A High-K+ Affinity Transporter (HKT) from Actinidia valvata Is Involved in Salt Tolerance in Kiwifruit

 , , and
, , and
Abstract
:1. Introduction
2. Results
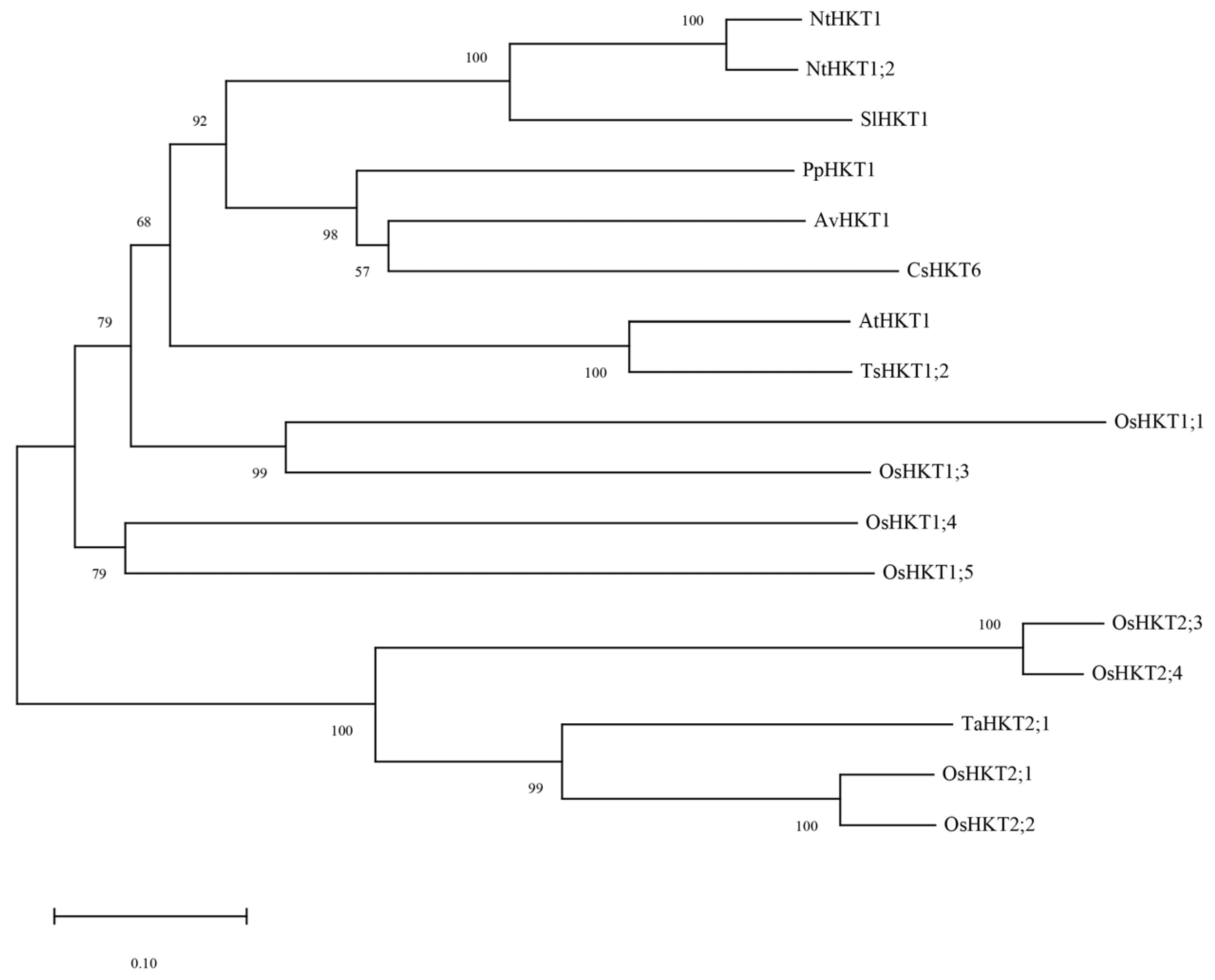
2.1. Phylogenetic Tree Analysis for AvHKT1
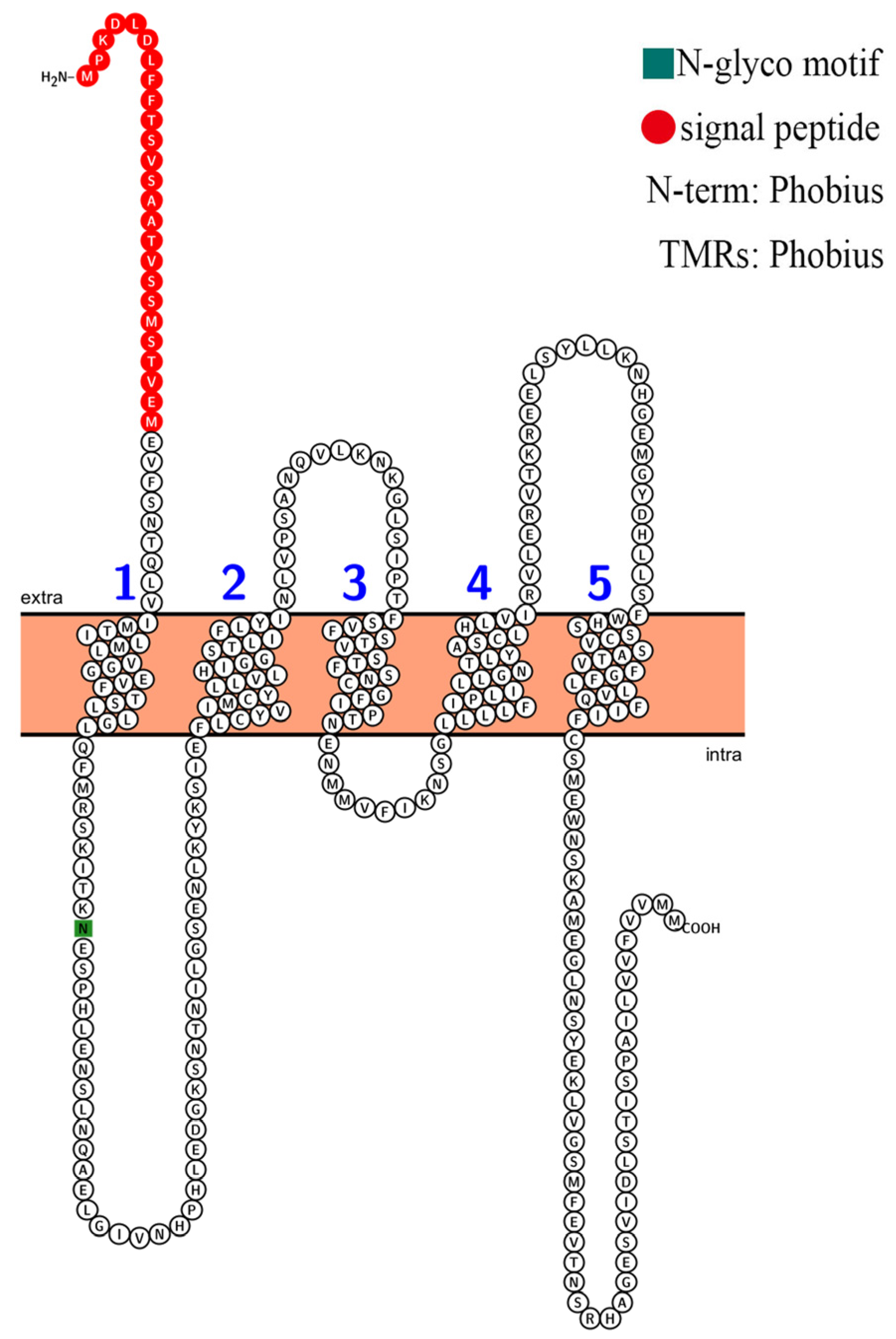
2.2. Conserved Sites and Trans-Membrane Domains for AvHKT1
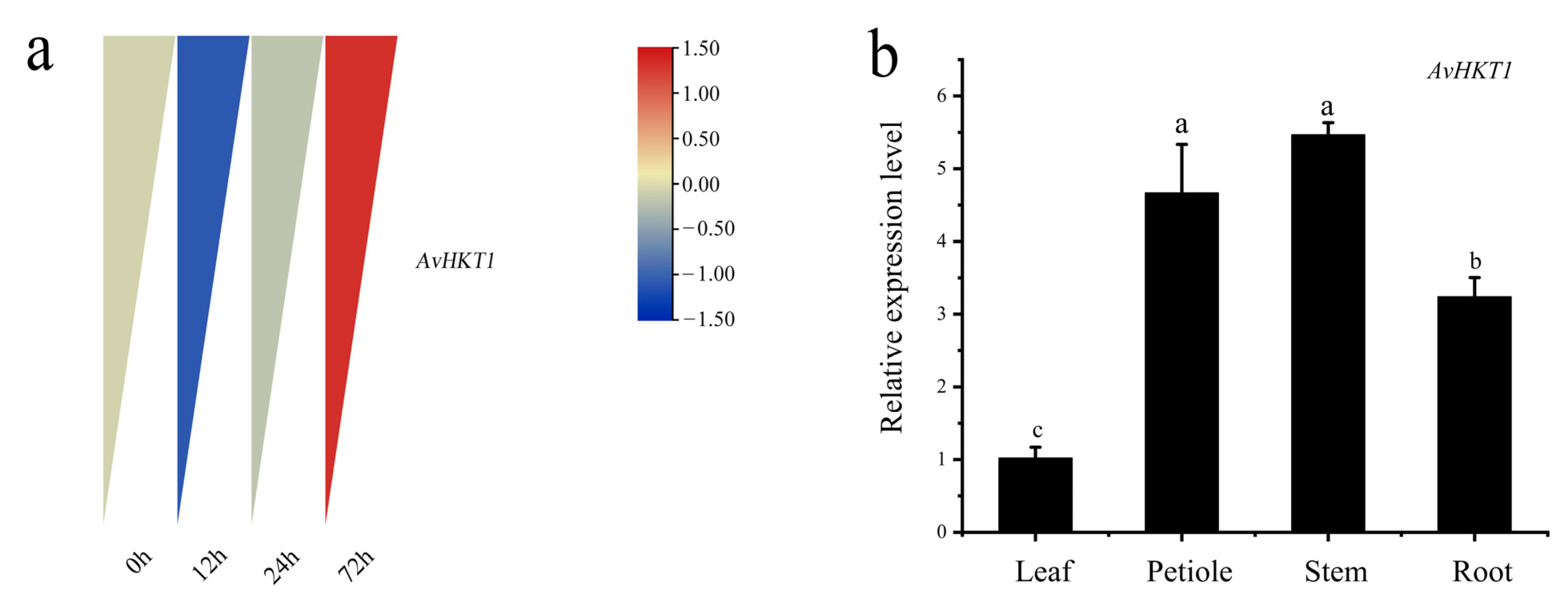
2.3. The Expression Pattern of AvHKT1
2.4. Upstream Promoter Region Analysis of AvHKT1
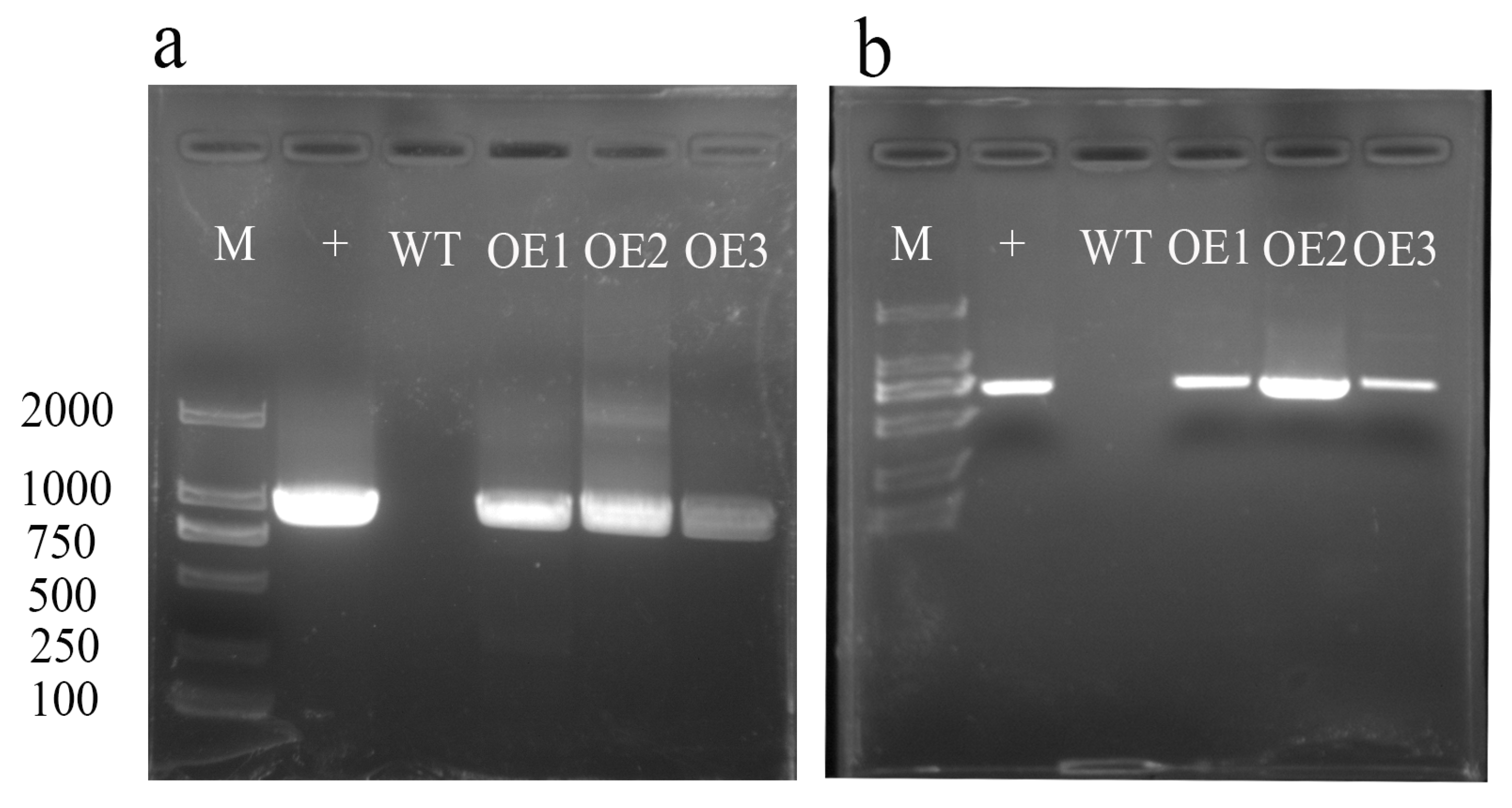
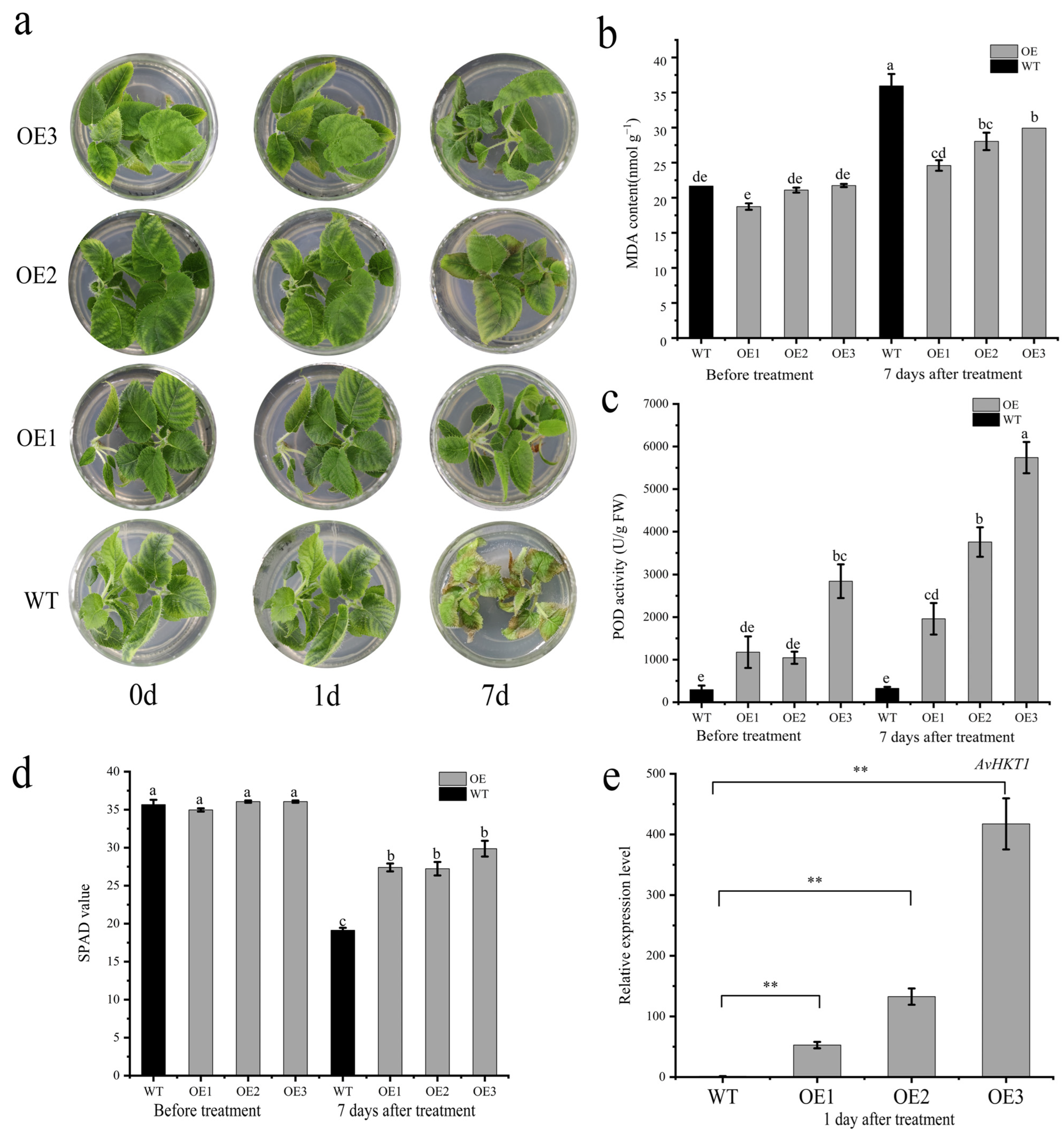
2.5. Overexpression of AvHKT1 Enhances Salt Stress Tolerance of A. chinensis
3. Discussion
4. Materials and Methods
4.1. Plant Material and Stress Treatment
4.2. Sequence Identification and Structure Analysis
4.3. Total RNA Extraction and RT-qPCR Analysis
4.4. Cloning CDS and Promoter Sequences of AvHKT1
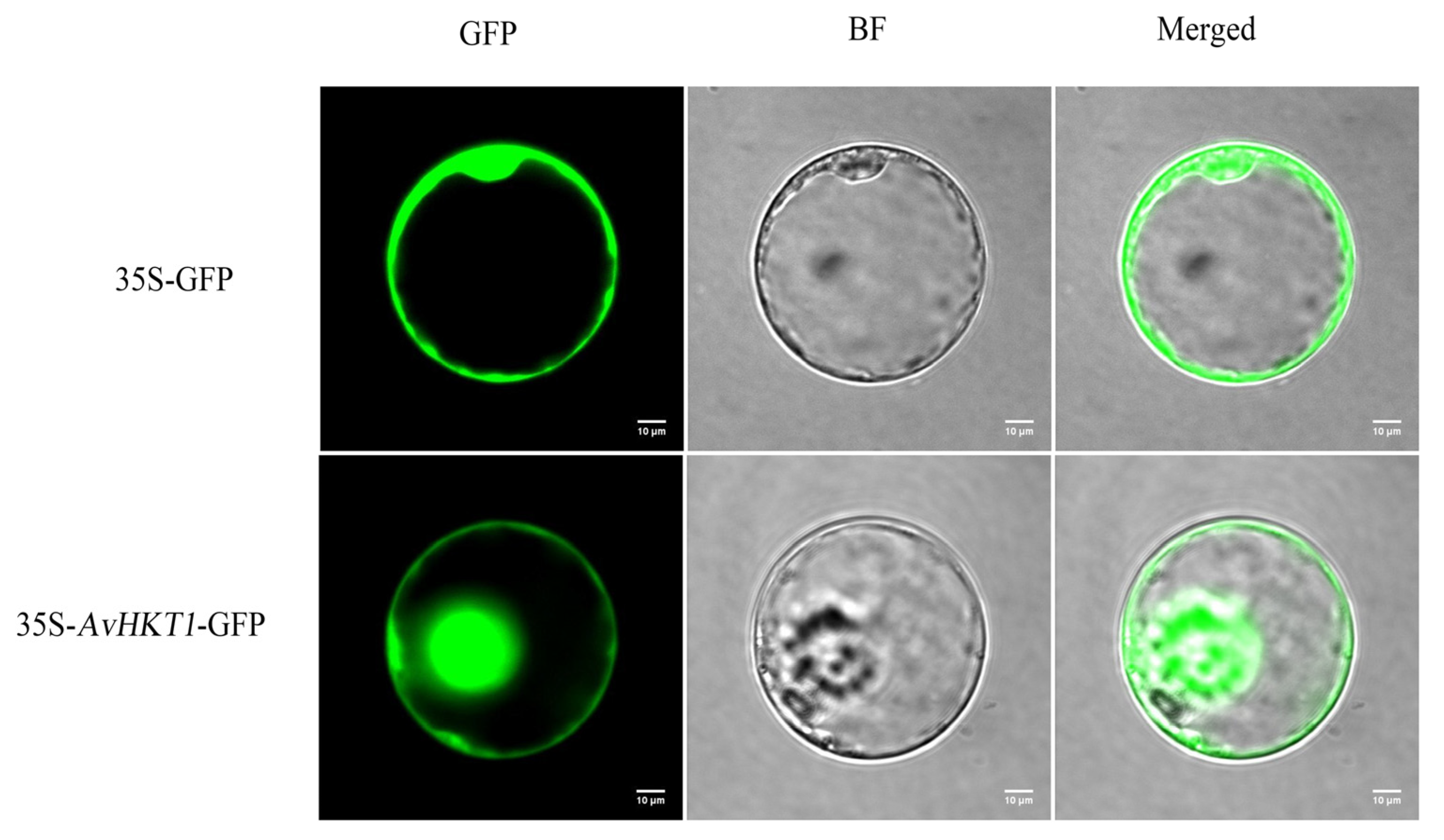
4.5. Subcellular Localization Analysis of AvHKT1
4.6. Transformation of Kiwifruit
4.7. Physiological Analysis
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hauser, F.; Horie, T. A conserved primary salt tolerance mechanism mediated by HKT transporters: A mechanism for sodium exclusion and maintenance of high K+/Na+ ratio in leaves during salinity stress. Plant Cell Environ. 2010, 33, 552–565. [Google Scholar] [CrossRef] [PubMed]
- Hamamoto, S.; Horie, T.; Hauser, F.; Deinlein, U.; Schroeder, J.I.; Uozumi, N. HKT transporters mediate salt stress resistance in plants: From structure and function to the field. Curr. Opin. Biotechnol. 2015, 32, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Maggio, A.; Bressan, R.A.; Yun, D.J. Role and Functional Differences of HKT1-Type Transporters in Plants under Salt Stress. Int. J. Mol. Sci. 2019, 20, 1059. [Google Scholar] [CrossRef]
- Kato, Y.; Sakaguchi, M.; Mori, Y.; Saito, K.; Nakamura, T.; Bakker, E.P.; Sato, Y.; Goshima, S.; Uozumi, N. Evidence in support of a four transmembrane-pore-transmembrane topology model for the Arabidopsis thaliana Na+/K+ translocating AtHKT1 protein, a member of the superfamily of K+ transporters. PNAS 2001, 98, 6488–6493. [Google Scholar] [CrossRef]
