
Personalized Cancer Vaccines Go Viral: Viral Vectors in the Era of Personalized Immunotherapy of Cancer
 , ,
, ,
Abstract
:1. Introduction
2. Tumor Antigens
3. High-Throughput Identification of Neoantigens for Personalized Cancer Vaccines
4. Viral-Vectored Vaccines Targeting Tumor Neoantigens
5. Manufacturing Viral Vector-Based Personalized Cancer Vaccines
6. Clinical Trial Landscape of Personalized Vaccines
7. Conclusions and Future Directions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Finn, O.J. The Dawn of Vaccines for Cancer Prevention. Nat. Rev. Immunol. 2018, 18, 183–194. [Google Scholar] [CrossRef]
- Shemesh, C.S.; Hsu, J.C.; Hosseini, I.; Shen, B.Q.; Rotte, A.; Twomey, P.; Girish, S.; Wu, B. Personalized Cancer Vaccines: Clinical Landscape, Challenges, and Opportunities. Mol. Ther. 2021, 29, 555–570. [Google Scholar] [CrossRef]
- Saxena, M.; van der Burg, S.H.; Melief, C.J.M.; Bhardwaj, N. Therapeutic Cancer Vaccines. Nat. Rev. Cancer. 2021, 21, 360–378. [Google Scholar] [CrossRef]
- Kantoff, P.W.; Higano, C.S.; Shore, N.D.; Berger, E.R.; Small, E.J.; Penson, D.F.; Redfern, C.H.; Ferrari, A.C.; Dreicer, R.; Sims, R.B.; et al. Sipuleucel-T Immunotherapy for Castration-Resistant Prostate Cancer. N. Engl. J. Med. 2010, 363, 411–422. [Google Scholar] [CrossRef]
- van der Burg, S.H. Correlates of Immune and Clinical Activity of Novel Cancer Vaccines. Semin. Immunol. 2018, 39, 119–136. [Google Scholar] [CrossRef]
- Tran, T.; Blanc, C.; Granier, C.; Saldmann, A.; Tanchot, C.; Tartour, E. Therapeutic Cancer Vaccine: Building the Future from Lessons of the Past. Semin. Immunopathol. 2019, 41, 69–85. [Google Scholar] [CrossRef]
- Sahin, U.; Tureci, O. Personalized Vaccines for Cancer Immunotherapy. Science 2018, 359, 1355–1360. [Google Scholar] [CrossRef]
- Peri, A.; Salomon, N.; Wolf, Y.; Kreiter, S.; Diken, M.; Samuels, Y. The Landscape of T Cell Antigens for Cancer Immunotherapy. Nat. Cancer 2023, 4, 937–954. [Google Scholar] [CrossRef]
- Buonaguro, L.; Tagliamonte, M. Selecting Target Antigens for Cancer Vaccine Development. Vaccines 2020, 8, 615. [Google Scholar] [CrossRef]
- Gros, A.; Parkhurst, M.R.; Tran, E.; Pasetto, A.; Robbins, P.F.; Ilyas, S.; Prickett, T.D.; Gartner, J.J.; Crystal, J.S.; Roberts, I.M.; et al. Prospective Identification of Neoantigen-Specific Lymphocytes in the Peripheral Blood of Melanoma Patients. Nat. Med. 2016, 22, 433–438. [Google Scholar] [CrossRef]
- Haen, S.P.; Loffler, M.W.; Rammensee, H.G.; Brossart, P. Towards New Horizons: Characterization, Classification and Implications of the Tumour Antigenic Repertoire. Nat. Rev. Clin. Oncol. 2020, 17, 595–610. [Google Scholar] [CrossRef]
- Klebanoff, C.A.; Wolchok, J.D. Shared Cancer Neoantigens: Making Private Matters Public. J. Exp. Med. 2018, 215, 5–7. [Google Scholar] [CrossRef]
- Rizvi, N.A.; Hellmann, M.D.; Snyder, A.; Kvistborg, P.; Makarov, V.; Havel, J.J.; Lee, W.; Yuan, J.; Wong, P.; Ho, T.S.; et al. Cancer Immunology. Mutational Landscape Determines Sensitivity to Pd-1 Blockade in Non-Small Cell Lung Cancer. Science 2015, 348, 124–128. [Google Scholar] [CrossRef]
- Jardim, D.L.; Goodman, A.; de Melo Gagliato, D.; Kurzrock, R. The Challenges of Tumor Mutational Burden as an Immunotherapy Biomarker. Cancer Cell. 2021, 39, 154–173. [Google Scholar] [CrossRef]
