Google Scholar] [CrossRef] [PubMed]
- Molinos-Quintana, A.; Trujillo-Hacha, P.; Piruat, J.I.; Bejarano-García, J.A.; García-Guerrero, E.; Pérez-Simón, J.A.; Muñoz, M. Human acute myeloid leukemia cells express Neurokinin-1 receptor, which is involved in the antileukemic effect of Neurokinin-1 receptor antagonists. Investig. New Drugs 2019, 37, 17–26. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimi, S.; Mirzavi, F.; Hashemy, S.I.; Ghadiri, M.K.; Stummer, W.; Gorji, A. The in vitro anti-cancer synergy of neurokinin-1 receptor antagonist, aprepitant, and 5-aminolevulinic acid in glioblastoma. Biofactors 2023, 49, 900–911. [Google Scholar] [CrossRef]
- Garnier, A.; Vykoukal, J.; Hubertus, J.; Alt, E.; VON Schweinitz, D.; Kappler, R.; Berger, M.; Ilmer, M. Targeting the neurokinin-1 receptor inhibits growth of human colon cancer cells. Int. J. Oncol. 2015, 47, 151–160. [Google Scholar] [CrossRef]
- Halik, P.K.; Lipiński, P.F.J.; Matalińska, J.; Koźmiński, P.; Misicka, A.; Gniazdowska, E. Radiochemical Synthesis and Evaluation of Novel Radioconjugates of Neurokinin 1 Receptor Antagonist Aprepitant Dedicated for NK1R-Positive Tumors. Molecules 2020, 25, 3756. [Google Scholar] [CrossRef]
- Cao, X.; Yang, Y.; Zhou, W.; Wang, Y.; Wang, X.; Ge, X.; Wang, F.; Zhou, F.; Deng, X.; Miao, L. Aprepitant inhibits the development and metastasis of gallbladder cancer via ROS and MAPK activation. BMC Cancer 2023, 23, 471. [Google Scholar] [CrossRef]
- Zheng, Y.; Sang, M.; Liu, F.; Gu, L.; Li, J.; Wu, Y.; Shan, B. Aprepitant inhibits the progression of esophageal squamous cancer by blocking the truncated neurokinin-1 receptor. Oncol. Rep. 2023, 50, 131. [Google Scholar] [CrossRef]
- Muñoz, M.; González-Ortega, A.; Coveñas, R. The NK-1 receptor is expressed in human leukemia and is involved in the antitumor action of aprepitant and other NK-1 receptor antagonists on acute lymphoblastic leukemia cell lines. Investig. New Drugs 2012, 30, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, M.; Berger, M.; Rosso, M.; Gonzalez-Ortega, A.; Carranza, A.; Coveñas, R. Antitumor activity of neurokinin-1 receptor antagonists in MG-63 human osteosarcoma xenografts. Int. J. Oncol. 2014, 44, 137–146. [Google Scholar] [CrossRef]
- Robinson, P.; Rosso, M.; Muñoz, M. Neurokinin-1 Receptor Antagonists as a Potential Novel Therapeutic Option for Osteosarcoma Patients. J. Clin. Med. 2023, 12, 2135. [Google Scholar] [CrossRef] [PubMed]
- Alsaeed, M.A.; Ebrahimi, S.; Alalikhan, A.; Hashemi, S.F.; Hashemy, S.I. The Potential In Vitro Inhibitory Effects of Neurokinin-1 Receptor (NK-1R) Antagonist, Aprepitant, in Osteosarcoma Cell Migration and Metastasis. BioMed Res. Int. 2022, 2022, 8082608. [Google Scholar] [CrossRef] [PubMed]
- Togari, A.; Arai, M.; Mizutani, S.; Mizutani, S.; Koshihara, Y.; Nagatsu, T. Expression of mRNAs for neuropeptide receptors and β-adrenergic receptors in human osteoblasts and human osteogenic sarcoma cells. Neurosci. Lett. 1997, 233, 125–128. [Google Scholar] [CrossRef]
- Krasselt, M.; Baerwald, C. Celecoxib for the treatment of musculoskeletal arthritis. Expert Opin. Pharmacother. 2019, 20, 1689–1702. [Google Scholar] [CrossRef]
- Tive, L. Celecoxib clinical profile. Rheumatology 2000, 39 (Suppl. S2), 21–28. [Google Scholar] [CrossRef]
- Teerawattananon, C.; Tantayakom, P.; Suwanawiboon, B.; Katchamart, W. Risk of perioperative bleeding related to highly selective cyclooxygenase-2 inhibitors: A systematic review and meta-analysis. Semin. Arthritis Rheum. 2017, 46, 520–528. [Google Scholar] [CrossRef]
- Ho, K.Y.; Gwee, K.A.; Cheng, Y.K.; Yoon, K.H.; Hee, H.T.; Omar, A.R. Nonsteroidal anti-inflammatory drugs in chronic pain: Implications of new data for clinical practice. J. Pain Res. 2018, 11, 1937–1948. [Google Scholar] [CrossRef]
- Walker, C. Are All Oral COX-2 Selective Inhibitors the Same? A Consideration of Celecoxib, Etoricoxib, and Diclofenac. Int. J. Rheumatol. 2018, 2018, 1302835. [Google Scholar] [CrossRef]
- Tołoczko-Iwaniuk, N.; Dziemiańczyk-Pakieła, D.; Nowaszewska, B.K.; Celińska-Janowicz, K.; Miltyk, W. Celecoxib in Cancer Therapy and Prevention—Review. Curr. Drug Targets 2019, 20, 302–315. [Google Scholar] [CrossRef] [PubMed]
- Saxena, P.; Sharma, P.K.; Purohit, P. A journey of celecoxib from pain to cancer. Prostaglandins Other Lipid Mediat. 2020, 147, 106379. [Google Scholar] [CrossRef] [PubMed]
- Kast, R.E. Adding high-dose celecoxib to increase effectiveness of standard glioblastoma chemoirradiation. Ann. Pharm. Fr. 2021, 79, 481–488. [Google Scholar] [CrossRef] [PubMed]
- North, G.L.T. Celecoxib as Adjunctive Therapy for Treatment of Colorectal Cancer. Ann. Pharmacother. 2001, 35, 1638–1643. [Google Scholar] [CrossRef]
- Li, J.; Hao, Q.; Cao, W.; Vadgama, J.V.; Wu, Y. Celecoxib in breast cancer prevention and therapy. Cancer Manag. Res. 2018, 10, 4653–4667. [Google Scholar] [CrossRef]
- Liu, R.; Xu, K.-P.; Tan, G.-S. Cyclooxygenase-2 inhibitors in lung cancer treatment: Bench to bed. Eur. J. Pharmacol. 2015, 769, 127–133. [Google Scholar] [CrossRef]
- Tudor, D.V.; Bâldea, I.; Lupu, M.; Kacso, T.; Kutasi, E.; Hopârtean, A.; Stretea, R.; Filip, A.G. COX-2 as a potential biomarker and therapeutic target in melanoma. Cancer Biol. Med. 2020, 17, 20–31. [Google Scholar] [CrossRef]
- Gore, E. Celecoxib and radiation therapy in non-small-cell lung cancer. Oncology 2004, 18 (Suppl. S14), 10–14. [Google Scholar]
- Futagami, S.; Suzuki, K.; Hiratsuka, T.; Shindo, T.; Hamamoto, T.; Ueki, N.; Kusunoki, M.; Miyake, K.; Gudis, K.; Tsukui, T.; et al. Chemopreventive effect of celecoxib in gastric cancer. Inflammopharmacology 2007, 15, 1–4. [Google Scholar] [CrossRef]
- Khan, Z.; Khan, N.; Tiwari, R.P.; Sah, N.K.; Prasad, G.B.; Bisen, P.S. Biology of Cox-2: An Application in Cancer Therapeutics. Curr. Drug Targets 2011, 12, 1082–1093. [Google Scholar] [CrossRef]
- Liu, G.; Yu, M.-Y.; Huang, X.; Zhu, D.; Cheng, S.; Ma, R.; Gu, G. Synergistic effect of celecoxib in tumor necrosis factor-related apoptosis-inducing ligand treatment in osteosarcoma cells. Mol. Med. Rep. 2014, 10, 2198–2202. [Google Scholar] [CrossRef]
- Liu, J.; Wu, J.; Zhou, L.; Pan, C.; Zhou, Y.; Du, W.; Chen, J.-M.; Zhu, X.; Shen, J.; Chen, S.; et al. ZD6474, a new treatment strategy for human osteosarcoma, and its potential synergistic effect with celecoxib. Oncotarget 2015, 6, 21341–21352. [Google Scholar] [CrossRef]
- Zhou, X.; Shi, X.; Ren, K.; Fan, G.-T.; Wu, S.J.; Zhao, J.-N. Celecoxib inhibits cell growth and modulates the expression of matrix metalloproteinases in human osteosarcoma MG-63 cell line. Eur. Rev. Med. Pharmacol. Sci. 2015, 19, 4087–4097. [Google Scholar]
- Liu, B.; Yan, S.; Qu, L.; Zhu, J. Celecoxib enhances anticancer effect of cisplatin and induces anoikis in osteosarcoma via PI3K/Akt pathway. Cancer Cell Int. 2017, 17, 1. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.; Wang, X.; Jeschke, U.; von Schönfeldt, V. COX-2-PGE2-EPs in gynecological cancers. Arch. Gynecol. Obstet. 2020, 301, 1365–1375. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.; White, A.C.; Borowsky, A.D. New insights into the functions of Cox-2 in skin and esophageal malignancies. Exp. Mol. Med. 2020, 52, 538–547. [Google Scholar] [CrossRef]
- Zmigrodzka, M.; Rzepecka, A.; Krzyzowska, M.; Witkowska-Pilaszewicz, O.; Cywinska, A.; Winnicka, A. The cyclooxygenase-2/prostaglandin E2 pathway and its role in the pathogenesis of human and dog hematological malignancies. J. Physiol. Pharmacol. 2018, 69, 653–661. [Google Scholar] [CrossRef]
- Sheng, J.; Sun, H.; Yu, F.-B.; Li, B.; Zhang, Y.; Zhu, Y.-T. The Role of Cyclooxygenase-2 in Colorectal Cancer. Int. J. Med. Sci. 2020, 17, 1095–1101. [Google Scholar] [CrossRef]
- Pu, D.; Yin, L.; Huang, L.; Qin, C.; Zhou, Y.; Wu, Q.; Li, Y.; Zhou, Q.; Li, L. Cyclooxygenase-2 Inhibitor: A Potential Combination Strategy With Immunotherapy in Cancer. Front. Oncol. 2021, 11, 637504. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Guo, W.; Ren, T.; Huang, Y.; Wang, S.; Liu, K.; Zheng, B.; Yang, K.; Zhang, H.; Liang, X. Tumor-associated macrophages promote lung metastasis and induce epithelial-mesenchymal transition in osteosarcoma by activating the COX-2/STAT3 axis. Cancer Lett. 2019, 440–441, 116–125. [Google Scholar] [CrossRef] [PubMed]