- Mäser, P.; Hosoo, Y.; Goshima, S.; Horie, T.; Eckelman, B.; Yamada, K.; Yoshida, K.; Bakker, E.P.; Shinmyo, A.; Oiki, S.; et al. Glycine residues in potassium channel-like selectivity filters determine potassium selectivity in four-loop-per-subunit HKT transporters from plants. PNAS 2002, 99, 6428–6433. [Google Scholar] [CrossRef]
- Durell, S.R.; Guy, H.R. Structural models of the KtrB, TrkH, and Trk1,2 symporters based on the structure of the KcsA K+ channel. Biophys. J. 1999, 77, 789–807. [Google Scholar] [CrossRef] [PubMed]
- Durell, S.R.; Hao, Y.; Nakamura, T.; Bakker, E.P.; Guy, H.R. Evolutionary Relationship between K+ Channels and Symporters. Biophys. J. 1999, 77, 775–788. [Google Scholar] [CrossRef]
- Horie, T.; Costa, A.; Kim, T.H.; Han, M.J.; Horie, R.; Leung, H.Y.; Miyao, A.; Hirochika, H.; An, G.; Schroeder, J.I. Rice OsHKT2;1 transporter mediates large Na+ influx component into K+-starved roots for growth. EMBO J. 2007, 26, 3003–3014. [Google Scholar] [CrossRef]
- Horie, T.; Yoshida, K.; Nakayama, H.; Yamada, K.; Oiki, S.; Shinmyo, A. Two types of HKT transporters with different properties of Na+ and K+ transport in Oryza sativa. Plant J. 2001, 27, 129–138. [Google Scholar] [CrossRef]
- Rubio, F.; Gassmann, W.; Schroeder, J.I. Sodium-driven potassium uptake by the plant potassium transporter HKT1 and mutations conferring salt tolerance. Science 1995, 270, 1660–1663. [Google Scholar] [CrossRef]
- Fu, L.; Shen, Q.; Kuang, L.; Yu, J.; Wu, D.; Zhang, G. Metabolite profiling and gene expression of Na/K transporter analyses reveal mechanisms of the difference in salt tolerance between barley and rice. Plant Physiol. Biochem. 2018, 130, 248–257. [Google Scholar] [CrossRef]
- van Bezouw, R.; Janssen, E.M.; Ashrafuzzaman, M.; Ghahramanzadeh, R.; Kilian, B.; Graner, A.; Visser, R.G.F.; van der Linden, C.G. Shoot sodium exclusion in salt stressed barley (Hordeum vulgare L.) is determined by allele specific increased expression of HKT1;5. J. Plant Physiol. 2019, 241, 153029. [Google Scholar] [CrossRef] [PubMed]
