
Unlocking the Potential of Carbon Quantum Dots for Cell Imaging, Intracellular Localization, and Gene Expression Control in Arabidopsis thaliana (L.) Heynh.


 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Characterization of CQDs
2.2. Cell Localization of CQDs in Arabidopsis thaliana
2.3. Co-Localization Analyzation of AGO3/GFP Fusion Proteins and CQDs
2.4. The Moving Trail of CQDs in Arabidopsis thaliana
2.5. Transcriptome Analysis
2.6. Gene Expression Validation through RT-qPCR
2.7. Analysis of Genetic Properties of Targeted Genes
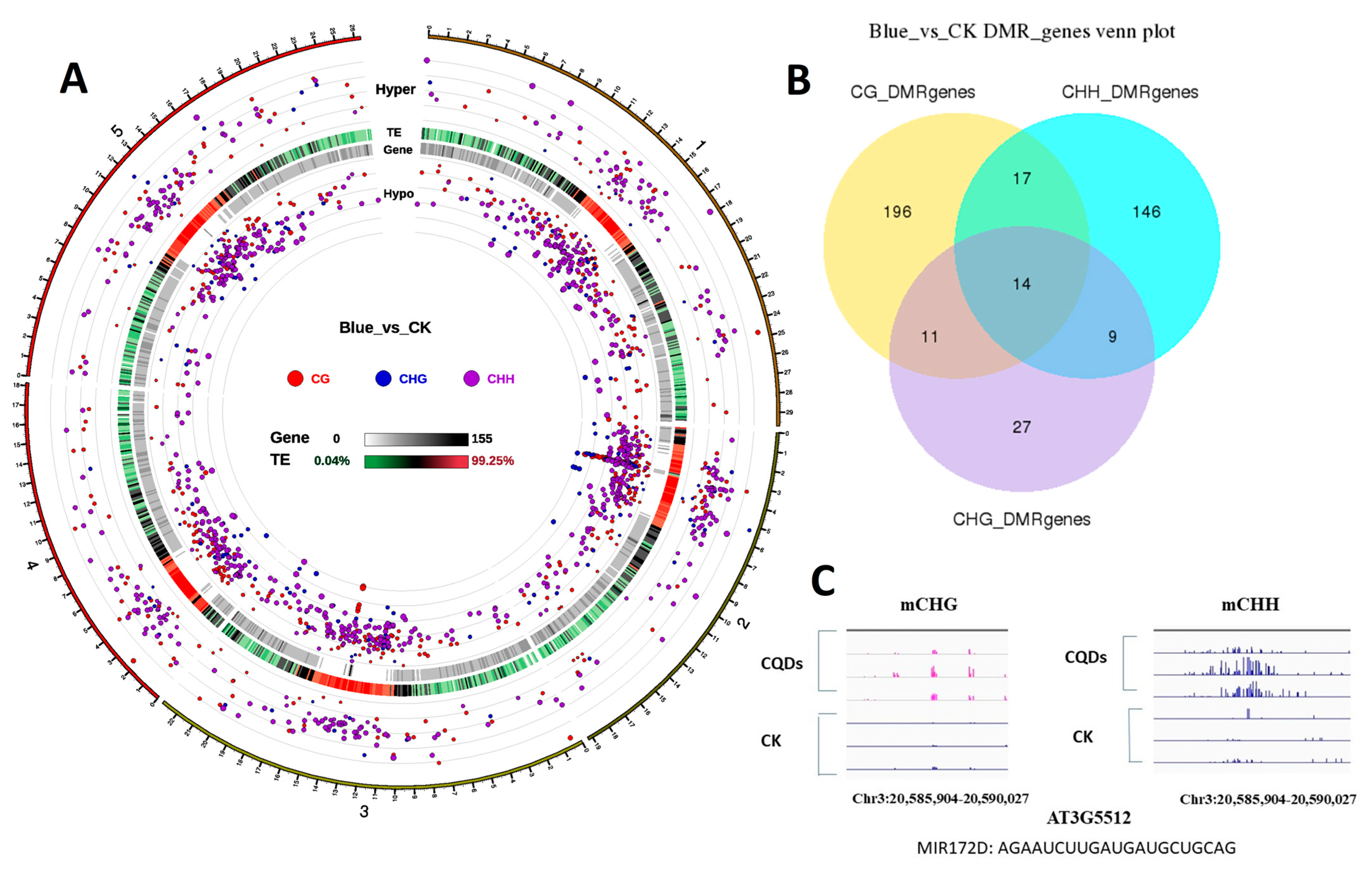
2.8. DNA Methylation
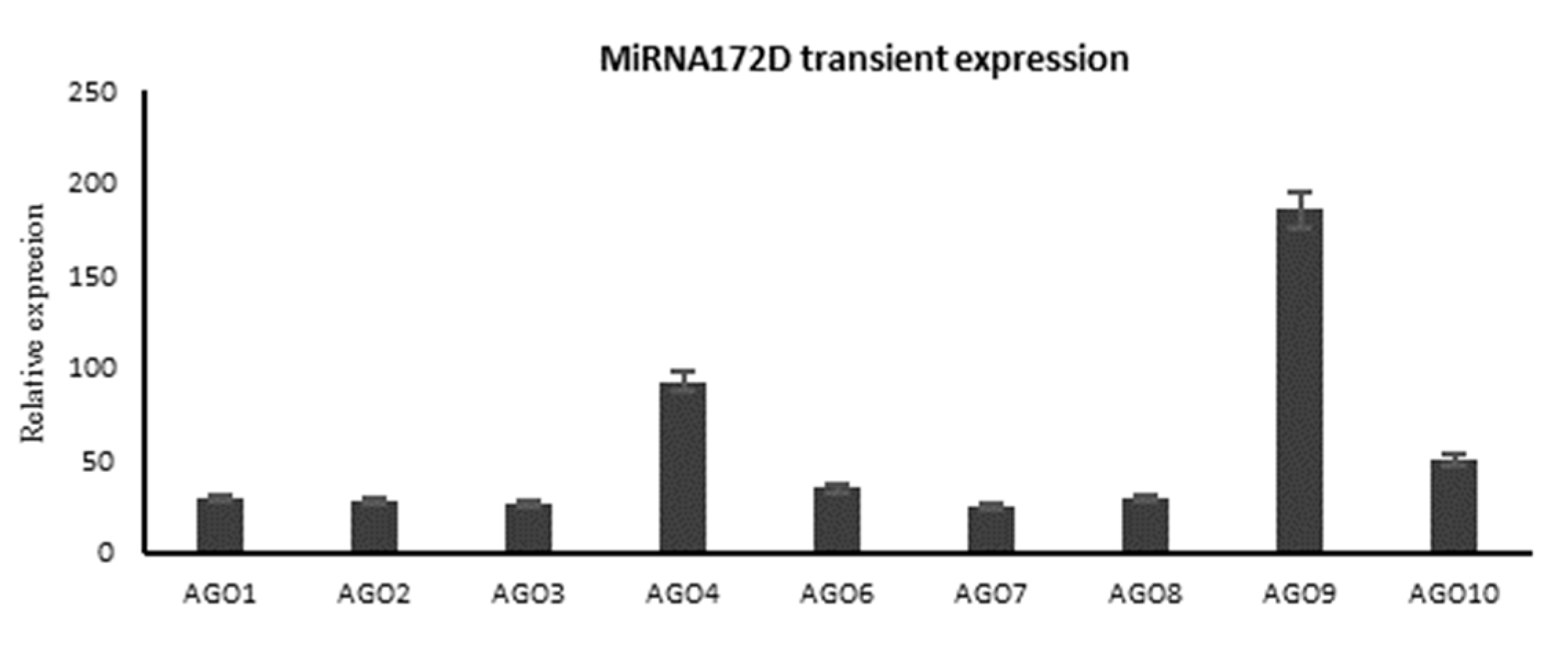
2.9. Transient Overexpression of microRNA172D in Arabidopsis
2.10. Hypothetical Model of CQDs in Plant Bodies and Cells
3. Discussion
4. Materials and Methods
4.1. Experiment Material, Instruments, and Chemical Reagents
4.2. Transcriptome Analysis
4.3. Identification of DEGs
4.4. Subcellular Localization Analysis
4.5. DNA Methylation
4.6. Vector Construction and Transient Transformation
4.7. RNA Extraction and RT-qPCR Analysis
5. Conclusions and Future Prospects
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Javed, T.; Shabbir, R.; Hussain, S.; Naseer, M.A.; Ejaz, I.; Ali, M.M.; Ahmar, S.; Yousef, A.F. Nanotechnology for Endorsing Abiotic Stresses: A Review on the Role of Nanoparticles and Nanocompositions. Funct. Plant Biol. 2022. [Google Scholar] [CrossRef] [PubMed]
- Abdelghany, A.M.; El-Banna, A.A.A.; Salama, E.A.A.; Ali, M.M.; Al-Huqail, A.A.; Ali, H.M.; Paszt, L.S.; El-Sorady, G.A.; Lamlom, S.F. The Individual and Combined Effect of Nanoparticles and Biofertilizers on Growth, Yield, and Biochemical Attributes of Peanuts (Arachis Hypogea L.). Agronomy 2022, 12, 398. [Google Scholar] [CrossRef]
- Das, R.; Bandyopadhyay, R.; Pramanik, P. Carbon Quantum Dots from Natural Resource: A Review. Mater. Today Chem. 2018, 8, 96–109. [Google Scholar] [CrossRef]
- Lim, S.Y.; Shen, W.; Gao, Z. Carbon Quantum Dots and Their Applications. Chem. Soc. Rev. 2015, 44, 362–381. [Google Scholar] [CrossRef]