- Zhou, M.; Wang, A.; Yin, B.; Wu, D.; Han, S.; Zhang, W.; Liu, J.; Sun, K. SND1 promotes the proliferation of osteosarcoma cells by upregulating COX-2/PGE2 expression via activation of NF-κB. Oncol. Rep. 2019, 41, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Naruse, T.; Nishida, Y.; Hosono, K.; Ishiguro, N. Meloxicam inhibits osteosarcoma growth, invasiveness and metastasis by COX-2-dependent and independent routes. Carcinogenesis 2006, 27, 584–592. [Google Scholar] [CrossRef]
- Qian, M.; Yang, X.; Li, Z.; Jiang, C.; Song, D.; Yan, W.; Liu, T.; Wu, Z.; Kong, J.; Wei, H.; et al. P50-associated COX-2 extragenic RNA (PACER) overexpression promotes proliferation and metastasis of osteosarcoma cells by activating COX-2 gene. Tumor Biol. 2016, 37, 3879–3886. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Cai, M.; Ji, F.; Lou, L.-M. The impact of COX-2 on invasion of osteosarcoma cell and its mechanism of regulation. Cancer Cell Int. 2014, 14, 27. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; He, M.; Xiao, Z.; Wu, H.; Wu, Y. Quantitative Assessment of the Association of COX-2 (Cyclooxygenase-2) Immunoexpression with Prognosis in Human Osteosarcoma: A Meta-Analysis. PLoS ONE 2013, 8, e82907. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Qu, P.; Zhao, H.; Zhao, T.; Cao, N. COX-2 promotes epithelial-mesenchymal transition and migration in osteosarcoma MG-63 cells via PI3K/AKT/NF-κB signaling. Mol. Med. Rep. 2019, 20, 3811–3819. [Google Scholar] [CrossRef]
- Masi, L.; Recenti, R.; Silvestri, S.; Pinzani, P.; Pepi, M.; Paglierani, M.; Brandi, M.L.; Franchi, A. Expression of Cyclooxygenase-2 in Osteosarcoma of Bone. Appl. Immunohistochem. Mol. Morphol. 2007, 15, 70–76. [Google Scholar] [CrossRef]
- Lee, E.J.; Choi, E.M.; Kim, S.R.; Park, J.H.; Kim, H.; Ha, K.S.; Kim, Y.M.; Kim, S.S.; Choe, M.; Kim, J.-I.; et al. Cyclooxygenase-2 promotes cell proliferation, migration and invasion in U2OS human osteosarcoma cells. Exp. Mol. Med. 2007, 39, 469–476. [Google Scholar] [CrossRef]
- Reckamp, K.L.; Krysan, K.; Morrow, J.D.; Milne, G.L.; Newman, R.A.; Tucker, C.; Elashoff, R.M.; Dubinett, S.M.; Figlin, R.A. A Phase I Trial to Determine the Optimal Biological Dose of Celecoxib when Combined with Erlotinib in Advanced Non–Small Cell Lung Cancer. Clin. Cancer Res. 2006, 12 Pt 1, 3381–3388. [Google Scholar] [CrossRef]
- Knudsen, J.F.; Carlsson, U.; Hammarström, P.; Sokol, G.H.; Cantilena, L.R. The Cyclooxygenase-2 Inhibitor Celecoxib Is a Potent Inhibitor of Human Carbonic Anhydrase II. Inflammation 2004, 28, 285–290. [Google Scholar] [CrossRef] [PubMed]
- Alper, A.B., Jr.; Tomlin, H.; Sadhwani, U.; Whelton, A.; Puschett, J. Effects of the Selective Cyclooxygenase-2 Inhibitor Analgesic Celecoxib on Renal Carbonic Anhydrase Enzyme Activity: A Randomized, Controlled Trial. Am. J. Ther. 2006, 13, 229–235. [Google Scholar] [CrossRef] [PubMed]
- Weber, A.; Casini, A.; Heine, A.; Kuhn, D.; Supuran, C.T.; Scozzafava, A.; Klebe, G. Unexpected Nanomolar Inhibition of Carbonic Anhydrase by COX-2-Selective Celecoxib: New Pharmacological Opportunities Due to Related Binding Site Recognition. J. Med. Chem. 2004, 47, 550–557. [Google Scholar] [CrossRef] [PubMed]
- De Monte, C.; Carradori, S.; Gentili, A.; Mollica, A.; Trisciuoglio, D.; Supuran, C.T. Dual Cyclooxygenase and Carbonic Anhydrase Inhibition by Nonsteroidal Anti-Inflammatory Drugs for the Treatment of Cancer. Curr. Med. Chem. 2015, 22, 2812–2818. [Google Scholar] [CrossRef]
- Mahboubi-Rabbani, M.; Zarghi, A. Dual Human Carbonic Anhydrase/Cyclooxygenase-2 Inhibitors: A Promising Approach for Cancer Treatment. Anti-Cancer Agents Med. Chem. 2021, 21, 2163–2180. [Google Scholar] [CrossRef]
- Pastorek, J.; Pastorekova, S. Hypoxia-induced carbonic anhydrase IX as a target for cancer therapy: From biology to clinical use. Semin. Cancer Biol. 2015, 31, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Takacova, M.; Kajanova, I.; Kolarcikova, M.; Lapinova, J.; Zatovicova, M.; Pastorekova, S. Understanding metabolic alterations and heterogeneity in cancer progression through validated immunodetection of key molecular components: A case of carbonic anhydrase IX. Cancer Metastasis Rev. 2021, 40, 1035–1053. [Google Scholar] [CrossRef]
- Potter, C.; Harris, A.L. Hypoxia Inducible Carbonic Anhydrase IX, Marker of Tumour: Hypoxia, Survival Pathway and Therapy Target. Cell Cycle 2004, 3, 159–162. [Google Scholar] [CrossRef]
- Andreucci, E.; Peppicelli, S.; Carta, F.; Brisotto, G.; Biscontin, E.; Ruzzolini, J.; Bianchini, F.; Biagioni, A.; Supuran, C.T.; Calorini, L. Carbonic anhydrase IX inhibition affects viability of cancer cells adapted to extracellular acidosis. J. Mol. Med. 2017, 95, 1341–1353. [Google Scholar] [CrossRef]
- Ward, C.; Meehan, J.; E Gray, M.; Murray, A.F.; Argyle, D.J.; Kunkler, I.H.; Langdon, S.P. The impact of tumour pH on cancer progression: Strategies for clinical intervention. Explor. Target. Anti-Tumor Ther. 2020, 1, 71–100. [Google Scholar] [CrossRef]
- Avnet, S.; Lemma, S.; Cortini, M.; Di Pompo, G.; Perut, F.; Lipreri, M.V.; Roncuzzi, L.; Columbaro, M.; Errani, C.; Longhi, A.; et al. The Release of Inflammatory Mediators from Acid-Stimulated Mesenchymal Stromal Cells Favours Tumour Invasiveness and Metastasis in Osteosarcoma. Cancers 2021, 13, 5855. [Google Scholar] [CrossRef] [PubMed]
- Avnet, S.; Di Pompo, G.; Chano, T.; Errani, C.; Ibrahim-Hashim, A.; Gillies, R.J.; Donati, D.M.; Baldini, N. Cancer-associated mesenchymal stroma fosters the stemness of osteosarcoma cells in response to intratumoral acidosis via NF-κB activation. Int. J. Cancer 2017, 140, 1331–1345. [Google Scholar] [CrossRef] [PubMed]
- Di Pompo, G.; Cortini, M.; Baldini, N.; Avnet, S. Acid Microenvironment in Bone Sarcomas. Cancers 2021, 13, 3848. [Google Scholar] [CrossRef]
- Feng, Z.; Ou, Y.; Hao, L. The roles of glycolysis in osteosarcoma. Front. Pharmacol. 2022, 13, 950886. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Wu, J.; Zhang, X.; Cao, L.; Wu, Y.; Miao, X. Transcription factor ELK1 accelerates aerobic glycolysis to enhance osteosarcoma chemoresistance through miR-134/PTBP1 signaling cascade. Aging 2021, 13, 6804–6819. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, E.; Tsukahara, T.; Emori, M.; Murata, K.; Akamatsu, A.; Shibayama, Y.; Hamada, S.; Watanabe, Y.; Kaya, M.; Hirohashi, Y.; et al. Osteosarcoma-initiating cells show high aerobic glycolysis and attenuation of oxidative phosphorylation mediated by LIN28B. Cancer Sci. 2020, 111, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Griffiths, J.R. How and Why Are Cancers Acidic? Carbonic Anhydrase IX and the Homeostatic Control of Tumour Extracellular pH. Cancers 2020, 12, 1616. [Google Scholar] [CrossRef]
- Okuno, K.; Matsubara, T.; Nakamura, T.; Iino, T.; Kakimoto, T.; Asanuma, K.; Matsumine, A.; Sudo, A. Carbonic anhydrase IX enhances tumor cell proliferation and tumor progression in osteosarcoma. OncoTargets Ther. 2018, 11, 6879–6886. [Google Scholar] [CrossRef]
- Perut, F.; Carta, F.; Bonuccelli, G.; Grisendi, G.; Di Pompo, G.; Avnet, S.; Sbrana, F.V.; Hosogi, S.; Dominici, M.; Kusuzaki, K.; et al. Carbonic anhydrase IX inhibition is an effective strategy for osteosarcoma treatment. Expert Opin. Ther. Targets 2015, 19, 1593–1605. [Google Scholar] [CrossRef]
- Matsubara, T.; DiResta, G.R.; Kakunaga, S.; Li, D.; Healey, J.H. Additive Influence of Extracellular pH, Oxygen Tension, and Pressure on Invasiveness and Survival of Human Osteosarcoma Cells. Front. Oncol. 2013, 3, 199. [Google Scholar] [CrossRef]
- Zhang, D.; Cui, G.; Sun, C.; Lei, L.; Williamson, R.A.; Wang, Y.; Zhang, J.; Chen, P.; Wang, A.; Fan, R.; et al. Hypoxia promotes osteosarcoma cell proliferation and migration through enhancing platelet-derived growth factor-BB/platelet-derived growth factor receptor-β axis. Biochem. Biophys. Res. Commun. 2019, 512, 360–366. [Google Scholar] [CrossRef]
- Nazon, C.; Pierrevelcin, M.; Willaume, T.; Lhermitte, B.; Weingertner, N.; Di Marco, A.; Bund, L.; Vincent, F.; Bierry, G.; Gomez-Brouchet, A.; et al. Together Intra-Tumor Hypoxia and Macrophagic Immunity Are Driven Worst Outcome in Pediatric High-Grade Osteosarcomas. Cancers 2022, 14, 1482. [Google Scholar] [CrossRef]