- Oda, Y.; Kobayashi, N.I.; Tanoi, K.; Ma, J.F.; Itou, Y.; Katsuhara, M.; Itou, T.; Horie, T. T-DNA Tagging-Based Gain-of-Function of OsHKT1;4 Reinforces Na Exclusion from Leaves and Stems but Triggers Na Toxicity in Roots of Rice Under Salt Stress. Int. J. Mol. Sci. 2018, 19, 235. [Google Scholar] [CrossRef] [PubMed]
- Mäser, P.; Eckelman, B.; Vaidyanathan, R.; Horie, T.; Fairbairn, D.J.; Kubo, M.; Yamagami, M.; Yamaguchi, K.; Nishimura, M.; Uozumi, N.; et al. Altered shoot/root Na+ distribution and bifurcating salt sensitivity in Arabidopsis by genetic disruption of the Na+ transporter AtHKT1. FEBS Lett. 2002, 531, 157–161. [Google Scholar] [CrossRef]
- Berthomieu, P.; Conéjéro, G.; Nublat, A.; Brackenbury, W.J.; Lambert, C.; Savio, C.; Uozumi, N.; Oiki, S.; Yamada, K.; Cellier, F.; et al. Functional analysis of AtHKT1 in Arabidopsis shows that Na+ recirculation by the phloem is crucial for salt tolerance. EMBO J. 2003, 22, 2004–2014. [Google Scholar] [CrossRef]
- Sunarpi; Horie, T.; Motoda, J.; Kubo, M.; Yang, H.; Yoda, K.; Horie, R.; Chan, W.-Y.; Leung, H.-Y.; Hattori, K.; et al. Enhanced salt tolerance mediated by AtHKT1 transporter-induced Na+ unloading from xylem vessels to xylem parenchyma cells. Plant J. 2005, 44, 928–938. [Google Scholar] [CrossRef]
- Hill, C.B.; Jha, D.; Bacic, A.; Tester, M.; Roessner, U. Characterization of ion contents and metabolic responses to salt stress of different Arabidopsis AtHKT1;1 genotypes and their parental strains. Mol. Plant 2013, 6, 350–368. [Google Scholar] [CrossRef]
- Ali, Z.; Park, H.C.; Ali, A.; Oh, D.H.; Aman, R.; Kropornicka, A.; Hong, H.; Choi, W.; Chung, W.S.; Kim, W.Y.; et al. TsHKT1;2, a HKT1 Homolog from the Extremophile Arabidopsis Relative Thellungiella salsuginea, Shows K+ Specificity in the Presence of NaCl. Plant Physiol. 2012, 158, 1463–1474. [Google Scholar] [CrossRef]
- Wang, L.; Liu, Y.; Li, D.; Feng, S.; Yang, J.; Zhang, J.; Zhang, J.; Wang, D.; Gan, Y. Improving salt tolerance in potato through overexpression of AtHKT1 gene. BMC Plant Biol. 2019, 19, 357–372. [Google Scholar] [CrossRef]