- McGranahan, N.; Furness, A.J.; Rosenthal, R.; Ramskov, S.; Lyngaa, R.; Saini, S.K.; Jamal-Hanjani, M.; Wilson, G.A.; Birkbak, N.J.; Hiley, C.T.; et al. Clonal Neoantigens Elicit T Cell Immunoreactivity and Sensitivity to Immune Checkpoint Blockade. Science 2016, 351, 1463–1469. [Google Scholar] [CrossRef]
- D’Alise, A.M.; Leoni, G.; Cotugno, G.; Troise, F.; Langone, F.; Fichera, I.; De Lucia, M.; Avalle, L.; Vitale, R.; Leuzzi, A.; et al. Adenoviral Vaccine Targeting Multiple Neoantigens as Strategy to Eradicate Large Tumors Combined with Checkpoint Blockade. Nat. Commun. 2019, 10, 2688. [Google Scholar] [CrossRef]
- Lee, C.H.; Yelensky, R.; Jooss, K.; Chan, T.A. Update on Tumor Neoantigens and Their Utility: Why It Is Good to Be Different. Trends Immunol. 2018, 39, 536–548. [Google Scholar] [CrossRef]
- Schumacher, T.N.; Scheper, W.; Kvistborg, P. Cancer Neoantigens. Annu Rev. Immunol. 2019, 37, 173–200. [Google Scholar] [CrossRef]
- Roudko, V.; Greenbaum, B.; Bhardwaj, N. Computational Prediction and Validation of Tumor-Associated Neoantigens. Front. Immunol. 2020, 11, 27. [Google Scholar] [CrossRef]
- Blass, E.; Ott, P.A. Advances in the Development of Personalized Neoantigen-Based Therapeutic Cancer Vaccines. Nat. Rev. Clin. Oncol. 2021, 18, 215–229. [Google Scholar] [CrossRef]
- Lang, F.; Schrors, B.; Lower, M.; Tureci, O.; Sahin, U. Identification of Neoantigens for Individualized Therapeutic Cancer Vaccines. Nat. Rev. Drug. Discov. 2022, 21, 261–282. [Google Scholar] [CrossRef]
- Jin, J.; Chen, Z.; Liu, J.; Du, H.; Zhang, G. Towards an Accurate and Robust Analysis Pipeline for Somatic Mutation Calling. Front. Genet. 2022, 13, 979928. [Google Scholar] [CrossRef]
- Kandoth, C.; McLellan, M.D.; Vandin, F.; Ye, K.; Niu, B.; Lu, C.; Xie, M.; Zhang, Q.; McMichael, J.F.; Wyczalkowski, M.A.; et al. Mutational Landscape and Significance across 12 Major Cancer Types. Nature 2013, 502, 333–339. [Google Scholar] [CrossRef]
- De Mattos-Arruda, L.; Vazquez, M.; Finotello, F.; Lepore, R.; Porta, E.; Hundal, J.; Amengual-Rigo, P.; Ng, C.K.Y.; Valencia, A.; Carrillo, J.; et al. Neoantigen Prediction and Computational Perspectives Towards Clinical Benefit: Recommendations from the Esmo Precision Medicine Working Group. Ann. Oncol. 2020, 31, 978–990. [Google Scholar] [CrossRef]
- Singh, N.K.; Riley, T.P.; Baker, S.C.B.; Borrman, T.; Weng, Z.; Baker, B.M. Emerging Concepts in Tcr Specificity: Rationalizing and (Maybe) Predicting Outcomes. J. Immunol. 2017, 199, 2203–2213. [Google Scholar] [CrossRef]
- Luo, J.; Kesselheim, A.S. Insulin Patents and Market Exclusivities: Unresolved Issues—Authors’ Reply. Lancet Diabetes Endocrinol. 2016, 4, 98–99. [Google Scholar] [CrossRef]
- Tappeiner, E.; Finotello, F.; Charoentong, P.; Mayer, C.; Rieder, D.; Trajanoski, Z. Timiner: Ngs Data Mining Pipeline for Cancer Immunology and Immunotherapy. Bioinformatics 2017, 33, 3140–3141. [Google Scholar] [CrossRef]
- Liu, C.; Zhang, Y.; Jian, X.; Tan, X.; Lu, M.; Ouyang, J.; Liu, Z.; Li, Y.; Xu, L.; Chen, L.; et al. Progeo-Neo V2.0: A One-Stop Software for Neoantigen Prediction and Filtering Based on the Proteogenomics Strategy. Genes 2022, 13, 783. [Google Scholar] [CrossRef]