- Anai, S.; Tanaka, M.; Shiverick, K.T.; Kim, W.; Takada, S.; Boehlein, S.; Goodison, S.; Mizokami, A.; Rosser, C.J. Increased Expression of Cyclooxygenase-2 Correlates With Resistance to Radiation in Human Prostate Adenocarcinoma Cells. J. Urol. 2007, 177, 1913–1917. [Google Scholar] [CrossRef] [PubMed]
- Ihnatko, R.; Kubes, M.; Takacova, M.; Sedlakova, O.; Sedlak, J.; Pastorek, J.; Kopacek, J.; Pastorekova, S. Extracellular acidosis elevates carbonic anhydrase IX in human glioblastoma cells via transcriptional modulation that does not depend on hypoxia. Int. J. Oncol. 2006, 29, 1025–1033. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rezuchova, I.; Bartosova, M.; Belvoncikova, P.; Takacova, M.; Zatovicova, M.; Jelenska, L.; Csaderova, L.; Meciarova, I.; Pohlodek, K. Carbonic Anhydrase IX in Tumor Tissue and Plasma of Breast Cancer Patients: Reliable Biomarker of Hypoxia and Prognosis. Int. J. Mol. Sci. 2023, 24, 4325. [Google Scholar] [CrossRef] [PubMed]
- Slater, A.F. Chloroquine: Mechanism of drug action and resistance in Plasmodium falciparum. Pharmacol. Ther. 1993, 57, 203–235. [Google Scholar] [CrossRef]
- Nirk, E.L.; Reggiori, F.; Mauthe, M. Hydroxychloroquine in rheumatic autoimmune disorders and beyond. EMBO Mol. Med. 2020, 12, e12476. [Google Scholar] [CrossRef] [PubMed]
- Schrezenmeier, E.; Dörner, T. Mechanisms of action of hydroxychloroquine and chloroquine: Implications for rheumatology. Nat. Rev. Rheumatol. 2020, 16, 155–166. [Google Scholar] [CrossRef]
- Ferreira, P.M.P.; de Sousa, R.W.R.; Ferreira, J.R.d.O.; Militão, G.C.G.; Bezerra, D.P. Chloroquine and hydroxychloroquine in antitumor therapies based on autophagy-related mechanisms. Pharmacol. Res. 2021, 168, 105582. [Google Scholar] [CrossRef]
- Allemailem, K.S.; Almatroudi, A.; Alrumaihi, F.; Almatroodi, S.A.; Alkurbi, M.O.; Basfar, G.T.; Rahmani, A.H.; Khan, A.A. Novel Approaches of Dysregulating Lysosome Functions in Cancer Cells by Specific Drugs and Its Nanoformulations: A Smart Approach of Modern Therapeutics. Int. J. Nanomed. 2021, 16, 5065–5098. [Google Scholar] [CrossRef]
- Niemann, B.; Puleo, A.; Stout, C.; Markel, J.; Boone, B.A. Biologic Functions of Hydroxychloroquine in Disease: From COVID-19 to Cancer. Pharmaceutics 2022, 14, 2551. [Google Scholar] [CrossRef] [PubMed]
- Hraběta, J.; Belhajová, M.; Šubrtová, H.; Rodrigo, M.A.M.; Heger, Z.; Eckschlager, T. Drug Sequestration in Lysosomes as One of the Mechanisms of Chemoresistance of Cancer Cells and the Possibilities of Its Inhibition. Int. J. Mol. Sci. 2020, 21, 4392. [Google Scholar] [CrossRef]
- Jogalekar, M.P.; Veerabathini, A.; Gangadaran, P. Recent developments in autophagy-targeted therapies in cancer. Exp. Biol. Med. 2021, 246, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Ning, B.; Liu, Y.; Huang, T.; Wei, Y. Autophagy and its role in osteosarcoma. Cancer Med. 2023, 12, 5676–5687. [Google Scholar] [CrossRef]
- Xu, R.; Ji, Z.; Xu, C.; Zhu, J. The clinical value of using chloroquine or hydroxychloroquine as autophagy inhibitors in the treatment of cancers: A systematic review and meta-analysis. Medicine 2018, 97, e12912. [Google Scholar] [CrossRef] [PubMed]
- Dalpke, A.; Zimmermann, S.; Heeg, K. Immunopharmacology of CpG DNA. Biol. Chem. 2002, 383, 1491–1500. [Google Scholar] [CrossRef]
- Torigoe, M.; Sakata, K.; Ishii, A.; Iwata, S.; Nakayamada, S.; Tanaka, Y. Hydroxychloroquine efficiently suppresses inflammatory responses of human class-switched memory B cells via Toll-like receptor 9 inhibition. Clin. Immunol. 2018, 195, 1–7. [Google Scholar] [CrossRef]
- Kužnik, A.; Benčina, M.; Švajger, U.; Jeras, M.; Rozman, B.; Jerala, R. Mechanism of Endosomal TLR Inhibition by Antimalarial Drugs and Imidazoquinolines. J. Immunol. 2011, 186, 4794–4804. [Google Scholar] [CrossRef]
- Zhu, X.; Pan, Y.; Li, Y.; Jiang, Y.; Shang, H.; Gowda, D.C.; Cui, L.; Cao, Y. Targeting Toll-like receptors by chloroquine protects mice from experimental cerebral malaria. Int. Immunopharmacol. 2012, 13, 392–397. [Google Scholar] [CrossRef]
- Han, J.; Li, X.; Luo, X.; He, J.; Huang, X.; Zhou, Q.; Han, Y.; Jie, H.; Zhuang, J.; Li, Y.; et al. The mechanisms of hydroxychloroquine in rheumatoid arthritis treatment: Inhibition of dendritic cell functions via Toll like receptor 9 signaling. Biomed. Pharmacother. 2020, 132, 110848. [Google Scholar] [CrossRef]
- Marastoni, S.; Madariaga, A.; Pesic, A.; Nair, S.N.; Li, Z.J.; Shalev, Z.; Ketela, T.; Colombo, I.; Mandilaras, V.; Cabanero, M.; et al. Repurposing Itraconazole and Hydroxychloroquine to Target Lysosomal Homeostasis in Epithelial Ovarian Cancer. Cancer Res. Commun. 2022, 2, 293–306. [Google Scholar] [CrossRef]
- Yasuda, H.; Leelahavanichkul, A.; Tsunoda, S.; Dear, J.W.; Takahashi, Y.; Ito, S.; Hu, X.; Zhou, H.; Doi, K.; Childs, R.; et al. Chloroquine and inhibition of Toll-like receptor 9 protect from sepsis-induced acute kidney injury. Am. J. Physiol. Renal Physiol. 2008, 294, F1050–F1058. [Google Scholar] [CrossRef]
- Zou, W.; Amcheslavsky, A.; Bar-Shavit, Z. CpG Oligodeoxynucleotides Modulate the Osteoclastogenic Activity of Osteoblasts via Toll-like Receptor 9. J. Biol. Chem. 2003, 278, 16732–16740. [Google Scholar] [CrossRef] [PubMed]
- Jing, Y.; Liang, H.; Zhang, Y.; Cleveland, J.; Yan, J.; Zhang, D. Up-regulation of Toll-like Receptor 9 in Osteosarcoma. Anticancer Res. 2015, 35, 5839–5843. [Google Scholar] [PubMed]
- Jing, Y.; Jia, M.; Zhuang, J.; Han, D.; Zhou, C.; Yan, J. TLR9 Exerts an Oncogenic Role in Promoting Osteosarcoma Progression Depending on the Regulation of NF-κB Signaling Pathway. Biol. Pharm. Bull. 2022, 45, 1733–1742. [Google Scholar] [CrossRef] [PubMed]
- Mohamed, F.E.Z.; Jalan, R.; Minogue, S.; Andreola, F.; Habtesion, A.; Hall, A.; Winstanley, A.; Damink, S.O.; Malagó, M.; Davies, N.; et al. Inhibition of TLR7 and TLR9 Reduces Human Cholangiocarcinoma Cell Proliferation and Tumor Development. Dig. Dis. Sci. 2022, 67, 1806–1821. [Google Scholar] [CrossRef]
- González-Reyes, S.; Marín, L.; González, L.; O González, L.; del Casar, J.M.; Lamelas, M.L.; González-Quintana, J.M.; Vizoso, F.J. Study of TLR3, TLR4 and TLR9 in breast carcinomas and their association with metastasis. BMC Cancer 2010, 10, 665. [Google Scholar] [CrossRef]
- Sheyhidin, I.; Nabi, G.; Hasim, A.; Zhang, R.-P.; Ainiwaer, J.; Ma, H.; Wang, H. Overexpression of TLR3, TLR4, TLR7 and TLR9 in esophageal squamous cell carcinoma. World J. Gastroenterol. 2011, 17, 3745–3751. [Google Scholar] [CrossRef]
- Diaz-Ruiz, A.; Nader-Kawachi, J.; Calderón-Estrella, F.; Bermudez, A.M.; Alvarez-Mejia, L.; Ríos, C. Dapsone, More than an Effective Neuro and Cytoprotective Drug. Curr. Neuropharmacol. 2022, 20, 194–210. [Google Scholar] [CrossRef]
- Ghaoui, N.; Hanna, E.; Abbas, O.; Kibbi, A.; Kurban, M. Update on the use of dapsone in dermatology. Int. J. Dermatol. 2020, 59, 787–795. [Google Scholar] [CrossRef]
- Wozel, G.; Blasum, C. Dapsone in dermatology and beyond. Arch. Dermatol. Res. 2014, 306, 103–124. [Google Scholar] [CrossRef] [PubMed]
- Matsushima, K.; Shichino, S.; Ueha, S. Thirty-five years since the discovery of chemotactic cytokines, interleukin-8 and MCAF: A historical overview. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2023, 99, 213–226. [Google Scholar] [CrossRef] [PubMed]
- Karpel-Massler, G.; Kast, R.E.; Siegelin, M.D.; Dwucet, A.; Schneider, E.; Westhoff, M.-A.; Wirtz, C.R.; Chen, X.Y.; Halatsch, M.-E.; Bolm, C. Anti-glioma Activity of Dapsone and Its Enhancement by Synthetic Chemical Modification. Neurochem. Res. 2017, 42, 3382–3389. [Google Scholar] [CrossRef] [PubMed]
- Calderón-Estrella, F.; Franco-Bourland, R.E.; Rios, C.; de Jesús-Nicolás, D.; Pineda, B.; Méndez-Armenta, M.; Mata-Bermúdez, A.; Diaz-Ruiz, A. Early treatment with dapsone after spinal cord injury in rats decreases the inflammatory response and promotes long-term functional recovery. Heliyon 2023, 9, e14687. [Google Scholar] [CrossRef] [PubMed]
- Geyfman, M.; Debabov, D.; Poloso, N.; Alvandi, N. Mechanistic insight into the activity of a sulfone compound dapsone on Propionibacterium (Newly Reclassified as Cutibacterium) Acnes-mediated cytokine production. Exp. Dermatol. 2019, 28, 190–197. [Google Scholar] [CrossRef]