- Byrt, C.S.; Xu, B.; Krishnan, M.; Lightfoot, D.J.; Athman, A.; Jacobs, A.K.; Watson Haigh, N.S.; Plett, D.; Munns, R.; Tester, M.; et al. The Na+ transporter, TaHKT1;5-D, limits shoot Na+ accumulation in bread wheat. Plant J. 2014, 80, 516–526. [Google Scholar] [CrossRef]
- Cao, Y.; Liang, X.; Yin, P.; Zhang, M.; Jiang, C. A domestication-associated reduction in K+ -preferring HKT transporter activity underlies maize shoot K+ accumulation and salt tolerance. New Phytol. 2019, 222, 301–317. [Google Scholar] [CrossRef]
- Zhang, M.; Cao, Y.; Wang, Z.; Wang, Z.Q.; Shi, J.; Liang, X.; Song, W.; Chen, Q.; Lai, J.; Jiang, C. A retrotransposon in an HKT1 family sodium transporter causes variation of leaf Na+ exclusion and salt tolerance in maize. New Phytol. 2018, 217, 1161–1176. [Google Scholar] [CrossRef]
- Henderson, S.W.; Dunlevy, J.D.; Wu, Y.; Blackmore, D.H.; Walker, R.R.; Edwards, E.J.; Gilliham, M.; Walker, A.R. Functional differences in transport properties of natural HKT1;1 variants influence shoot Na+ exclusion in grapevine rootstocks. New Phytol. 2018, 217, 1113–1127. [Google Scholar] [CrossRef]
- Wu, Y.; Henderson, S.W.; Wege, S.; Zheng, F.; Walker, A.R.; Walker, R.R.; Gilliham, M. The grapevine NaE sodium exclusion locus encodes sodium transporters with diverse transport properties and localisation. J. Plant Physiol. 2020, 246–247, 153113. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; He, H.; Yu, D. Overexpression of a novel soybean gene modulating Na+ and K+ transport enhances salt tolerance in transgenic tobacco plants. Physiol. Plant. 2011, 141, 11–18. [Google Scholar] [CrossRef]
- Kawakami, Y.; Imran, S.; Katsuhara, M.; Tada, Y. Na+ Transporter SvHKT1;1 from a Halophytic Turf Grass Is Specifically Upregulated by High Na+ Concentration and Regulates Shoot Na+ Concentration. Int. J. Mol. Sci. 2020, 21, 6100. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Cao, H.; Cheng, J.; He, X.; Sohail, H.; Niu, M.; Huang, Y.; Bie, Z. Pumpkin CmHKT1;1 Controls Shoot Na⁺ Accumulation via Limiting Na⁺ Transport from Rootstock to Scion in Grafted Cucumber. Int. J. Mol. Sci. 2018, 19, 2648. [Google Scholar] [CrossRef]