- Khodakovskaya, M.V.; de Silva, K.; Biris, A.S.; Dervishi, E.; Villagarcia, H. Carbon Nanotubes Induce Growth Enhancement of Tobacco Cells. ACS Nano 2012, 6, 2128–2135. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.-P.; Zhou, B.; Lin, Y.; Wang, W.; Fernando, K.A.S.; Pathak, P.; Meziani, M.J.; Harruff, B.A.; Wang, X.; Wang, H.; et al. Quantum-Sized Carbon Dots for Bright and Colorful Photoluminescence. J. Am. Chem. Soc. 2006, 128, 7756–7757. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Zhang, J.; Li, Z.; Wu, M. Hydrothermal Route for Cutting Graphene Sheets into Blue-Luminescent Graphene Quantum Dots. Adv. Mater. 2010, 22, 734–738. [Google Scholar] [CrossRef]
- Torney, F.; Trewyn, B.G.; Lin, V.S.-Y.; Wang, K. Mesoporous Silica Nanoparticles Deliver DNA and Chemicals into Plants. Nat. Nanotechnol. 2007, 2, 295–300. [Google Scholar] [CrossRef]
- Michalet, X.; Pinaud, F.F.; Bentolila, L.A.; Tsay, J.M.; Doose, S.; Li, J.J.; Sundaresan, G.; Wu, A.M.; Gambhir, S.S.; Weiss, S. Quantum Dots for Live Cells, in Vivo Imaging, and Diagnostics. Science 2005, 307, 538–544. [Google Scholar] [CrossRef]
- Tripathi, S.; Sarkar, S. Influence of Water Soluble Carbon Dots on the Growth of Wheat Plant. Appl. Nanosci. 2015, 5, 609–616. [Google Scholar] [CrossRef]
- Park, S.Y.; Lee, H.U.; Park, E.S.; Lee, S.C.; Lee, J.-W.; Jeong, S.W.; Kim, C.H.; Lee, Y.-C.; Huh, Y.S.; Lee, J. Photoluminescent Green Carbon Nanodots from Food-Waste-Derived Sources: Large-Scale Synthesis, Properties, and Biomedical Applications. ACS Appl. Mater. Interfaces 2014, 6, 3365–3370. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zheng, Y.; Zhang, H.; Liu, Z.; Su, W.; Chen, S.; Liu, Y.; Zhuang, J.; Lei, B. Phytotoxicity, Uptake, and Translocation of Fluorescent Carbon Dots in Mung Bean Plants. ACS Appl. Mater. Interfaces 2016, 8, 19939–19945. [Google Scholar] [CrossRef] [PubMed]
- Nair, R.; Poulose, A.C.; Nagaoka, Y.; Yoshida, Y.; Maekawa, T.; Kumar, D.S. Uptake of FITC Labeled Silica Nanoparticles and Quantum Dots by Rice Seedlings: Effects on Seed Germination and Their Potential as Biolabels for Plants. J. Fluoresc. 2011, 21, 2057–2068. [Google Scholar] [CrossRef]