- Kast, R.E. Research Supporting a Pilot Study of Metronomic Dapsone during Glioblastoma Chemoirradiation. Med. Sci. 2021, 9, 12. [Google Scholar] [CrossRef]
- A Booth, S.; E Moody, C.; Dahl, M.V.; Herron, M.J.; Nelson, R.D. Dapsone Suppresses Integrin-Mediated Neutrophil Adherence Function. J. Investig. Dermatol. 1992, 98, 135–140. [Google Scholar] [CrossRef]
- Schmidt, E.; Reimer, S.; Kruse, N.; Bröcker, E.B.; Zillikens, D. The IL-8 release from cultured human keratinocytes, mediated by antibodies to bullous pemphigoid autoantigen 180, is inhibited by dapsone. Clin. Exp. Immunol. 2001, 124, 157–162. [Google Scholar] [CrossRef]
- Gremese, E.; Tolusso, B.; Bruno, D.; Perniola, S.; Ferraccioli, G.; Alivernini, S. The forgotten key players in rheumatoid arthritis: IL-8 and IL-17—Unmet needs and therapeutic perspectives. Front. Med. 2023, 10, 956127. [Google Scholar] [CrossRef]
- Kast, R.E. Dapsone as treatment adjunct in ARDS. Exp. Lung Res. 2020, 46, 157–161. [Google Scholar] [CrossRef]
- Kanwar, B.A.; Khattak, A.; Balentine, J.; Lee, J.H.; Kast, R.E. Benefits of Using Dapsone in Patients Hospitalized with COVID-19. Vaccines 2022, 10, 195. [Google Scholar] [CrossRef] [PubMed]
- Du, L.; Han, X.-G.; Tu, B.; Wang, M.-Q.; Qiao, H.; Zhang, S.-H.; Fan, Q.-M.; Tang, T.-T. CXCR1/Akt signaling activation induced by mesenchymal stem cell-derived IL-8 promotes osteosarcoma cell anoikis resistance and pulmonary metastasis. Cell Death Dis. 2018, 9, 714. [Google Scholar] [CrossRef]
- Zhang, Y.; Liu, Z.; Yang, X.; Lu, W.; Chen, Y.; Lin, Y.; Wang, J.; Lin, S.; Yun, J.-P. H3K27 acetylation activated-COL6A1 promotes osteosarcoma lung metastasis by repressing STAT1 and activating pulmonary cancer-associated fibroblasts. Theranostics 2021, 11, 1473–1492. [Google Scholar] [CrossRef]
- Jiang, H.; Wang, X.; Miao, W.; Wang, B.; Qiu, Y. CXCL8 promotes the invasion of human osteosarcoma cells by regulation of PI3K/Akt signaling pathway. Apmis 2017, 125, 773–780. [Google Scholar] [CrossRef]
- Liu, T.; Ma, Q.; Zhang, Y.; Wang, X.; Xu, K.; Yan, K.; Dong, W.; Fan, Q.; Zhang, Y.; Qiu, X. Self-seeding circulating tumor cells promote the proliferation and metastasis of human osteosarcoma by upregulating interleukin-8. Cell Death Dis. 2019, 10, 575. [Google Scholar] [CrossRef] [PubMed]
- Rutkowski, P.; Kamińska, J.; Kowalska, M.; Ruka, W.; Steffen, J. Cytokine and cytokine receptor serum levels in adult bone sarcoma patients: Correlations with local tumor extent and prognosis. J. Surg. Oncol. 2003, 84, 151–159. [Google Scholar] [CrossRef]
- Xiao, H.; Chen, L.; Luo, G.; Son, H.; Prectoni, J.H.; Zheng, W. Effect of the cytokine levels in serum on osteosarcoma. Tumor Biol. 2014, 35, 1023–1028. [Google Scholar] [CrossRef]
- Reinecke, J.B.; Roberts, R.D. Targetable Intercellular Signaling Pathways Facilitate Lung Colonization in Osteosarcoma. Adv. Exp. Med. Biol. 2020, 1258, 111–123. [Google Scholar] [CrossRef]
- Gross, A.C.; Cam, H.; Phelps, D.A.; Saraf, A.J.; Bid, H.K.; Cam, M.; London, C.A.; Winget, S.A.; Arnold, M.A.; Brandolini, L.; et al. IL-6 and CXCL8 mediate osteosarcoma-lung interactions critical to metastasis. J. Clin. Investig. 2018, 3, e99791. [Google Scholar] [CrossRef]
- Allende, C.; Higgins, B.; Johns, J. Comparison of serum cytokine concentrations between healthy dogs and canine osteosarcoma patients at the time of diagnosis. Veter. Immunol. Immunopathol. 2020, 227, 110084. [Google Scholar] [CrossRef]
- Zha, Z.; Su, A.; Huo, S. Activation of GPER suppresses the malignancy of osteosarcoma cells via down regulation of IL-6 and IL-8. Arch. Biochem. Biophys. 2018, 660, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Han, X.-G.; Du, L.; Qiao, H.; Tu, B.; Wang, Y.-G.; Qin, A.; Dai, K.-R.; Fan, Q.-M.; Tang, T.-T. CXCR1 knockdown improves the sensitivity of osteosarcoma to cisplatin. Cancer Lett. 2015, 369, 405–415. [Google Scholar] [CrossRef] [PubMed]
- Kawano, M.; Tanaka, K.; Itonaga, I.; Iwasaki, T.; Tsumura, H. Interaction between human osteosarcoma and mesenchymal stem cells via an interleukin-8 signaling loop in the tumor microenvironment. Cell Commun. Signal. 2018, 16, 13. [Google Scholar] [CrossRef] [PubMed]
- Pietrovito, L.; Leo, A.; Gori, V.; Lulli, M.; Parri, M.; Becherucci, V.; Piccini, L.; Bambi, F.; Taddei, M.L.; Chiarugi, P. Bone marrow-derived mesenchymal stem cells promote invasiveness and transendothelial migration of osteosarcoma cells via a mesenchymal to amoeboid transition. Mol. Oncol. 2018, 12, 659–676. [Google Scholar] [CrossRef] [PubMed]
- Kast, R.E. Erlotinib augmentation with dapsone for rash mitigation and increased anti-cancer effectiveness. Springerplus 2015, 4, 638. [Google Scholar] [CrossRef] [PubMed]
- Toledo-Guzmán, M.E.; Hernández, M.I.; Gómez-Gallegos, A.; Ortiz-Sánchez, E. ALDH as a Stem Cell Marker in Solid Tumors. Curr. Stem Cell Res. Ther. 2019, 14, 375–388. [Google Scholar] [CrossRef]
- Kang, X.; Jadhav, S.; Annaji, M.; Huang, C.-H.; Amin, R.; Shen, J.; Ashby, C.R., Jr.; Tiwari, A.K.; Babu, R.J.; Chen, P. Advancing Cancer Therapy with Copper/Disulfiram Nanomedicines and Drug Delivery Systems. Pharmaceutics 2023, 15, 1567. [Google Scholar] [CrossRef]
- Chico, M.A.; Mesas, C.; Doello, K.; Quiñonero, F.; Perazzoli, G.; Ortiz, R.; Prados, J.; Melguizo, C. Cancer Stem Cells in Sarcomas: In Vitro Isolation and Role as Prognostic Markers: A Systematic Review. Cancers 2023, 15, 2449. [Google Scholar] [CrossRef]
- Mu, X.; Patel, S.; Mektepbayeva, D.; Mahjoub, A.; Huard, J.; Weiss, K. Retinal Targets ALDH Positive Cancer Stem Cell and Alters the Phenotype of Highly Metastatic Osteosarcoma Cells. Sarcoma 2015, 2015, 784954. [Google Scholar] [CrossRef]
- Brown, H.K.; Tellez-Gabriel, M.; Heymann, D. Cancer stem cells in osteosarcoma. Cancer Lett. 2017, 386, 189–195. [Google Scholar] [CrossRef]
- Yu, S.; Fourman, M.S.; Mahjoub, A.; Mandell, J.B.; Crasto, J.A.; Greco, N.G.; Weiss, K.R. Lung cells support osteosarcoma cell migration and survival. BMC Cancer 2017, 17, 78. [Google Scholar] [CrossRef] [PubMed]
- Avdonkina, N.; Danilova, A.; Misyurin, V.; Prosekina, E.; Girdyuk, D.; Emelyanova, N.; Nekhaeva, T.; Gafton, G.; Baldueva, I. Biological features of tissue and bone sarcomas investigated using an in vitro model of clonal selection. Pathol. Res. Pract. 2020, 217, 153214. [Google Scholar] [CrossRef] [PubMed]
- Mandell, J.B.; Douglas, N.; Ukani, V.; Beumer, J.H.; Guo, J.; Payne, J.; Newman, R.; Mancinelli, L.; Intini, G.; Anderson, C.J.; et al. ALDH1A1 Gene Expression and Cellular Copper Levels between Low and Highly Metastatic Osteosarcoma Provide a Case for Novel Repurposing with Disulfiram and Copper. Sarcoma 2022, 2022, 7157507. [Google Scholar] [CrossRef] [PubMed]
- Belayneh, R.; Weiss, K. The Role of ALDH in the Metastatic Potential of Osteosarcoma Cells and Potential ALDH Targets. Adv. Exp. Med. Biol. 2020, 1258, 157–166. [Google Scholar] [CrossRef]
- Chu, M.; An, X.; Fu, C.; Yu, H.; Zhang, D.; Li, Q.; Man, X.; Dai, X.; Li, Z. Disulfiram/Copper Induce Ferroptosis in Triple-Negative Breast Cancer Cell Line MDA-MB-231. Front. Biosci. 2023, 28, 186. [Google Scholar] [CrossRef]
- Zheng, P.; Wu, Y.; Wang, Y.; Hu, F. Disulfiram suppresses epithelial-mesenchymal transition (EMT), migration and invasion in cervical cancer through the HSP90A/NDRG1 pathway. Cell. Signal. 2023, 109, 110771. [Google Scholar] [CrossRef]
- Zhang, S.; Zong, Y.; Chen, L.; Li, Q.; Li, Z.; Meng, R. The immunomodulatory function and antitumor effect of disulfiram: Paving the way for novel cancer therapeutics. Discov. Oncol. 2023, 14, 103. [Google Scholar] [CrossRef]
- Huang, S.; Xie, P.; Huang, X.; Chen, Z.; Yang, J.; Wang, J.; Liu, C.; Li, H.; Zhou, B. Disulfiram combined with chemoimmunotherapy potentiates pancreatic cancer treatment efficacy through the activation of cGAS-STING signaling pathway via suppressing PARP1 expression. Am. J. Cancer Res. 2023, 13, 2055–2065. [Google Scholar]
- Gan, Y.; Liu, T.; Feng, W.; Wang, L.; Li, L.; Ning, Y. Drug repositioning of disulfiram induces endometrioid epithelial ovarian cancer cell death via the both apoptosis and cuproptosis pathways. Oncol. Res. 2023, 31, 333–343. [Google Scholar] [CrossRef]
- Bermudez, N.M.; Rodríguez-Tamez, G.; Perez, S.; Tosti, A. Onychomycosis: Old and New. J. Fungi 2023, 9, 559. [Google Scholar] [CrossRef]