- Ukwatta, J.; Pabuayon, I.C.M.; Park, J.; Chen, J.; Chai, X.; Zhang, H.; Zhu, J.K.; Xin, Z.; Shi, H. Comparative physiological and transcriptomic analysis reveals salinity tolerance mechanisms in Sorghum bicolor (L.) Moench. Planta 2021, 254, 98–118. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.T.; Ren, Z.J.; Liu, Z.Q.; Feng, X.; Guo, R.Q.; Li, B.G.; Li, L.G.; Jing, H.C. SbHKT1;4, a member of the high-affinity potassium transporter gene family from Sorghum bicolor, functions to maintain optimal Na+/K+ balance under Na+ stress. J. Integr. Plant Biol. 2014, 56, 315–332. [Google Scholar] [CrossRef] [PubMed]
- Nawaz, I.; Iqbal, M.; Hakvoort, H.W.J.; de Boer, A.H.; Schat, H. Analysis of Arabidopsis thaliana HKT1 and Eutrema salsugineum/botschantzevii HKT1;2 Promoters in Response to Salt Stress in Athkt1:1 Mutant. Mol. Biotechnol. 2019, 61, 442–450. [Google Scholar] [CrossRef]
- Zamani Babgohari, M.; Ebrahimie, E.; Niazi, A. In silico analysis of high affinity potassium transporter (HKT) isoforms in different plants. Aquat. Biosyst. 2014, 10, 9–23. [Google Scholar] [CrossRef] [PubMed]
- Tounsi, S.; Saïdi, M.N.; Abdelhedi, R.; Feki, K.; Bahloul, N.; Alcon, C.; Masmoudi, K.; Brini, F. Functional analysis of TmHKT1;4-A2 promoter through deletion analysis provides new insight into the regulatory mechanism underlying abiotic stress adaptation. Planta 2021, 253, 18–31. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Jing, W.; Xiao, L.; Jin, Y.; Shen, L.; Zhang, W. The Rice High-Affinity Potassium Transporter1;1 Is Involved in Salt Tolerance and Regulated by an MYB-Type Transcription Factor. Plant Physiol. 2015, 168, 1076–1090. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.C.; Lim, S.D.; Moon, J.C.; Jang, C.S. A rice really interesting new gene H2-type E3 ligase, OsSIRH2-14, enhances salinity tolerance via ubiquitin/26S proteasome-mediated degradation of salt-related proteins. Plant Cell Environ. 2019, 42, 3061–3076. [Google Scholar] [CrossRef]