- Azeem, I.; Adeel, M.; Ahmad, M.A.; Shakoor, N.; Jiangcuo, G.D.; Azeem, K.; Ishfaq, M.; Shakoor, A.; Ayaz, M.; Xu, M.; et al. Uptake and Accumulation of Nano/Microplastics in Plants: A Critical Review. Nanomaterials 2021, 11, 2935. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gerstein, M.; Snyder, M. RNA-Seq: A Revolutionary Tool for Transcriptomics. Nat. Rev. Genet. 2009, 10, 57–63. [Google Scholar] [CrossRef]
- Steinitz-Sears, L.M. Chromosome Studies in Arabidopsis Thaliana. Genetics 1963, 48, 483–490. [Google Scholar] [CrossRef]
- Röbbelen, G. Arabidopsis Research. Science 1965, 150, 1192. [Google Scholar] [CrossRef]
- Meyerowitz, E.M. Arabidopsis Thaliana. Annu. Rev. Genet. 1987, 21, 93–111. [Google Scholar] [CrossRef]
- Baulcombe, D. RNA Silencing in Plants. Nature 2004, 431, 356–363. [Google Scholar] [CrossRef]
- Hamilton, A.J.; Baulcombe, D.C. A Species of Small Antisense RNA in Posttranscriptional Gene Silencing in Plants. Science 1999, 286, 950–952. [Google Scholar] [CrossRef]
- Bartel, D.P. MicroRNAs: Genomics, Biogenesis, Mechanism, and Function. Cell 2004, 116, 281–297. [Google Scholar] [CrossRef] [PubMed]
- Wassenegger, M.; Heimes, S.; Riedel, L.; Sänger, H.L. RNA-Directed de Novo Methylation of Genomic Sequences in Plants. Cell 1994, 76, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Vaucheret, H. Plant ARGONAUTES. Trends Plant Sci. 2008, 13, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Mallory, A.; Vaucheret, H. Form, Function, and Regulation of ARGONAUTE Proteins. Plant Cell 2010, 22, 3879–3889. [Google Scholar] [CrossRef]
- Li, Z.; Li, W.; Guo, M.; Liu, S.; Liu, L.; Yu, Y.; Mo, B.; Chen, X.; Gao, L. Origin, Evolution and Diversification of Plant ARGONAUTE Proteins. Plant J. 2022, 109, 1086–1097. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, X.; Liu, J.; Kiba, T.; Woo, J.; Ojo, T.; Hafner, M.; Tuschl, T.; Chua, N.; Wang, X. Deep Sequencing of Small RNAs Specifically Associated with Arabidopsis AGO1 and AGO4 Uncovers New AGO Functions. Plant J. 2011, 67, 292–304. [Google Scholar] [CrossRef]
- Zhang, Z.; Liu, X.; Guo, X.; Wang, X.-J.; Zhang, X. Arabidopsis AGO3 Predominantly Recruits 24-Nt Small RNAs to Regulate Epigenetic Silencing. Nat. Plants 2016, 2, 16049. [Google Scholar] [CrossRef]
