
Evaluation of Local Retinal Function in Light-Damaged Rats Using Multifocal Electroretinograms and Multifocal Visual Evoked Potentials
 , and
, and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
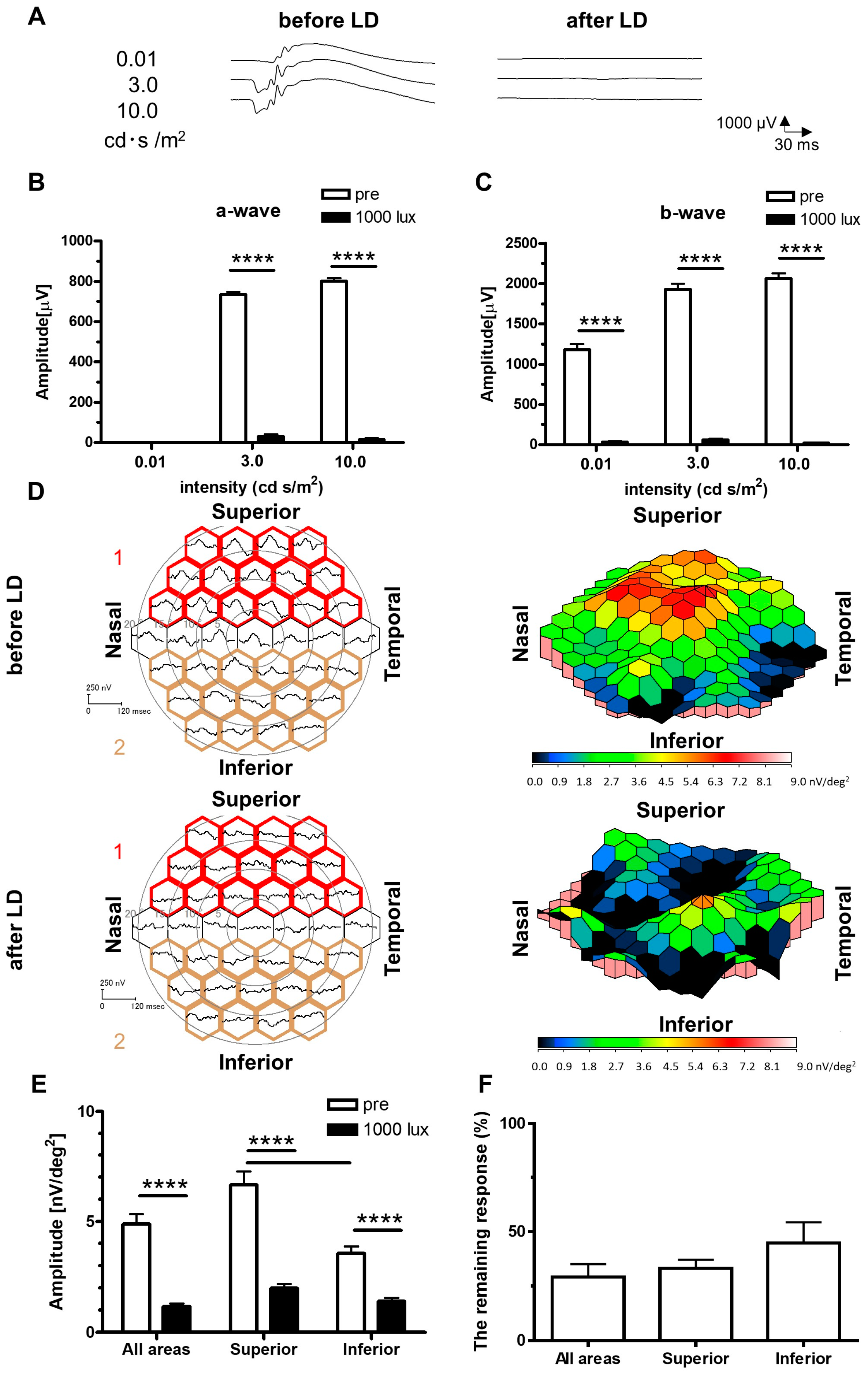
2. Results
2.1. Recordings of ERGs and mfERGs on the LD Rats
2.2. Recordings of VEPs and mfVEPs on the LD Rats
2.3. Histological Evaluation of the LD Model
2.4. Correlation of Electrophysiological and Histological Evaluations
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Induction of Light Damage
4.3. Recording of ERGs and VEPs
4.4. Recording of mfERGs and mfVEPs
4.5. Histological Studies of the Retinas with HE Staining
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hartong, D.T.; Berson, E.L.; Dryja, T.P. Retinitis pigmentosa. Lancet 2006, 368, 1795–1809. [Google Scholar] [CrossRef]
- Lim, L.S.; Mitchell, P.; Seddon, J.M.; Holz, F.G.; Wong, T.Y. Age-related macular degeneration. Lancet 2012, 379, 1728–1738. [Google Scholar] [CrossRef]
- Furukawa, A.; Koriyama, Y. A role of heat shock protein 70 in photoreceptor cell death: Potential as a novel therapeutic target in retinal degeneration. CNS Neurosci. Ther. 2016, 22, 7–14. [Google Scholar] [CrossRef]
- Maurya, M.; Bora, K.; Blomfield, A.; Pavlovich, M.; Huang, S.; Liu, C.H.; Chen, J. Oxidative stress in retinal pigment epithelium degeneration: From pathogenesis to therapeutic targets in dry age-related macular degeneration. Neural Regen. Res. 2023, 18, 2173. [Google Scholar]
- Mody, S.; Joshi, A. Age-related macular degeneration and its association with neurodegenerative disorders. Cureus 2023, 15, e34920. [Google Scholar] [CrossRef]
- Momozawa, Y.; Akiyama, M.; Kamatani, Y.; Arakawa, S.; Yasuda, M.; Yoshida, S.; Oshima, Y.; Mori, R.; Tanaka, K.; Mori, K.; et al. Low-frequency coding variants in CETP and CFB are associated with susceptibility of exudative age-related macular degeneration in the Japanese population. Hum. Mol. Genet. 2016, 25, 5027–5034. [Google Scholar]
- Allikmets, R.; Shroyer, N.F.; Singh, N.; Seddon, J.M.; Lewis, R.A.; Bernstein, P.S.; Peiffer, A.; Zabriskie, N.A.; Li, Y.; Hutchinson, A.; et al. Mutation of the stargardt disease gene (ABCR) in age-related macular degeneration. Science 1997, 277, 1805–1807. [Google Scholar] [CrossRef]
- Hogan, M.J. Role of the retinal pigment epithelium in macular disease. Trans. Am. Acad. Ophthalmol. Otolaryngol. 1972, 76, 64–80. [Google Scholar]
- Young, R.W. Pathophysiology of age-related macular degeneration. Surv. Ophthalmol. 1987, 31, 291–306. [Google Scholar] [CrossRef]