- Lestner, J.; Hope, W.W. Itraconazole: An update on pharmacology and clinical use for treatment of invasive and allergic fungal infections. Expert Opin. Drug Metab. Toxicol. 2013, 9, 911–926. [Google Scholar] [CrossRef] [PubMed]
- McKinsey, D.S. Treatment and Prevention of Histoplasmosis in Adults Living with HIV. J. Fungi 2021, 7, 429. [Google Scholar] [CrossRef]
- Rahi, M.S.; Jindal, V.; Pednekar, P.; Parekh, J.; Gunasekaran, K.; Sharma, S.; Stender, M.; Jaiyesimi, I.A. Fungal infections in hematopoietic stem-cell transplant patients: A review of epidemiology, diagnosis, and management. Ther. Adv. Infect. Dis. 2021, 8, 20499361211039050. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lan, C.; Qin, S.; Qin, Z.; Zhang, Z.; Zhang, P.; Cao, W. Efficacy of anti-fungal agents for invasive fungal infection prophylaxis in liver transplant recipients: A network meta-analysis. Mycoses 2022, 65, 906–917. [Google Scholar] [CrossRef] [PubMed]
- Li, C.-L.; Fang, Z.-X.; Wu, Z.; Hou, Y.-Y.; Wu, H.-T.; Liu, J. Repurposed itraconazole for use in the treatment of malignancies as a promising therapeutic strategy. Biomed. Pharmacother. 2022, 154, 113616. [Google Scholar] [CrossRef]
- Wei, X.; Liu, W.; Wang, J.Q.; Tang, Z. “Hedgehog pathway”: A potential target of itraconazole in the treatment of cancer. J. Cancer Res. Clin. Oncol. 2020, 146, 297–304. [Google Scholar] [CrossRef]
- Lin, M.; Chuang, Y.; Wu, H.; Hsu, C.; Lin, N.; Huang, M.; Lou, P. Targeting tumor O-glycosylation modulates cancer–immune-cell crosstalk and enhances anti-PD-1 immunotherapy in head and neck cancer. Mol. Oncol. 2023. [Google Scholar] [CrossRef]
- Lam, A.; Hoang, J.D.; Singleton, A.; Han, X.; Bleier, B.S. Itraconazole and clarithromycin inhibit P-glycoprotein activity in primary human sinonasal epithelial cells. Int. Forum Allergy Rhinol. 2015, 5, 477–480. [Google Scholar] [CrossRef]
- Steinhilber, D.; Jaschonek, K.; Knospe, J.; Morof, O.; Roth, H.J. Effects of novel antifungal azole derivatives on the 5-lipoxygenase and cyclooxygenase pathway. Arzneimittelforschung 1990, 40, 1260–1263. [Google Scholar]
- Saitoh, Y.; Setoguchi, T.; Nagata, M.; Tsuru, A.; Nakamura, S.; Nagano, S.; Ishidou, Y.; Nagao-Kitamoto, H.; Yokouchi, M.; Maeda, S.; et al. Combination of Hedgehog inhibitors and standard anticancer agents synergistically prevent osteosarcoma growth. Int. J. Oncol. 2016, 48, 235–242. [Google Scholar] [CrossRef]
- Lézot, F.; Corre, I.; Morice, S.; Rédini, F.; Verrecchia, F. SHH Signaling Pathway Drives Pediatric Bone Sarcoma Progression. Cells 2020, 9, 536. [Google Scholar] [CrossRef] [PubMed]
- Lo, W.W.; Wunder, J.S.; Dickson, B.C.; Campbell, V.; McGovern, K.; Alman, B.A.; Andrulis, I.L. Involvement and targeted intervention of dysregulated Hedgehog signaling in osteosarcoma. Cancer 2014, 120, 537–547. [Google Scholar] [CrossRef] [PubMed]
- Lo, W.W.; Pinnaduwage, D.; Gokgoz, N.; Wunder, J.S.; Andrulis, I.L. Aberrant Hedgehog Signaling and Clinical Outcome in Osteosarcoma. Sarcoma 2014, 2014, 261804. [Google Scholar] [CrossRef]
- Chan, L.H.; Wang, W.; Yeung, W.; Deng, Y.; Yuan, P.; Mak, K.K.-L. Hedgehog signaling induces osteosarcoma development through Yap1 and H19 overexpression. Oncogene 2014, 33, 4857–4866. [Google Scholar] [CrossRef]
- Yang, W.; Liu, X.; Choy, E.; Mankin, H.; Hornicek, F.J.; Duan, Z. Targeting hedgehog-GLI-2 pathway in osteosarcoma. J. Orthop. Res. 2013, 31, 502–509. [Google Scholar] [CrossRef] [PubMed]
- Yao, Z.; Han, L.; Chen, Y.; He, F.; Sun, B.; Kamar, S.; Zhang, Y.; Yang, Y.; Wang, C.; Yang, Z. Hedgehog signalling in the tumourigenesis and metastasis of osteosarcoma, and its potential value in the clinical therapy of osteosarcoma. Cell Death Dis. 2018, 9, 701. [Google Scholar] [CrossRef] [PubMed]
- Martins-Neves, S.R.; Sampaio-Ribeiro, G.; Gomes, C.M.F. Self-Renewal and Pluripotency in Osteosarcoma Stem Cells’ Chemoresistance: Notch, Hedgehog, and Wnt/β-Catenin Interplay with Embryonic Markers. Int. J. Mol. Sci. 2023, 24, 8401. [Google Scholar] [CrossRef] [PubMed]
- Cheng, D.; Li, J.; Li, S.; Yang, Q.; Fan, C. CNOT1 cooperates with LMNA to aggravate osteosarcoma tumorigenesis through the Hedgehog signaling pathway. Mol. Oncol. 2017, 11, 388–404. [Google Scholar] [CrossRef]
- Kumar, R.M.R.; Fuchs, B. Hedgehog Signaling Inhibitors as Anti-Cancer Agents in Osteosarcoma. Cancers 2015, 7, 784–794. [Google Scholar] [CrossRef]
- Wang, C.; Jing, J.; Hu, X.; Yu, S.; Yao, F.; Li, Z.; Cheng, L. Gankyrin activates the hedgehog signalling to drive metastasis in osteosarcoma. J. Cell. Mol. Med. 2021, 25, 6232–6241. [Google Scholar] [CrossRef]
- Qu, W.; Li, D.; Wang, Y.; Wu, Q.; Hao, D. Activation of Sonic Hedgehog Signaling Is Associated with Human Osteosarcoma Cells Radioresistance Characterized by Increased Proliferation, Migration, and Invasion. Med. Sci. Monit. 2018, 24, 3764–3771. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Kang, X.; Li, Z.; Chen, L.; Ma, Q.; Fan, P. Hedgehog/GLI1 signaling pathway regulates the resistance to cisplatin in human osteosarcoma. J. Cancer 2021, 12, 6676–6684. [Google Scholar] [CrossRef] [PubMed]
- Shen, P.-W.; Chou, Y.-M.; Li, C.-L.; Liao, E.-C.; Huang, H.-S.; Yin, C.-H.; Chen, C.-L.; Yu, S.-J. Itraconazole improves survival outcomes in patients with colon cancer by inducing autophagic cell death and inhibiting transketolase expression. Oncol. Lett. 2021, 22, 768. [Google Scholar] [CrossRef] [PubMed]
- Sawasaki, M.; Tsubamoto, H.; Sugihara, A.; Ikuta, S.; Sakai, Y.; Osaki, Y.; Sonoda, T. First-Line Gemcitabine, Nab-Paclitaxel, and Oxaliplatin Chemotherapy With Itraconazole in Patients With Metastatic Pancreatic Cancer: A Single Institution Experience. Anticancer Res. 2022, 42, 6063–6069. [Google Scholar] [CrossRef] [PubMed]
- Sawasaki, M.; Tsubamoto, H.; Nakamoto, Y.; Kakuno, A.; Sonoda, T. S-1, Oxaliplatin, Nab-paclitaxel and Itraconazole for Conversion Surgery for Advanced or Recurrent Gastric Cancer. Anticancer Res. 2020, 40, 991–997. [Google Scholar] [CrossRef]
- Rudin, C.M.; Brahmer, J.R.; Juergens, R.A.; Hann, C.L.; Ettinger, D.S.; Sebree, R.; Smith, R.; Aftab, B.T.; Huang, P.; Liu, J.O. Phase 2 Study of Pemetrexed and Itraconazole as Second-Line Therapy for Metastatic Nonsquamous Non–Small-Cell Lung Cancer. J. Thorac. Oncol. 2013, 8, 619–623. [Google Scholar] [CrossRef]
- Tsubamoto, H.; Sonoda, T.; Yamasaki, M.; Inoue, K. Impact of combination chemotherapy with itraconazole on survival of patients with refractory ovarian cancer. Anticancer Res. 2014, 34, 2481–2487. [Google Scholar]
- Vreugdenhil, G.; Raemaekers, J.M.M.; van Dijke, B.J.; de Pauw, B.E. Itraconazole and multidrug resistance: Possible effects on remission rate and disease-free survival in acute leukemia. Ann. Hematol. 1993, 67, 107–109. [Google Scholar] [CrossRef]
- Gupta, S.; Kim, J.; Gollapudi, S. Reversal of daunorubicin resistance in P388/ADR cells by itraconazole. J. Clin. Investig. 1991, 87, 1467–1469. [Google Scholar] [CrossRef]
- Warzecha, J.; Göttig, S.; Chow, K.; Brüning, C.; Percic, D.; Boehrer, S.; Brude, E.; Kurth, A. Inhibition of Osteosarcoma Cell Proliferation by the Hedgehog-Inhibitor Cyclopamine. J. Chemother. 2007, 19, 554–561. [Google Scholar] [CrossRef]
- Aftab, B.T.; Dobromilskaya, I.; Liu, J.O.; Rudin, C.M. Itraconazole Inhibits Angiogenesis and Tumor Growth in Non–Small Cell Lung Cancer. Cancer Res. 2011, 71, 6764–6772. [Google Scholar] [CrossRef] [PubMed]
- Nacev, B.A.; Grassi, P.; Dell, A.; Haslam, S.M.; Liu, J.O. The Antifungal Drug Itraconazole Inhibits Vascular Endothelial Growth Factor Receptor 2 (VEGFR2) Glycosylation, Trafficking, and Signaling in Endothelial Cells*. J. Biol. Chem. 2011, 286, 44045–44056. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Abe, Y.; Kawai, A.; Furihata, T.; Endo, T.; Takeda, H. Pharmacokinetic Drug Interactions of an Orally Available TRH Analog (Rovatirelin) with a CYP3A4/5 and P-Glycoprotein Inhibitor (Itraconazole). J. Clin. Pharmacol. 2020, 60, 1314–1323. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Unadkat, J.D.; Mao, Q. Interactions of azole antifungal agents with the human breast cancer resistance protein. J. Pharm. Sci. 2007, 96, 3226–3235. [Google Scholar] [CrossRef] [PubMed]