- Gan, D.; Zhan, M.; Yang, F.; Zhang, Q.; Hu, K.; Xu, W.; Lu, Q.; Zhang, L.; Liang, D. Expression Analysis of Argonaute, Dicer-like, and RNA-Dependent RNA Polymerase Genes in Cucumber (Cucumis Sativus L.) in Response to Abiotic Stress. J. Genet. 2017, 96, 235–249. [Google Scholar] [CrossRef]
- Ma, W.; Wu, F.; Sheng, P.; Wang, X.; Zhang, Z.; Zhou, K.; Zhang, H.; Hu, J.; Lin, Q.; Cheng, Z.; et al. The LBD12-1 Transcription Factor Suppresses Apical Meristem Size by Repressing Argonaute 10 Expression. Plant Physiol. 2017, 173, 801–811. [Google Scholar] [CrossRef]
- Wang, B.; Duan, C.; Wang, X.; Hou, Y.; Yan, J.; Gao, C.; Kim, J.; Zhang, H.; Zhu, J. HOS1 Regulates Argonaute1 by Promoting Transcription of the MicroRNA Gene MIR168b in Arabidopsis. Plant J. 2015, 81, 861–870. [Google Scholar] [CrossRef]
- Schweizer, D. Reverse Fluorescent Chromosome Banding with Chromomycin and DAPI. Chromosoma 1976, 58, 307–324. [Google Scholar] [CrossRef]
- Williamson, D.H.; Fennell, D.J. The Use of Fluorescent DNA-Binding Agent for Detecting and Separating Yeast Mitochondrial DNA. In Methods in Cell Biology; Academic Press: Cambridge, MA, USA, 1975; pp. 335–351. [Google Scholar]
- Li, H.; Shi, W.; Huang, W.; Yao, E.-P.; Han, J.; Chen, Z.; Liu, S.; Shen, Y.; Wang, M.; Yang, Y. Carbon Quantum Dots/TiO x Electron Transport Layer Boosts Efficiency of Planar Heterojunction Perovskite Solar Cells to 19%. Nano Lett. 2017, 17, 2328–2335. [Google Scholar] [CrossRef]
- Dong, Y.; Chen, C.; Lin, J.; Zhou, N.; Chi, Y.; Chen, G. Electrochemiluminescence Emission from Carbon Quantum Dot-Sulfite Coreactant System. Carbon N. Y. 2013, 56, 12–17. [Google Scholar] [CrossRef]
- Ai, L.; Song, Z.; Nie, M.; Yu, J.; Liu, F.; Song, H.; Zhang, B.; Waterhouse, G.I.N.; Lu, S. Solid-state Fluorescence from Carbon Dots Widely Tunable from Blue to Deep Red through Surface Ligand Modulation. Angew. Chem. Int. Ed. 2023, 62, e202217822. [Google Scholar] [CrossRef] [PubMed]
- Qu, S.; Zhou, D.; Li, D.; Ji, W.; Jing, P.; Han, D.; Liu, L.; Zeng, H.; Shen, D. Toward Efficient Orange Emissive Carbon Nanodots through Conjugated Sp 2 -Domain Controlling and Surface Charges Engineering. Adv. Mater. 2016, 28, 3516–3521. [Google Scholar] [CrossRef] [PubMed]