- Congdon, N.; O’Colmain, B.; Klaver, C.C.W.; Klein, R.; Muñoz, B.; Friedman, D.S.; Kempen, J.; Taylor, H.R.; Mitchell, P. Eye Diseases Prevalence Research Group. Causes and prevalence of visual impairment among adults in the united states. Arch. Ophthalmol. 2004, 122, 477–485. [Google Scholar]
- Wong, W.L.; Su, X.; Li, X.; Cheung, C.M.G.; Klein, R.; Cheng, C.Y.; Wong, T.Y. Global prevalence of age-related macular degeneration and disease burden projection for 2020 and 2040: A systematic review and meta-analysis. Lancet Glob. Health 2014, 2, e106–e116. [Google Scholar] [CrossRef]
- Kiuchi, K.; Yoshizawa, K.; Shikata, N.; Matsumura, M.; Tsubura, A. Nicotinamide prevents N-methyl-N-nitrosourea-induced photoreceptor cell apoptosis in Sprague-Dawley rats and C57BL mice. Exp. Eye Res. 2002, 74, 383–392. [Google Scholar] [CrossRef]
- Gao, Y.; Deng, X.G.; Sun, Q.N.; Zhong, Z.Q. Ganoderma spore lipid inhibits N-methyl-N-nitrosourea-induced retinal photoreceptor apoptosis in vivo. Exp. Eye Res. 2010, 90, 397–404. [Google Scholar] [CrossRef]
- Sugano, E.; Tabata, K.; Takezawa, T.; Shiraiwa, R.; Muraoka, H.; Metoki, T.; Kudo, A.; Iwama, Y.; Nakazawa, M.; Tomita, H. N-Methyl-N-Nitrosourea-Induced photoreceptor degeneration is inhibited by nicotinamide via the blockade of upstream events before the phosphorylation of signalling proteins. Biomed. Res. Int. 2019, 2019, 3238719. [Google Scholar] [CrossRef]
- Li, G.; Anderson, R.E.; Tomita, H.; Adler, R.; Liu, X.; Zack, D.J.; Rajala, R.V.S. Nonredundant role of Akt2 for neuroprotection of rod photoreceptor cells from light-induced cell death. J. Neurosci. 2007, 27, 203–211. [Google Scholar] [CrossRef]
- Tomita, H.; Kotake, Y.; Anderson, R.E. Mechanism of protection from light-induced retinal degeneration by the synthetic antioxidant phenyl-N-tert-butylnitrone. Investig. Ophthalmol. Vis. Sci. 2005, 46, 427–434. [Google Scholar] [CrossRef]
- Kikuchi, Y.; Sugano, E.; Yuki, S.; Tabata, K.; Endo, Y.; Takita, Y.; Onoguchi, R.; Ozaki, T.; Fukuda, T.; Takai, Y.; et al. SIG-1451, a novel, non-steroidal anti-inflammatory compound, attenuates light-induced photoreceptor degeneration by affecting the inflammatory process. Int. J. Mol. Sci. 2022, 23, 8802. [Google Scholar] [CrossRef]
- Tabata, K.; Sugano, E.; Hatakeyama, A.; Watanabe, Y.; Suzuki, T.; Ozaki, T.; Fukuda, T.; Tomita, H. Phototoxicities caused by continuous light exposure were not induced in retinal ganglion cells transduced by an optogenetic gene. Int. J. Mol. Sci. 2021, 22, 6732. [Google Scholar] [CrossRef]
- Mullen, R.J.; LaVail, M.M. Inherited retinal dystrophy: Primary defect in pigment epithelium determined with experimental rat chimeras. Science 1976, 192, 799–801. [Google Scholar] [CrossRef]