- Abid, M.; Gu, S.; Zhang, Y.-J.; Sun, S.; Li, Z.; Bai, D.-F.; Sun, L.; Qi, X.-J.; Zhong, Y.-P.; Fang, J.-B. Comparative transcriptome and metabolome analysis reveal key regulatory defense networks and genes involved in enhanced salt tolerance of Actinidia (kiwifruit). Hortic. Res. 2022, 9, uhac189. [Google Scholar] [CrossRef] [PubMed]
- Shahzad, K.; Rauf, M.; Ahmed, M.; Malik, Z.A.; Habib, I.; Ahmed, Z.; Mahmood, K.; Ali, R.; Masmoudi, K.; Lemtiri-Chlieh, F.; et al. Functional characterisation of an intron retaining K+ transporter of barley reveals intron-mediated alternate splicing. Plant Biol. 2015, 17, 840–851. [Google Scholar] [CrossRef]
- Horie, T.; Hauser, F.; Schroeder, J.I. HKT transporter-mediated salinity resistance mechanisms in Arabidopsis and monocot crop plants. Trends Plant Sci. 2009, 14, 660–668. [Google Scholar] [CrossRef]
- Zhang, H.; Kim, M.-S.; Sun, Y.; Dowd, S.E.; Shi, H.; Paré, P.W. Soil Bacteria Confer Plant Salt Tolerance by Tissue-Specific Regulation of the Sodium Transporter HKT1. Mol. Plant-Microbe Interact. 2008, 21, 737–744. [Google Scholar] [CrossRef]
- Zhang, W.D.; Wang, P.; Bao, Z.; Ma, Q.; Duan, L.J.; Bao, A.K.; Zhang, J.L.; Wang, S.M. SOS1, HKT1;5, and NHX1 Synergistically Modulate Na+ Homeostasis in the Halophytic Grass Puccinellia tenuiflora. Front. Plant Sci. 2017, 8, 576–584. [Google Scholar] [CrossRef]
- Xu, M.; Chen, C.; Cai, H.; Wu, L. Overexpression of PeHKT1;1 Improves Salt Tolerance in Populus. Genes 2018, 9, 475. [Google Scholar] [CrossRef]
- Tounsi, S.; Feki, K.; Saïdi, M.N.; Maghrebi, S.; Brini, F.; Masmoudi, K. Promoter of the TmHKT1;4-A1 gene of Triticum monococcum directs stress inducible, developmental regulated and organ specific gene expression in transgenic Arbidopsis thaliana. World J. Microbiol. Biotechnol. 2018, 34, 99–113. [Google Scholar] [CrossRef]
- Crizel, R.L.; Perin, E.C.; Siebeneichler, T.J.; Borowski, J.M.; Messias, R.S.; Rombaldi, C.V.; Galli, V. Abscisic acid and stress induced by salt: Effect on the phenylpropanoid, L-ascorbic acid and abscisic acid metabolism of strawberry fruits. Plant Physiol. Biochem. 2020, 152, 211–220. [Google Scholar] [CrossRef] [PubMed]