- Xu, Q.; Pu, P.; Zhao, J.; Dong, C.; Gao, C.; Chen, Y.; Chen, J.; Liu, Y.; Zhou, H. Preparation of Highly Photoluminescent Sulfur-Doped Carbon Dots for Fe (III) Detection. J. Mater. Chem. A 2015, 3, 542–546. [Google Scholar] [CrossRef]
- Yang, G.; Wan, X.; Liu, Y.; Li, R.; Su, Y.; Zeng, X.; Tang, J. Luminescent Poly(Vinyl Alcohol)/Carbon Quantum Dots Composites with Tunable Water-Induced Shape Memory Behavior in Different PH and Temperature Environments. ACS Appl. Mater. Interfaces 2016, 8, 34744–34754. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Wang, Z.; Li, X.; Li, Y.; Tan, Z.; Fan, L.; Yang, S. Bright Multicolor Bandgap Fluorescent Carbon Quantum Dots for Electroluminescent Light-Emitting Diodes. Adv. Mater. 2017, 29, 1604436. [Google Scholar] [CrossRef]
- Zheng, B.; Liu, X.; Hu, J.; Wang, F.; Hu, X.; Zhu, Y.; Lv, X.; Du, J.; Xiao, D. Construction of Hydrophobic Interface on Natural Biomaterials for Higher Efficient and Reversible Radioactive Iodine Adsorption in Water. J. Hazard. Mater. 2019, 368, 81–89. [Google Scholar] [CrossRef]
- Hsu, P.-C.; Chang, H.-T. Synthesis of High-Quality Carbon Nanodots from Hydrophilic Compounds: Role of Functional Groups. Chem. Commun. 2012, 48, 3984. [Google Scholar] [CrossRef]
- Yan, M.; Zhou, M.; Chen, J.; Zhao, T.; Tang, L.; Bi, H. Fluorescent CDs@PCL Hybrids via Tartaric Acid, CDs-Cocatalyzed Polymerization. Mater. Sci. Eng. C 2017, 79, 76–83. [Google Scholar] [CrossRef]
- Wiśniewski, M. The Consequences of Water Interactions with Nitrogen-Containing Carbonaceous Quantum Dots—The Mechanistic Studies. Int. J. Mol. Sci. 2022, 23, 14292. [Google Scholar] [CrossRef]
- Stillwell, W. Membrane Transport. In An Introduction to Biological Membranes; Elsevier: Amsterdam, The Netherlands, 2016; pp. 423–451. [Google Scholar]
- You, S.; Cai, Q.; Zheng, Y.; He, B.; Shen, J.; Yang, W.; Yin, M. Perylene-Cored Star-Shaped Polycations for Fluorescent Gene Vectors and Bioimaging. ACS Appl. Mater. Interfaces 2014, 6, 16327–16334. [Google Scholar] [CrossRef]
- Dong, D.; Zhang, J.; Zhang, R.; Li, F.; Li, Y.; Jia, Y. Multiprobe Assay for Clinical SEPT9 Methylation Based on the Carbon Dot-Modified Liquid-Exfoliated Graphene Field Effect Transistor with a Potential to Present a Methylation Panorama. ACS Omega 2020, 5, 16228–16237. [Google Scholar] [CrossRef] [PubMed]