- Vermeer, L.M.; Isringhausen, C.D.; Ogilvie, B.W.; Buckley, D.B. Evaluation of Ketoconazole and Its Alternative Clinical CYP3A4/5 Inhibitors as Inhibitors of Drug Transporters: The In Vitro Effects of Ketoconazole, Ritonavir, Clarithromycin, and Itraconazole on 13 Clinically-Relevant Drug Transporters. Drug Metab. Dispos. 2016, 44, 453–459. [Google Scholar] [CrossRef] [PubMed]
- Lempers, V.J.C.; van den Heuvel, J.J.M.W.; Russel, F.G.M.; Aarnoutse, R.E.; Burger, D.M.; Brüggemann, R.J.; Koenderink, J.B. Inhibitory Potential of Antifungal Drugs on ATP-Binding Cassette Transporters P-Glycoprotein, MRP1 to MRP5, BCRP, and BSEP. Antimicrob. Agents Chemother. 2016, 60, 3372–3379. [Google Scholar] [CrossRef]
- Li, C.; Horton, J.K.; Sale, M.; Curd, L.; Goti, V.; Tao, W.; Beelen, A. Pharmacokinetic Drug–Drug Interaction Studies Between Trilaciclib and Midazolam, Metformin, Rifampin, Itraconazole, and Topotecan in Healthy Volunteers and Patients with Extensive-Stage Small-Cell Lung Cancer. Clin. Drug Investig. 2022, 42, 679–692. [Google Scholar] [CrossRef]
- Kurosawa, M.; Okabe, M.; Hara, N.; Kawamura, K.; Suzuki, S.; Sakurada, K.; Asaka, M. Reversal effect of itraconazole on adriamycin and etoposide resistance in human leukemia cells. Ann. Hematol. 1996, 72, 17–21. [Google Scholar] [CrossRef]
- Takimoto, Y.; Tsubamoto, H.; Taniguchi, R.; Sakata, K.; Takada, Y.; Adachi, J.; Tomonaga, T.; Ueda, T.; Nakagawa, K.; Narita, S.; et al. Itraconazole Repolarizes Tumor-associated Macrophages and Suppresses Cervical Cancer Cell Growth. Anticancer Res. 2023, 43, 569–580. [Google Scholar] [CrossRef]
- Weng, N.; Zhang, Z.; Tan, Y.; Zhang, X.; Wei, X.; Zhu, Q. Repurposing antifungal drugs for cancer therapy. J. Adv. Res. 2023, 48, 259–273. [Google Scholar] [CrossRef]
- Wu, H.-T.; Li, C.-L.; Fang, Z.-X.; Chen, W.-J.; Lin, W.-T.; Liu, J. Induced Cell Cycle Arrest in Triple-Negative Breast Cancer by Combined Treatment of Itraconazole and Rapamycin. Front. Pharmacol. 2022, 13, 873131. [Google Scholar] [CrossRef] [PubMed]
- Takimoto, Y.; Tsubamoto, H.; Isono-Taniguchi, R.; Ueda, T.; Sakata, K.; Nakagawa, K.; Narita, S.; Wakimoto, Y.; Shibahara, H.; Nishiumi, S. Itraconazole Modulates Phospholipid Levels in Tumor-associated Macrophages. Anticancer Res. 2023, 43, 1981–1984. [Google Scholar] [CrossRef]
- Remm, F.; Franz, W.-M.; Brenner, C. Gliptins and their target dipeptidyl peptidase 4: Implications for the treatment of vascular disease. Eur. Heart J. Cardiovasc. Pharmacother. 2016, 2, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Yurttas, A.G.; Dasci, M.F. Exploring the molecular mechanism of linagliptin in osteosarcoma cell lines for anti-cancer activity. Pathol. Res. Pract. 2023, 248, 154640. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Fu, B.; Luo, D.; Wang, H.; Cao, H.; Chen, X.; Tian, L.; Yu, X. The Multiple Biological Functions of Dipeptidyl Peptidase-4 in Bone Metabolism. Front. Endocrinol. 2022, 13, 856954. [Google Scholar] [CrossRef]
- Won, K.Y.; Park, H.-R.; Park, Y.-K. Prognostic Implication of Immunohistochemical Runx2 Expression in Osteosarcoma. Tumori J. 2009, 95, 311–316. [Google Scholar] [CrossRef]
- Kurek, K.C.; Del Mare, S.; Salah, Z.; Abdeen, S.; Sadiq, H.; Lee, S.-H.; Gaudio, E.; Zanesi, N.; Jones, K.B.; DeYoung, B.; et al. Frequent Attenuation of the WWOX Tumor Suppressor in Osteosarcoma Is Associated with Increased Tumorigenicity and Aberrant RUNX2 Expression. Cancer Res. 2010, 70, 5577–5586. [Google Scholar] [CrossRef]
- Sadikovic, B.; Thorner, P.; Chilton-MacNeill, S.; Martin, J.W.; Cervigne, N.K.; Squire, J.; Zielenska, M. Expression analysis of genes associated with human osteosarcoma tumors shows correlation of RUNX2 overexpression with poor response to chemotherapy. BMC Cancer 2010, 10, 202. [Google Scholar] [CrossRef]
- Zhang, H.; Lin, H.; Mo, X.; Chen, G.; Lin, L. Synergistic relationship between dipeptidyl peptidase IV and neutral endopeptidase expression and the combined prognostic significance in osteosarcoma patients. Med. Oncol. 2013, 30, 608. [Google Scholar] [CrossRef]
- Dohi, O.; Ohtani, H.; Hatori, M.; Sato, E.; Hosaka, M.; Nagura, H.; Itoi, E.; Kokubun, S. Histogenesis-specific expression of fibroblast activation protein and dipeptidylpeptidase-IV in human bone and soft tissue tumours. Histopathology 2009, 55, 432–440. [Google Scholar] [CrossRef]
- Hwang, H.-J.; Chung, H.S.; Jung, T.W.; Ryu, J.Y.; Hong, H.C.; Seo, J.A.; Kim, S.G.; Kim, N.H.; Choi, K.M.; Choi, D.S.; et al. The dipeptidyl peptidase-IV inhibitor inhibits the expression of vascular adhesion molecules and inflammatory cytokines in HUVECs via Akt- and AMPK-dependent mechanisms. Mol. Cell. Endocrinol. 2015, 405, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Nirwan, N.; Vohora, D. Linagliptin in Combination With Metformin Ameliorates Diabetic Osteoporosis Through Modulating BMP-2 and Sclerostin in the High-Fat Diet Fed C57BL/6 Mice. Front. Endocrinol. 2022, 13, 944323. [Google Scholar] [CrossRef] [PubMed]
- Liao, F.; Hu, X.; Chen, R. The effects of Omarigliptin on promoting osteoblastic differentiation. Bioengineered 2021, 12, 11837–11846. [Google Scholar] [CrossRef] [PubMed]
- Dong, C.; Yang, H.; Wang, Y.; Yan, X.; Li, D.; Cao, Z.; Ning, Y.; Zhang, C. Anagliptin stimulates osteoblastic cell differentiation and mineralization. Biomed. Pharmacother. 2020, 129, 109796. [Google Scholar] [CrossRef] [PubMed]
- Shao, H.; Wu, R.; Cao, L.; Gu, H.; Chai, F. Trelagliptin stimulates osteoblastic differentiation by increasing runt-related transcription factor 2 (RUNX2): A therapeutic implication in osteoporosis. Bioengineered 2021, 12, 960–968. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Qian, K.; Sun, Y.; Zhao, Y.; Zhou, Y.; Xue, Y.; Hong, X. Omarigliptin ameliorated high glucose-induced nucleotide oligomerization domain-like receptor protein 3 (NLRP3) inflammasome activation through activating adenosine monophosphate-activated protein kinase α (AMPKα) in renal glomerular endothelial cells. Bioengineered 2021, 12, 4805–4815. [Google Scholar] [CrossRef]
- Hu, Y.; Xu, J.; Wang, J.; Zhu, L.; Wang, J.; Zhang, Q. DPP-4 Inhibitors Suppress Tau Phosphorylation and Promote Neuron Autophagy through the AMPK/mTOR Pathway to Ameliorate Cognitive Dysfunction in Diabetic Mellitus. ACS Chem. Neurosci. 2023. [Google Scholar] [CrossRef]
- Al-Damry, N.T.; Attia, H.A.; Al-Rasheed, N.M.; Al-Rasheed, N.M.; Mohamad, R.A.; Al-Amin, M.A.; Dizmiri, N.; Atteya, M. Sitagliptin attenuates myocardial apoptosis via activating LKB-1/AMPK/Akt pathway and suppressing the activity of GSK-3β and p38α/MAPK in a rat model of diabetic cardiomyopathy. Biomed. Pharmacother. 2018, 107, 347–358. [Google Scholar] [CrossRef]
- Zeng, Y.; Li, C.; Guan, M.; Zheng, Z.; Li, J.; Xu, W.; Wang, L.; He, F.; Xue, Y. The DPP-4 inhibitor sitagliptin attenuates the progress of atherosclerosis in apolipoprotein-E-knockout mice via AMPK- and MAPK-dependent mechanisms. Cardiovasc. Diabetol. 2014, 13, 32. [Google Scholar] [CrossRef]
- Chen, Z.; Yu, J.; Fu, M.; Dong, R.; Yang, Y.; Luo, J.; Hu, S.; Li, W.; Xu, X.; Tu, L. Dipeptidyl peptidase-4 inhibition improves endothelial senescence by activating AMPK/SIRT1/Nrf2 signaling pathway. Biochem. Pharmacol. 2020, 177, 113951. [Google Scholar] [CrossRef]
- Ideta, T.; Shirakami, Y.; Miyazaki, T.; Kochi, T.; Sakai, H.; Moriwaki, H.; Shimizu, M. The Dipeptidyl Peptidase-4 Inhibitor Teneligliptin Attenuates Hepatic Lipogenesis via AMPK Activation in Non-Alcoholic Fatty Liver Disease Model Mice. Int. J. Mol. Sci. 2015, 16, 29207–29218. [Google Scholar] [CrossRef] [PubMed]
- Monami, M.; Dicembrini, I.; Antenore, A.; Mannucci, E. Dipeptidyl Peptidase-4 Inhibitors and Bone Fractures: A meta-analysis of randomized clinical trials. Diabetes Care 2011, 34, 2474–2476. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Xie, X.; Zhang, D.; Mao, F.; Wang, S.; Liao, Y. Saxagliptin enhances osteogenic differentiation in MC3T3-E1 cells, dependent on the activation of AMP-activated protein kinase α (AMPKα)/runt-related transcription factor-2 (Runx-2). Bioengineered 2022, 13, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Cordero, O.J. CD26 and Cancer. Cancers 2022, 14, 5194. [Google Scholar] [CrossRef]