- Waadt, R.; Seller, C.A.; Hsu, P.K.; Takahashi, Y.; Munemasa, S.; Schroeder, J.I. Plant hormone regulation of abiotic stress responses. Nat. Rev. Mol. Cell Biol. 2022, 23, 680–694. [Google Scholar] [CrossRef] [PubMed]
- Ismail, A.; Riemann, M.; Nick, P. The jasmonate pathway mediates salt tolerance in grapevines. J. Exp. Bot. 2012, 63, 2127–2139. [Google Scholar] [CrossRef] [PubMed]
- An, X.; Hao, Y.; Li, E.; Xu, K.; Cheng, C. Functional identification of apple MdJAZ2 in Arabidopsis with reduced JA-sensitivity and increased stress tolerance. Plant Cell Rep. 2017, 36, 255–265. [Google Scholar] [CrossRef]
- Li, W.; Dang, C.; Ye, Y.; Wang, Z.; Hu, B.; Zhang, F.; Zhang, Y.; Qian, X.; Shi, J.; Guo, Y.; et al. Overexpression of Grapevine VvIAA18 Gene Enhanced Salt Tolerance in Tobacco. Int. J. Mol. Sci. 2020, 21, 1323. [Google Scholar] [CrossRef]
- Dave, A.; Sanadhya, P.; Joshi, P.S.; Agarwal, P.; Agarwal, P.K. Molecular cloning and characterization of high-affinity potassium transporter (AlHKT2;1) gene promoter from halophyte Aeluropus lagopoides. Int. J. Biol. Macromol. 2021, 181, 1254–1264. [Google Scholar] [CrossRef]
- Sun, Y.; Xu, W.; Jia, Y.; Wang, M.; Xia, G. The wheat TaGBF1 gene is involved in the blue-light response and salt tolerance. Plant J. 2015, 84, 1219–1230. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- Lalitha, S. Primer Premier 5. Biotech Softw. Internet Rep. 2000, 1, 270–272. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Trinidad, J.L.; Longkumer, T.; Kohli, A. Rice Protoplast Isolation and Transfection for Transient Gene Expression Analysis. Methods Mol. Biol. 2021, 2238, 313–324. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Atkinson, R.; Janssen, B. The choice of agrobacterium strain for transformation of kiwifruit. Acta Hortic. 2007, 753, 227–232. [Google Scholar] [CrossRef]
- Abdelbaki, A.; Schlerf, M.; Retzlaff, R.; Machwitz, M.; Verrelst, J.; Udelhoven, T. Comparison of Crop Trait Retrieval Strategies Using UAV-Based VNIR Hyperspectral Imaging. Remote Sens. 2021, 13, 1748. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cis-Acting Element | Sequence | Function Annotation |