- Gomes, F.M.; Ramos, I.B.; Wendt, C.; Girard-Dias, W.; De Souza, W.; Machado, E.A.; Miranda, K. New Insights into the in Situ Microscopic Visualization and Quantification of Inorganic Polyphosphate Stores by 4′,6-Diamidino-2-Phenylindole (DAPI)-Staining. Eur. J. Histochem. 2013, 57, 34. [Google Scholar] [CrossRef]
- Loukanov, A.; Mladenova, P.; Toshev, S.; Karailiev, A.; Ustinovich, E.; Nakabayashi, S. Real Time Monitoring and Quantification of Uptake Carbon Nanodots in Eukaryotic Cells. Microsc. Res. Tech. 2018, 81, 1541–1547. [Google Scholar] [CrossRef]
- Chandrakar, V.; Yadu, B.; Korram, J.; Satnami, M.L.; Dubey, A.; Kumar, M.; Keshavkant, S. Carbon Dot Induces Tolerance to Arsenic by Regulating Arsenic Uptake, Reactive Oxygen Species Detoxification and Defense-Related Gene Expression in Cicer Arietinum L. Plant Physiol. Biochem. 2020, 156, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Yakusheva, A.; Aly-Eldeen, M.; Gusev, A.; Zakharova, O.; Kuznetsov, D. Cyan Fluorescent Carbon Quantum Dots with Amino Derivatives for the Visual Detection of Copper (II) Cations in Sea Water. Nanomaterials 2023, 13, 1004. [Google Scholar] [CrossRef] [PubMed]
- Le, N.; Zhang, M.; Kim, K. Quantum Dots and Their Interaction with Biological Systems. Int. J. Mol. Sci. 2022, 23, 10763. [Google Scholar] [CrossRef]
- Gao, M.; Chang, J.; Wang, Z.; Zhang, H.; Wang, T. Advances in Transport and Toxicity of Nanoparticles in Plants. J. Nanobiotechnology 2023, 21, 75. [Google Scholar] [CrossRef]
- Wiśniewski, M.; Czarnecka, J.; Bolibok, P.; Świdziński, M.; Roszek, K. New Insight into the Fluorescence Quenching of Nitrogen-Containing Carbonaceous Quantum Dots—From Surface Chemistry to Biomedical Applications. Materials 2021, 14, 2454. [Google Scholar] [CrossRef] [PubMed]
- Wei, T.; Deng, K.; Wang, H.; Zhang, L.; Wang, C.; Song, W.; Zhang, Y.; Chen, C. Comparative Transcriptome Analyses Reveal Potential Mechanisms of Enhanced Drought Tolerance in Transgenic Salvia Miltiorrhiza Plants Expressing AtDREB1A from Arabidopsis. Int. J. Mol. Sci. 2018, 19, 827. [Google Scholar] [CrossRef] [PubMed]