- Cruickshank, J.M. The Role of Beta-Blockers in the Treatment of Hypertension. Adv. Exp. Med. Biol. 2016, 956, 149–166. [Google Scholar] [CrossRef]
- MacCormack, J.K.; Armstrong-Carter, E.L.B.; Gaudier-Diaz, M.M.; Meltzer-Brody, S.; Sloan, E.K.; Lindquist, K.A.; Muscatell, K.A. β-Adrenergic Contributions to Emotion and Physiology During an Acute Psychosocial Stressor. Psychosom. Med. 2021, 83, 959–968. [Google Scholar] [CrossRef]
- Brantigan, C.; Brantigan, T.; Joseph, N. Effect of beta blockade and beta stimulation on stage fright. Am. J. Med. 1982, 72, 88–94. [Google Scholar] [CrossRef]
- Qiao, G.; Bucsek, M.J.; Winder, N.M.; Chen, M.; Giridharan, T.; Olejniczak, S.H.; Hylander, B.L.; Repasky, E.A. β-Adrenergic signaling blocks murine CD8+ T-cell metabolic reprogramming during activation: A mechanism for immunosuppression by adrenergic stress. Cancer Immunol. Immunother. 2019, 68, 11–22. [Google Scholar] [CrossRef]
- Peixoto, R.; Pereira, M.d.L.; Oliveira, M. Beta-Blockers and Cancer: Where Are We? Pharmaceuticals 2020, 13, 105. [Google Scholar] [CrossRef]
- Vojvodic, A.; Vojvodic, P.; Vlaskovic-Jovicevic, T.; Sijan, G.; Dimitrijevic, S.; Peric-Hajzler, Z.; Matovic, D.; Wollina, U.; Tirant, M.; Thuong, N.V.; et al. Beta Blockers and Melanoma. Open Access Maced. J. Med. Sci. 2019, 7, 3110–3112. [Google Scholar] [CrossRef]
- Phadke, S.; Clamon, G. Beta blockade as adjunctive breast cancer therapy: A review. Crit. Rev. Oncol. Hematol. 2019, 138, 173–177. [Google Scholar] [CrossRef] [PubMed]
- Pantziarka, P.; Bouche, G.; Sukhatme, V.; Meheus, L.; Rooman, I.; Sukhatme, V.P. Repurposing Drugs in Oncology (ReDO)—Propranolol as an anti-cancer agent. Ecancermedicalscience 2016, 10, 680. [Google Scholar] [CrossRef] [PubMed]
- Wagner, M.J.; Cranmer, L.D.; Loggers, E.T.; Pollack, S.M. Propranolol for the treatment of vascular sarcomas. J. Exp. Pharmacol. 2018, 10, 51–58. [Google Scholar] [CrossRef]
- Cole, S.W.; Sood, A.K. Molecular Pathways: Beta-Adrenergic Signaling in Cancer. Clin. Cancer Res. 2012, 18, 1201–1206. [Google Scholar] [CrossRef]
- Yan, J.; Chen, Y.; Luo, M.; Hu, X.; Li, H.; Liu, Q.; Zou, Z. Chronic stress in solid tumor development: From mechanisms to interventions. J. Biomed. Sci. 2023, 30, 8. [Google Scholar] [CrossRef] [PubMed]
- Ammons, D.T.; MacDonald, C.R.; Chow, L.; Repasky, E.A.; Dow, S. Chronic adrenergic stress and generation of myeloid-derived suppressor cells: Implications for cancer immunotherapy in dogs. Veter. Comp. Oncol. 2023, 21, 159–165. [Google Scholar] [CrossRef]
- An, J.; Feng, L.; Ren, J.; Li, Y.; Li, G.; Liu, C.; Yao, Y.; Yao, Y.; Jiang, Z.; Gao, Y.; et al. Chronic stress promotes breast carcinoma metastasis by accumulating myeloid-derived suppressor cells through activating β-adrenergic signaling. OncoImmunology 2021, 10, 2004659. [Google Scholar] [CrossRef]
- Cao, M.; Huang, W.; Chen, Y.; Li, G.; Liu, N.; Wu, Y.; Wang, G.; Li, Q.; Kong, D.; Xue, T.; et al. Chronic restraint stress promotes the mobilization and recruitment of myeloid-derived suppressor cells through β-adrenergic-activated CXCL5-CXCR2-Erk signaling cascades. Int. J. Cancer 2021, 149, 460–472. [Google Scholar] [CrossRef]
- Liu, Y.; Tian, S.; Ning, B.; Huang, T.; Li, Y.; Wei, Y. Stress and cancer: The mechanisms of immune dysregulation and management. Front. Immunol. 2022, 13, 1032294. [Google Scholar] [CrossRef]
- Jiang, W.; Li, Y.; Li, Z.-Z.; Sun, J.; Li, J.-W.; Wei, W.; Li, L.; Zhang, C.; Huang, C.; Yang, S.-Y.; et al. Chronic restraint stress promotes hepatocellular carcinoma growth by mobilizing splenic myeloid cells through activating β-adrenergic signaling. Brain Behav. Immun. 2019, 80, 825–838. [Google Scholar] [CrossRef]
- Ieguchi, K.; Funakoshi, M.; Mishima, T.; Takizawa, K.; Omori, T.; Nakamura, F.; Watanabe, M.; Tsuji, M.; Kiuchi, Y.; Kobayashi, S.; et al. The Sympathetic Nervous System Contributes to the Establishment of Pre-Metastatic Pulmonary Microenvironments. Int. J. Mol. Sci. 2022, 23, 10652. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Liu, D.; Guo, L.; Cheng, X.; Guo, N.; Shi, M. Chronic psychological stress promotes lung metastatic colonization of circulating breast cancer cells by decorating a pre-metastatic niche through activating β-adrenergic signaling. J. Pathol. 2018, 244, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.-K.; Liu, J.-X.; Zheng, C.-X.; Liu, L.; Ma, C.; Tian, J.-Y.; Yuan, Y.; Cao, Y.; Xing, S.-J.; Liu, S.-Y.; et al. Region-specific sympatho-adrenergic regulation of specialized vasculature in bone homeostasis and regeneration. iScience 2023, 26, 107455. [Google Scholar] [CrossRef] [PubMed]
- Zhang, R.; Yin, H.; Yang, M.; Lei, X.; Zhen, D.; Zhang, Z. Advanced Progress of the Relationship Between Antihypertensive Drugs and Bone Metabolism. Hypertension 2023, 80, 2255–2264. [Google Scholar] [CrossRef]
- Bandala, C.; Ávila-Luna, A.; Gómez-López, M.; Estrada-Villaseñor, E.; Montes, S.; Alfaro-Rodríguez, A.; Miliar-García, Á.; Cortés-Altamirano, J.L.; Peralta, R.; Ibáñez-Cervantes, G.; et al. Catecholamine levels and gene expression of their receptors in tissues of adults with osteosarcoma. Arch. Physiol. Biochem. 2021, 127, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Karsenty, G.; Khosla, S. The crosstalk between bone remodeling and energy metabolism: A translational perspective. Cell Metab. 2022, 34, 805–817. [Google Scholar] [CrossRef]
- Conceição, F.; Sousa, D.M.; Tojal, S.; Lourenço, C.; Carvalho-Maia, C.; Estevão-Pereira, H.; Lobo, J.; Couto, M.; Rosenkilde, M.M.; Jerónimo, C.; et al. The Secretome of Parental and Bone Metastatic Breast Cancer Elicits Distinct Effects in Human Osteoclast Activity after Activation of β2 Adrenergic Signaling. Biomolecules 2023, 13, 622. [Google Scholar] [CrossRef]
- Guo, Q.; Chen, N.; Qian, C.; Qi, C.; Noller, K.; Wan, M.; Liu, X.; Zhang, W.; Cahan, P.; Cao, X. Sympathetic Innervation Regulates Osteocyte-Mediated Cortical Bone Resorption during Lactation. Adv. Sci. 2023, 10, e2207602. [Google Scholar] [CrossRef]
- Worton, L.E.; Srinivasan, S.; Threet, D.; Ausk, B.J.; Huber, P.; Kwon, R.Y.; Bain, S.D.; Gross, T.S.; Gardiner, E.M. Beta 2 Adrenergic Receptor Selective Antagonist Enhances Mechanically Stimulated Bone Anabolism in Aged Mice. JBMR Plus 2022, 7, e10712. [Google Scholar] [CrossRef]
- Koh, S.P.; Leadbitter, P.; Smithers, F.; Tan, S.T. β-blocker therapy for infantile hemangioma. Expert Rev. Clin. Pharmacol. 2020, 13, 899–915. [Google Scholar] [CrossRef]
- Solernó, L.M.; Sobol, N.T.; Gottardo, M.F.; Capobianco, C.S.; Ferrero, M.R.; Vásquez, L.; Alonso, D.F.; Garona, J. Propranolol blocks osteosarcoma cell cycle progression, inhibits angiogenesis and slows xenograft growth in combination with cisplatin-based chemotherapy. Sci. Rep. 2022, 12, 15058. [Google Scholar] [CrossRef] [PubMed]
- Duckett, M.M.; Phung, S.K.; Nguyen, L.; Khammanivong, A.; Dickerson, E.B.; Dusenbery, K.; Lawrence, J. The adrenergic receptor antagonists propranolol and carvedilol decrease bone sarcoma cell viability and sustained carvedilol reduces clonogenic survival and increases radiosensitivity in canine osteosarcoma cells. Veter. Comp. Oncol. 2020, 18, 128–140. [Google Scholar] [CrossRef] [PubMed]
- Majeska, R.J.; Minkowitz, B.; Bastian, W.; Einhorn, T.A. Effects of β-adrenergic blockade in an osteoblast-like cell line. J. Orthop. Res. 1992, 10, 379–384. [Google Scholar] [CrossRef] [PubMed]
- Golden, D.; Saria, E.A.; Hansen, M.F. Regulation of Osteoblast Migration Involving Receptor Activator of Nuclear Factor-kappa B (RANK) Signaling. J. Cell. Physiol. 2015, 230, 2951–2960. [Google Scholar] [CrossRef] [PubMed]
- González-Suárez, E.; Sanz-Moreno, A. RANK as a therapeutic target in cancer. FEBS J. 2016, 283, 2018–2033. [Google Scholar] [CrossRef]
- Kitazawa, S.; Kitazawa, R. RANK ligand is a prerequisite for cancer-associated osteolytic lesions. J. Pathol. 2002, 198, 228–236. [Google Scholar] [CrossRef]
- Roodman, G.D. Biology of Osteoclast Activation in Cancer. J. Clin. Oncol. 2001, 19, 3562–3571. [Google Scholar] [CrossRef]
- Branstetter, D.; Rohrbach, K.; Huang, L.-Y.; Soriano, R.; Tometsko, M.; Blake, M.; Jacob, A.P.; Dougall, W.C. RANK and RANK ligand expression in primary human osteosarcoma. J. Bone Oncol. 2015, 4, 59–68. [Google Scholar] [CrossRef]