- Tomita, H.; Sugano, E.; Murayama, N.; Ozaki, T.; Nishiyama, F.; Tabata, K.; Takahashi, M.; Saito, T.; Tamai, M. Restoration of the majority of the visual spectrum by using modified Volvox channelrhodopsin-1. Mol. Ther. 2014, 22, 1434–1440. [Google Scholar] [CrossRef]
- Sugano, E.; Tabata, K.; Takahashi, M.; Nishiyama, F.; Shimizu, H.; Sato, M.; Tamai, M.; Tomita, H. Local and systemic responses following intravitreous injection of AAV2-encoded modified Volvox channelrhodopsin-1 in a genetically blind rat model. Gene Ther. 2016, 23, 158–166. [Google Scholar] [CrossRef]
- LaVail, M.M.; Nishikawa, S.; Steinberg, R.H.; Naash, M.I.; Duncan, J.L.; Trautmann, N.; Matthes, M.T.; Yasumura, D.; Lau-Villacorta, C.; Chen, J.; et al. Phenotypic characterization of P23H and S334ter rhodopsin transgenic rat models of inherited retinal degeneration. Exp. Eye Res. 2018, 167, 56–90. [Google Scholar] [CrossRef] [PubMed]
- Deans, M.R.; Volgyi, B.; Goodenough, D.A.; Bloomfield, S.A.; Paul, D.L. Connexin36 is essential for transmission of rod-mediated visual signals in the mammalian retina. Neuron 2002, 36, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Kominami, T.; Ueno, S.; Okado, S.; Nakanishi, A.; Kondo, M.; Terasaki, H. Contributions of second- and third-order retinal neurons to cone electroretinograms after loss of rod function in rhodopsin P347L transgenic rabbits. Investig. Ophthalmol. Vis. Sci. 2017, 58, 1417–1424. [Google Scholar] [CrossRef] [PubMed]
- Ranchon, I.; Chen, S.; Alvarez, K.; Anderson, R.E. Systemic administration of phenyl-N-tert-butylnitrone protects the retina from light damage. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1375–1379. [Google Scholar]
- Schatz, A.; Arango-Gonzalez, B.; Fischer, D.; Enderle, H.; Bolz, S.; Röck, T.; Naycheva, L.; Grimm, C.; Messias, A.; Zrenner, E.; et al. Transcorneal electrical stimulation shows neuroprotective effects in retinas of light-exposed rats. Investig. Ophthalmol. Vis. Sci. 2012, 53, 5552–5561. [Google Scholar] [CrossRef]
- Noell, W.K.; Walker, V.S.; Kang, B.S.; Berman, S. Retinal damage by light in rats. Investig. Ophthalmol. 1966, 5, 450–473. [Google Scholar]
- Robson, A.G.; Frishman, L.J.; Grigg, J.; Hamilton, R.; Jeffrey, B.G.; Kondo, M.; Li, S.; McCulloch, D.L. ISCEV Standard for full-field clinical electroretinography (2022 update). Doc. Ophthalmol. 2022, 144, 165–177. [Google Scholar] [CrossRef]
- Hoffmann, M.B.; Bach, M.; Kondo, M.; Li, S.; Walker, S.; Holopigian, K.; Viswanathan, S.; Robson, A.G. ISCEV standard for clinical multifocal electroretinography (mfERG) (2021 update). Doc. Ophthalmol. 2021, 142, 5–16. [Google Scholar] [CrossRef] [PubMed]