|---|---|---|
| ABRE | CACGTG | Abscisic acid responsiveness |
| ARE | AAACCA | Anaerobic induction |
| Box 4 | ATTAAT | Light responsiveness |
| CGTCA-motif | CGTCA | MeJA-responsiveness |
| G-Box | CACGTG | Light responsiveness |
| MYB | CAACAG | MYB binding site |
| P-box | CCTTTTG | Gibberellin-responsive element |
| TGA-element | AACGAC | Auxin-responsive element |
| TGACG-motif | TGACG | MeJA-responsiveness |
| Protein | UniProt Accession |
|---|---|
| AtHKT1 | Q84TI7.1 |
| CsHKT6 | A0A067FEV5 |
| NtHKT1 | A0A1S3YZS4 |
| NtHKT1;2 | A0A1S3Z6K8 |
| OsHKT1;1 | Q7XPF8 |
| OsHKT1;3 | Q6H501 |
| OsHKT1;4 | Q7XPF7 |
| OsHKT1;5 | Q0JNB6 |
| OsHKT2;1 | Q0D9S3 |
| OsHKT2;2 | Q93XI5.1 |
| OsHKT2;3 | Q8L481 |
| OsHKT2;4 | Q8L4K5 |
| PpHKT1 | A0A251QTK2 |
| TaHKT2;1 | AAA52749 |
| SlHKT1 | A0A3Q7H8N9 |
| TsHKT1;2 | BAJ34563 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gu, S.; Han, S.; Abid, M.; Bai, D.; Lin, M.; Sun, L.; Qi, X.; Zhong, Y.; Fang, J. A High-K+ Affinity Transporter (HKT) from Actinidia valvata Is Involved in Salt Tolerance in Kiwifruit. Int. J. Mol. Sci. 2023, 24, 15737. https://doi.org/10.3390/ijms242115737
Gu S, Han S, Abid M, Bai D, Lin M, Sun L, Qi X, Zhong Y, Fang J. A High-K+ Affinity Transporter (HKT) from Actinidia valvata Is Involved in Salt Tolerance in Kiwifruit. International Journal of Molecular Sciences. 2023; 24(21):15737. https://doi.org/10.3390/ijms242115737
Chicago/Turabian StyleGu, Shichao, Shiming Han, Muhammad Abid, Danfeng Bai, Miaomiao Lin, Leiming Sun, Xiujuan Qi, Yunpeng Zhong, and Jinbao Fang. 2023. "A High-K+ Affinity Transporter (HKT) from Actinidia valvata Is Involved in Salt Tolerance in Kiwifruit" International Journal of Molecular Sciences 24, no. 21: 15737. https://doi.org/10.3390/ijms242115737
APA StyleGu, S., Han, S., Abid, M., Bai, D., Lin, M., Sun, L., Qi, X., Zhong, Y., & Fang, J. (2023). A High-K+ Affinity Transporter (HKT) from Actinidia valvata Is Involved in Salt Tolerance in Kiwifruit. International Journal of Molecular Sciences, 24(21), 15737. https://doi.org/10.3390/ijms242115737