- Rasmussen, M.; Fenoy, E.; Harndahl, M.; Kristensen, A.B.; Nielsen, I.K.; Nielsen, M.; Buus, S. Pan-Specific Prediction of Peptide-Mhc Class I Complex Stability, a Correlate of T Cell Immunogenicity. J. Immunol. 2016, 197, 1517–1524. [Google Scholar] [CrossRef]
- Rubinsteyn, A.; Hodes, I.; Kodysh, J.; Hammerbacher, J. Vaxrank: A Computational Tool for Designing Personalized Cancer Vaccines. Biorxiv 2018, 142919. [Google Scholar]
- Leoni, G.; D’Alise, A.M.; Tucci, F.G.; Micarelli, E.; Garzia, I.; De Lucia, M.; Langone, F.; Nocchi, L.; Cotugno, G.; Bartolomeo, R.; et al. Venus, a Novel Selection Approach to Improve the Accuracy of Neoantigens’ Prediction. Vaccines 2021, 9, 880. [Google Scholar] [CrossRef]
- Luksza, M.; Sethna, Z.M.; Rojas, L.A.; Lihm, J.; Bravi, B.; Elhanati, Y.; Soares, K.; Amisaki, M.; Dobrin, A.; Hoyos, D.; et al. Neoantigen Quality Predicts Immunoediting in Survivors of Pancreatic Cancer. Nature 2022, 606, 389–395. [Google Scholar] [CrossRef]
- Tsang, K.Y.; Zaremba, S.; Nieroda, C.A.; Zhu, M.Z.; Hamilton, J.M.; Schlom, J. Generation of Human Cytotoxic T Cells Specific for Human Carcinoembryonic Antigen Epitopes from Patients Immunized with Recombinant Vaccinia-Cea Vaccine. J. Natl. Cancer Inst. 1995, 87, 982–990. [Google Scholar] [CrossRef]
- Kass, E.; Schlom, J.; Thompson, J.; Guadagni, F.; Graziano, P.; Greiner, J.W. Induction of Protective Host Immunity to Carcinoembryonic Antigen (Cea), a Self-Antigen in Cea Transgenic Mice, by Immunizing with a Recombinant Vaccinia-Cea Virus. Cancer Res. 1999, 59, 676–683. [Google Scholar]
- Kantor, J.; Irvine, K.; Abrams, S.; Kaufman, H.; DiPietro, J.; Schlom, J. Antitumor Activity and Immune Responses Induced by a Recombinant Carcinoembryonic Antigen-Vaccinia Virus Vaccine. J. Natl. Cancer Inst. 1992, 84, 1084–1091. [Google Scholar] [CrossRef]
- Holmgren, J.; Czerkinsky, C. Mucosal Immunity and Vaccines. Nat. Med. 2005, 11, S45–S53. [Google Scholar] [CrossRef]
- Tang, Y.; Le, L.P.; Matthews, Q.L.; Han, T.; Wu, H.; Curiel, D.T. Derivation of a Triple Mosaic Adenovirus Based on Modification of the Minor Capsid Protein Ix. Virology 2008, 377, 391–400. [Google Scholar] [CrossRef]
- Sasso, E.; D’Alise, A.M.; Zambrano, N.; Scarselli, E.; Folgori, A.; Nicosia, A. New Viral Vectors for Infectious Diseases and Cancer. Semin. Immunol. 2020, 50, 101430. [Google Scholar] [CrossRef]
- Mennechet, F.J.D.; Paris, O.; Ouoba, A.R.; Arenas, S.S.; Sirima, S.B.; Dzomo, G.R.T.; Diarra, A.; Traore, I.T.; Kania, D.; Eichholz, K.; et al. A Review of 65 Years of Human Adenovirus Seroprevalence. Expert Rev. Vaccines 2019, 18, 597–613. [Google Scholar] [CrossRef]
- Palmer, C.D.; Rappaport, A.R.; Davis, M.J.; Hart, M.G.; Scallan, C.D.; Hong, S.J.; Gitlin, L.; Kraemer, L.D.; Kounlavouth, S.; Yang, A.; et al. Individualized, Heterologous Chimpanzee Adenovirus and Self-Amplifying Mrna Neoantigen Vaccine for Advanced Metastatic Solid Tumors: Phase 1 Trial Interim Results. Nat. Med. 2022, 28, 1619–1629. [Google Scholar] [CrossRef]
- Bechter, O.; D’Alise, A.M.; Leoni, G.; Cotugno, G.; Siani, L.; Vitale, R.; Ruzza, V.; Garzia, I.; Antonucci, L.; Micarelli, E.; et al. Abstract Lb196: Nous-Pev, a Personalized Cancer Immunotherapy Targeting Neoantigens, Induces Long Lasting, Tumor Infiltrating Memory T Cells. Cancer Res. 2023, 83, LB196. [Google Scholar] [CrossRef]