- Ball, S.L.; Petry, H.M. Noninvasive assessment of retinal function in rats using multifocal electroretinography. Investig. Ophthalmol. Vis. Sci. 2000, 41, 610–617. [Google Scholar]
- Chu, P.H.W.; Li, H.Y.; Chin, M.P.; So, K.F.; Chan, H.H.L. Effect of Lycium Barbarum (wolfberry) polysaccharides on preserving retinal function after partial optic nerve transection. PLoS ONE 2013, 8, e81339. [Google Scholar] [CrossRef]
- Odom, J.V.; Bach, M.; Brigell, M.; Holder, G.E.; McCulloch, D.L.; Mizota, A.; Tormene, A.P. ISCEV standard for clinical visual evoked potentials: (2016 update). Doc. Ophthalmol. 2016, 133, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lozano, D.C.; Twa, M.D. Development of a rat schematic eye from in vivo biometry and the correction of lateral magnification in sd-oct imaging. Investig. Ophthalmol. Vis. Sci. 2013, 54, 6446–6455. [Google Scholar] [CrossRef] [PubMed]
- Stone, J. Mechanisms of photoreceptor death and survival in mammalian retina. Prog. Retin. Eye Res. 1999, 18, 689–735. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Cao, W.; Anderson, R.E. Alleviation of constant-light–induced photoreceptor degeneration by adaptation of adult albino rat to bright cyclic light. Investig. Ophthalmol. Vis. Sci. 2003, 44, 4968–4975. [Google Scholar] [CrossRef]
- Wen, R.; Song, Y.; Cheng, T.; Matthes, M.; Yasumura, D.; LaVail, M.; Steinberg, R. Injury-induced upregulation of bFGF and CNTF mRNAs in the rat retina. J. Neurosci. 1995, 15, 7377–7385. [Google Scholar] [CrossRef]
- Wen, R.; Cheng, T.; Li, Y.; Cao, W.; Steinberg, R.H. α 2-adrenergic agonists induce basic fibroblast growth factor expression in photoreceptors In Vivo and ameliorate light damage. J. Neurosci. 1996, 16, 5986–5992. [Google Scholar] [CrossRef]
- Liu, C.; Peng, M.; Laties, A.M.; Wen, R. Preconditioning with bright light evokes a protective response against light damage in the rat retina. J. Neurosci. 1998, 18, 1337–1344. [Google Scholar] [CrossRef]
- LaVail, M.M.; Unoki, K.; Yasumura, D.; Matthes, M.T.; Yancopoulos, G.D.; Steinberg, R.H. Multiple growth factors, cytokines, and neurotrophins rescue photoreceptors from the damaging effects of constant light. Proc. Natl. Acad. Sci. USA 1992, 89, 11249–11253. [Google Scholar] [CrossRef]
- Cao, W.; Tombran-Tink, J.; Elias, R.; Sezate, S.; Mrazek, D.; McGinnis, J.F. In vivo protection of photoreceptors from light damage by pigment epithelium-derived factor. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1646–1652. [Google Scholar]