- Allan, R.A.; Miller, J.J. Influence of S -Adenosylmethionine on DAPI-Induced Fluorescence of Polyphosphate in the Yeast Vacuole. Can. J. Microbiol. 1980, 26, 912–920. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Liu, Q.; Chen, B.; Wang, Q.; Shi, X.; Xiao, Z.; Lin, J.; Fang, X. Carbon Nanotubes as Molecular Transporters for Walled Plant Cells. Nano Lett. 2009, 9, 1007–1010. [Google Scholar] [CrossRef] [PubMed]
- Santana, I.; Jeon, S.-J.; Kim, H.-I.; Islam, M.R.; Castillo, C.; Garcia, G.F.H.; Newkirk, G.M.; Giraldo, J.P. Targeted Carbon Nanostructures for Chemical and Gene Delivery to Plant Chloroplasts. ACS Nano 2022, 16, 12156–12173. [Google Scholar] [CrossRef]
- Rezaei Cherati, S.; Anas, M.; Liu, S.; Shanmugam, S.; Pandey, K.; Angtuaco, S.; Shelton, R.; Khalfaoui, A.N.; Alena, S.V.; Porter, E.; et al. Comprehensive Risk Assessment of Carbon Nanotubes Used for Agricultural Applications. ACS Nano 2022, 16, 12061–12072. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, Z.; Ali, M.M.; Yi, X.; Zhang, L.; Wang, S.; Chen, F. Unlocking the Potential of Carbon Quantum Dots for Cell Imaging, Intracellular Localization, and Gene Expression Control in Arabidopsis thaliana (L.) Heynh. Int. J. Mol. Sci. 2023, 24, 15700. https://doi.org/10.3390/ijms242115700
Lin Z, Ali MM, Yi X, Zhang L, Wang S, Chen F. Unlocking the Potential of Carbon Quantum Dots for Cell Imaging, Intracellular Localization, and Gene Expression Control in Arabidopsis thaliana (L.) Heynh. International Journal of Molecular Sciences. 2023; 24(21):15700. https://doi.org/10.3390/ijms242115700
Chicago/Turabian StyleLin, Zhimin, Muhammad Moaaz Ali, Xiaoyan Yi, Lijuan Zhang, Shaojuan Wang, and Faxing Chen. 2023. "Unlocking the Potential of Carbon Quantum Dots for Cell Imaging, Intracellular Localization, and Gene Expression Control in Arabidopsis thaliana (L.) Heynh." International Journal of Molecular Sciences 24, no. 21: 15700. https://doi.org/10.3390/ijms242115700
APA StyleLin, Z., Ali, M. M., Yi, X., Zhang, L., Wang, S., & Chen, F. (2023). Unlocking the Potential of Carbon Quantum Dots for Cell Imaging, Intracellular Localization, and Gene Expression Control in Arabidopsis thaliana (L.) Heynh. International Journal of Molecular Sciences, 24(21), 15700. https://doi.org/10.3390/ijms242115700