- Bago-Horvath, Z.; Schmid, K.; Rössler, F.; Nagy-Bojarszky, K.; Funovics, P.; Sulzbacher, I. Impact of RANK signalling on survival and chemotherapy response in osteosarcoma. Pathology 2014, 46, 411–415. [Google Scholar] [CrossRef]
- Zeng, J.; Peng, Y.; Wang, D.; Ayesha, K.; Chen, S. The interaction between osteosarcoma and other cells in the bone microenvironment: From mechanism to clinical applications. Front. Cell Dev. Biol. 2023, 11, 1123065. [Google Scholar] [CrossRef]
- Zhu, J.; Liu, Y.; Zhu, Y.; Zeng, M.; Xie, J.; Lei, P.; Li, K.; Hu, Y. Role of RANK and Akt1 activation in human osteosarcoma progression: A clinicopathological study. Exp. Ther. Med. 2017, 13, 2862–2866. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Navet, B.; Ando, K.; Vargas-Franco, J.W.; Brion, R.; Amiaud, J.; Mori, K.; Yagita, H.; Mueller, C.G.; Verrecchia, F.; Dumars, C.; et al. The Intrinsic and Extrinsic Implications of RANKL/RANK Signaling in Osteosarcoma: From Tumor Initiation to Lung Metastases. Cancers 2018, 10, 398. [Google Scholar] [CrossRef] [PubMed]
- Trieb, K.; Windhager, R. Receptor activator of nuclear factor κB expression is a prognostic factor in human osteosarcoma. Oncol. Lett. 2015, 10, 1813–1815. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Shen, W.; Hu, X.; Zhang, Y.; Zhuo, Y.; Li, T.; Mei, F.; Li, X.; Xiao, L.; Chu, T. Quetiapine inhibits osteoclastogenesis and prevents human breast cancer-induced bone loss through suppression of the RANKL-mediated MAPK and NF-κB signaling pathways. Breast Cancer Res. Treat. 2015, 149, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Mi, G.; Wang, Y.; Ye, E.; Gao, Y.; Liu, Q.; Chen, P.; Zhu, Y.; Yang, H.; Yang, Z. The antipsychotic drug quetiapine stimulates oligodendrocyte differentiation by modulating the cell cycle. Neurochem. Int. 2018, 118, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Cardona, J.G.; Smith, M.D.; Wang, J.; Kirby, L.; Schott, J.T.; Davidson, T.; Karnell, J.L.; Whartenby, K.A.; Calabresi, P.A. Quetiapine has an additive effect to triiodothyronine in inducing differentiation of oligodendrocyte precursor cells through induction of cholesterol biosynthesis. PLoS ONE 2019, 14, e0221747. [Google Scholar] [CrossRef]
- Wang, Y.; Huang, N.; Li, H.; Liu, S.; Chen, X.; Yu, S.; Wu, N.; Bian, X.W.; Shen, H.Y.; Li, C.; et al. Promoting oligodendroglial-oriented differentiation of glioma stem cell: A repurposing of quetiapine for the treatment of malignant glioma. Oncotarget 2017, 8, 37511–37524. [Google Scholar] [CrossRef]
- Wang, X.; Su, Y.; Li, T.; Yu, G.; Wang, Y.; Chen, X.; Yin, C.; Tang, Z.; Yi, C.; Xiao, L.; et al. Quetiapine promotes oligodendroglial process outgrowth and membrane expansion by orchestrating the effects of Olig1. Glia 2021, 69, 1709–1722. [Google Scholar] [CrossRef]
- Bi, X.; Zhang, Y.; Yan, B.; Fang, S.; He, J.; Zhang, D.; Zhang, Z.; Kong, J.; Tan, Q.; Li, X. Quetiapine prevents oligodendrocyte and myelin loss and promotes maturation of oligodendrocyte progenitors in the hippocampus of global cerebral ischemia mice. J. Neurochem. 2012, 123, 14–20. [Google Scholar] [CrossRef]
- Ma, L.; Yang, F.; Zhao, R.; Li, L.; Kang, X.; Xiao, L.; Jiang, W. Quetiapine attenuates cognitive impairment and decreases seizure susceptibility possibly through promoting myelin development in a rat model of malformations of cortical development. Brain Res. 2015, 1622, 443–451. [Google Scholar] [CrossRef]
- Dandash, O.; Yücel, M.; Daglas, R.; Pantelis, C.; McGorry, P.; Berk, M.; Fornito, A. Differential effect of quetiapine and lithium on functional connectivity of the striatum in first episode mania. Transl. Psychiatry 2018, 8, 59. [Google Scholar] [CrossRef] [PubMed]
- Su, C.; Lin, S.; Wang, H.; Hsu, F.; Chung, J.G.; Hsu, L. The inhibitory effect and mechanism of quetiapine on tumor progression in hepatocellular carcinoma in vivo. Environ. Toxicol. 2022, 37, 92–100. [Google Scholar] [CrossRef] [PubMed]
- Cupp, M.A.; Cariolou, M.; Tzoulaki, I.; Aune, D.; Evangelou, E.; Berlanga-Taylor, A.J. Neutrophil to lymphocyte ratio and cancer prognosis: An umbrella review of systematic reviews and meta-analyses of observational studies. BMC Med. 2020, 18, 360. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Xiang, D.; Wan, J.; Yang, L.; Zheng, C. Focus on the Dynamics of Neutrophil-to-Lymphocyte Ratio in Cancer Patients Treated with Immune Checkpoint Inhibitors: A Meta-Analysis and Systematic Review. Cancers 2022, 14, 5297. [Google Scholar] [CrossRef]
- Wu, L.; Saxena, S.; Singh, R.K. Neutrophils in the Tumor Microenvironment. Adv. Exp. Med. Biol. 2020, 1224, 1–20. [Google Scholar] [CrossRef]
- Xia, W.-K.; Liu, Z.-L.; Shen, D.; Lin, Q.-F.; Su, J.; Mao, W.-D. Prognostic performance of pre-treatment NLR and PLR in patients suffering from osteosarcoma. World J. Surg. Oncol. 2016, 14, 127. [Google Scholar] [CrossRef]
- Yapar, A.; Tokgöz, M.A.; Yapar, D.; Atalay, I.B.; Ulucaköy, C.; Güngör, B. Diagnostic and prognostic role of neutrophil/lymphocyte ratio, platelet/lymphocyte ratio, and lymphocyte/monocyte ratio in patients with osteosarcoma. Jt. Dis. Relat. Surg. 2021, 32, 489–496. [Google Scholar] [CrossRef]
- Tang, H.; Liu, D.; Lu, J.; He, J.; Ji, S.; Liao, S.; Wei, Q.; Lu, S.; Liu, Y. Significance of the neutrophil-to-lymphocyte ratio in predicting the response to neoadjuvant chemotherapy in extremity osteosarcoma: A multicentre retrospective study. BMC Cancer 2022, 22, 33. [Google Scholar] [CrossRef]
- Peng, L.-P.; Li, J.; Li, X.-F. Prognostic value of neutrophil/lymphocyte, platelet/lymphocyte, lymphocyte/monocyte ratios and Glasgow prognostic score in osteosarcoma: A meta-analysis. World J. Clin. Cases 2022, 10, 2194–2205. [Google Scholar] [CrossRef]
- Vasquez, L.; León, E.; Beltran, B.; Maza, I.; Oscanoa, M.; Geronimo, J. Pretreatment Neutrophil-to-Lymphocyte Ratio and Lymphocyte Recovery: Independent Prognostic Factors for Survival in Pediatric Sarcomas. J. Pediatr. Hematol. Oncol. 2017, 39, 538–546. [Google Scholar] [CrossRef]
- Yang, S.; Wu, C.; Wang, L.; Shan, D.; Chen, B. Pretreatment inflammatory indexes as prognostic predictors for survival in osteosarcoma patients. Int. J. Clin. Exp. Pathol. 2020, 13, 515–524. [Google Scholar]
- Huang, X.; Liu, Y.; Liang, W.; Luo, K.; Qin, Y.; Li, F.; Xie, T.; Qin, H.; He, J.; Wei, Q. A new model of preoperative systemic inflammatory markers predicting overall survival of osteosarcoma: A multicenter retrospective study. BMC Cancer 2022, 22, 1370. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Wang, Y.; Ye, Q.; Wang, Y.; Min, L.; Luo, Y.; Zhou, Y.; Tu, C. Lung Immune Prognostic Index Could Predict Metastasis in Patients with Osteosarcoma. Front. Surg. 2022, 9, 923427. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Huang, Y.; Sun, Y.; Zhang, J.; Yao, Y.; Shen, Z.; Xiang, D.; He, A. Prognostic value of inflammation-based scores in patients with osteosarcoma. Sci. Rep. 2016, 6, 39862. [Google Scholar] [CrossRef] [PubMed]
- Peng, H.; Fang, X.; Xu, Y.; Wei, L.; Qin, Y.; Yang, F.; Lu, S.; Zhao, J. Prognostic value of preoperative P-CRP in patients with osteosarcoma: A retrospective study of 101 cases. Medicine 2022, 101, e30382. [Google Scholar] [CrossRef]
- Yang, Q.; Chen, T.; Yao, Z.; Zhang, X. Prognostic value of pre-treatment Naples prognostic score (NPS) in patients with osteosarcoma. World J. Surg. Oncol. 2020, 18, 24. [Google Scholar] [CrossRef]
- Ouyang, H.; Wang, Z. Predictive value of the systemic immune inflammation index for cancer-specific survival of osteosarcoma in children. Front. Public Health 2022, 10, 879523. [Google Scholar] [CrossRef]
- Tian, K.; Li, P.J.; Zhang, Y. Preoperative Predictors of Early Mortality Risk in People with Osteosarcoma of the Extremities Treated with Standard Therapy. Cancer Manag. Res. 2022, 14, 437–447. [Google Scholar] [CrossRef]
- Nirala, B.K.; Yamamichi, T.; Yustein, J.T. Deciphering the Signaling Mechanisms of Osteosarcoma Tumorigenesis. Int. J. Mol. Sci. 2023, 24, 11367. [Google Scholar] [CrossRef]
- Adamopoulos, C.; Gargalionis, A.N.; Basdra, E.K.; Papavassiliou, A.G. Deciphering signaling networks in osteosarcoma pathobiology. Exp. Biol. Med. 2016, 241, 1296–1305. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).