- Ortín-Martínez, A.; Valiente-Soriano, F.J.; García-Ayuso, D.; Alarcón-Martínez, L.; Jiménez-López, M.; Bernal-Garro, J.M.; Nieto-López, L.; Nadal-Nicolás, F.M.; Villegas-Pérez, M.P.; Wheeler, L.A.; et al. A novel in vivo model of focal light emitting diode-induced cone-photoreceptor phototoxicity: Neuroprotection afforded by Brimonidine, BDNF, PEDF or bFGF. PLoS ONE 2014, 9, e113798. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.; Hollyfield, J.G. Basic fibroblast growth factor: Increased gene expression in inherited and light-induced photoreceptor degeneration. Exp. Eye Res. 1996, 62, 181–190. [Google Scholar] [CrossRef] [PubMed]
- O’Driscoll, C.; O’Connor, J.; O’Brien, C.J.; Cotter, T.G. Basic fibroblast growth factor-induced protection from light damage in the mouse retina in vivo. J. Neurochem. 2008, 105, 524–536. [Google Scholar] [CrossRef] [PubMed]
- Hubel, D.H.; Wiesel, T.N. Receptive fields of single neurones in the cat’s striate cortex. J. Physiol. 1959, 148, 574–591. [Google Scholar] [CrossRef]
- Bi, A.; Cui, J.; Ma, Y.P.; Olshevskaya, E.; Pu, M.; Dizhoor, A.M.; Pan, Z.H. Ectopic expression of a microbial-type rhodopsin restores visual responses in mice with photoreceptor degeneration. Neuron 2006, 50, 23–33. [Google Scholar] [CrossRef]
- Tomita, H.; Sugano, E.; Yawo, H.; Ishizuka, T.; Isago, H.; Narikawa, S.; Kügler, S.; Tamai, M. Restoration of visual response in aged dystrophic rcs rats using AAV-mediated channelopsin-2 gene transfer. Investig. Ophthalmol. Vis. Sci. 2007, 48, 3821–3826. [Google Scholar] [CrossRef]
- Sahel, J.A.; Boulanger-Scemama, E.; Pagot, C.; Arleo, A.; Galluppi, F.; Martel, J.N.; Esposti, S.D.; Delaux, A.; de Saint Aubert, J.B.; de Montleau, C.; et al. Partial recovery of visual function in a blind patient after optogenetic therapy. Nat. Med. 2021, 27, 1223–1229. [Google Scholar] [CrossRef]
- Hatakeyama, A.; Sugano, E.; Sayama, T.; Watanabe, Y.; Suzuki, T.; Tabata, K.; Endo, Y.; Sakajiri, T.; Fukuda, T.; Ozaki, T.; et al. Properties of a single amino acid residue in the third transmembrane domain determine the kinetics of ambient light-sensitive channelrhodopsin. Int. J. Mol. Sci. 2023, 24, 5054. [Google Scholar] [CrossRef]
- Watanabe, Y.; Sugano, E.; Tabata, K.; Hatakeyama, A.; Sakajiri, T.; Fukuda, T.; Ozaki, T.; Suzuki, T.; Sayama, T.; Tomita, H. Development of an optogenetic gene sensitive to daylight and its implications in vision restoration. NPJ Regen. Med. 2021, 6, 64. [Google Scholar] [CrossRef]
- Wright, W.; Gajjeraman, S.; Batabyal, S.; Pradhan, S.; Bhattacharya, S.; Mahapatra, V.; Tripathy, A.; Mohanty, S. Restoring vision in mice with retinal degeneration using multicharacteristic opsin. Neurophotonics 2017, 4, 041505. [Google Scholar] [CrossRef]
- Lu, Q.; Ganjawala, T.H.; Krstevski, A.; Abrams, G.W.; Pan, Z.H. Comparison of AAV-mediated optogenetic vision restoration between retinal ganglion cell expression and on bipolar cell targeting. Mol. Ther. Methods Clin. Dev. 2020, 18, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Busskamp, V.; Duebel, J.; Balya, D.; Fradot, M.; Viney, T.J.; Siegert, S.; Groner, A.C.; Cabuy, E.; Forster, V.; Seeliger, M.; et al. Genetic reactivation of cone photoreceptors restores visual responses in retinitis pigmentosa. Science 2010, 329, 413–417. [Google Scholar] [CrossRef] [PubMed]
- Maurya, S.; Mary, B.; Jayandharan, G.R. Rational engineering and preclinical evaluation of neddylation and sumoylation site modified adeno-associated virus vectors in murine models of hemophilia b and leber congenital amaurosis. Hum. Gene Ther. 2019, 30, 1461–1476. [Google Scholar] [CrossRef] [PubMed]
- Da Costa, R.; Röger, C.; Segelken, J.; Barben, M.; Grimm, C.; Neidhardt, J. A novel method combining vitreous aspiration and intravitreal AAV2/8 injection results in retina-wide transduction in adult mice. Investig. Ophthalmol. Vis. Sci. 2016, 57, 5326–5334. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Fan, N.; Wang, N.; Feng, B.; Yang, M.; Liu, G.; Wang, Y.; Zhu, X.; Kaufman, P.L.; Pang, I.H.; et al. Effects of lentivirus-mediated C3 expression on trabecular meshwork cells and intraocular pressure. Investig. Ophthalmol. Vis. Sci. 2018, 59, 4937–4944. [Google Scholar] [CrossRef] [PubMed]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takita, Y.; Sugano, E.; Kitabayashi, K.; Tabata, K.; Saito, A.; Yokoyama, T.; Onoguchi, R.; Fukuda, T.; Ozaki, T.; Bai, L.; et al. Evaluation of Local Retinal Function in Light-Damaged Rats Using Multifocal Electroretinograms and Multifocal Visual Evoked Potentials. Int. J. Mol. Sci. 2023, 24, 16433. https://doi.org/10.3390/ijms242216433
Takita Y, Sugano E, Kitabayashi K, Tabata K, Saito A, Yokoyama T, Onoguchi R, Fukuda T, Ozaki T, Bai L, et al. Evaluation of Local Retinal Function in Light-Damaged Rats Using Multifocal Electroretinograms and Multifocal Visual Evoked Potentials. International Journal of Molecular Sciences. 2023; 24(22):16433. https://doi.org/10.3390/ijms242216433
Chicago/Turabian StyleTakita, Yuya, Eriko Sugano, Kazuya Kitabayashi, Kitako Tabata, Akito Saito, Takanori Yokoyama, Reina Onoguchi, Tomokazu Fukuda, Taku Ozaki, Lanlan Bai, and et al. 2023. "Evaluation of Local Retinal Function in Light-Damaged Rats Using Multifocal Electroretinograms and Multifocal Visual Evoked Potentials" International Journal of Molecular Sciences 24, no. 22: 16433. https://doi.org/10.3390/ijms242216433