- D’Alise, A.M.; Brasu, N.; De Intinis, C.; Leoni, G.; Russo, V.; Langone, F.; Baev, D.; Micarelli, E.; Petiti, L.; Picelli, S.; et al. Adenoviral-Based Vaccine Promotes Neoantigen-Specific Cd8(+) T Cell Stemness and Tumor Rejection. Sci. Transl. Med. 2022, 14, eabo7604. [Google Scholar] [CrossRef]
- Carroll, M.W.; Moss, B. Host Range and Cytopathogenicity of the Highly Attenuated Mva Strain of Vaccinia Virus: Propagation and Generation of Recombinant Viruses in a Nonhuman Mammalian Cell Line. Virology 1997, 238, 198–211. [Google Scholar] [CrossRef]
- Sutter, G.; Staib, C. Vaccinia Vectors as Candidate Vaccines: The Development of Modified Vaccinia Virus Ankara for Antigen Delivery. Curr. Drug. Targets Infect. Disord. 2003, 3, 263–271. [Google Scholar] [CrossRef]
- Goepfert, P.A.; Elizaga, M.L.; Sato, A.; Qin, L.; Cardinali, M.; Hay, C.M.; Hural, J.; DeRosa, S.C.; DeFawe, O.D.; Tomaras, G.D.; et al. Phase 1 Safety and Immunogenicity Testing of DNA and Recombinant Modified Vaccinia Ankara Vaccines Expressing Hiv-1 Virus-Like Particles. J. Infect. Dis. 2011, 203, 610–619. [Google Scholar] [CrossRef]
- McCurdy, L.H.; Larkin, B.D.; Martin, J.E.; Graham, B.S. Modified Vaccinia Ankara: Potential as an Alternative Smallpox Vaccine. Clin. Infect. Dis. 2004, 38, 1749–1753. [Google Scholar] [CrossRef]
- Swadling, L.; Capone, S.; Antrobus, R.D.; Brown, A.; Richardson, R.; Newell, E.W.; Halliday, J.; Kelly, C.; Bowen, D.; Fergusson, J.; et al. A Human Vaccine Strategy Based on Chimpanzee Adenoviral and Mva Vectors That Primes, Boosts, and Sustains Functional Hcv-Specific T Cell Memory. Sci. Transl. Med. 2014, 6, 261ra153. [Google Scholar] [CrossRef]
- Vollmar, J.; Arndtz, N.; Eckl, K.M.; Thomsen, T.; Petzold, B.; Mateo, L.; Schlereth, B.; Handley, A.; King, L.; Hulsemann, V.; et al. Safety and Immunogenicity of Imvamune, a Promising Candidate as a Third Generation Smallpox Vaccine. Vaccine 2006, 24, 2065–2070. [Google Scholar] [CrossRef]
- Ye, T.; Li, F.; Ma, G.; Wei, W. Enhancing Therapeutic Performance of Personalized Cancer Vaccine Via Delivery Vectors. Adv. Drug Deliv. Rev. 2021, 177, 113927. [Google Scholar] [CrossRef]
- Cappuccini, F.; Bryant, R.; Pollock, E.; Carter, L.; Verrill, C.; Hollidge, J.; Poulton, I.; Baker, M.; Mitton, C.; Baines, A.; et al. Safety and Immunogenicity of Novel 5t4 Viral Vectored Vaccination Regimens in Early Stage Prostate Cancer: A Phase I Clinical Trial. J. Immunother. Cancer 2020, 8, e000928. [Google Scholar] [CrossRef]
- Leoni, G.; D’Alise, A.M.; Cotugno, G.; Langone, F.; Garzia, I.; De Lucia, M.; Fichera, I.; Vitale, R.; Bignone, V.; Tucci, F.G.; et al. A Genetic Vaccine Encoding Shared Cancer Neoantigens to Treat Tumors with Microsatellite Instability. Cancer Res. 2020, 80, 3972–3982. [Google Scholar] [CrossRef]
- Li, X.; Le, Y.; Zhang, Z.; Nian, X.; Liu, B.; Yang, X. Viral Vector-Based Gene Therapy. Int. J. Mol. Sci. 2023, 24, 7736. [Google Scholar] [CrossRef]
- EMA. Guidelines on Good Manufacturing Practice Specific to Advanced Therapy Medicinal Products; European Medicines Agency: Amsterdam, The Netherlands, 2017; Volume 4, p. 82. [Google Scholar]
- Perrinjaquet, M.; Schlegel, C.R. Personalized Neoantigen Cancer Vaccines: An Analysis of the Clinical and Commercial Potential of Ongoing Development Programs. Drug Discov. Today 2023, 28, 103773. [Google Scholar] [CrossRef]
- Khang, M.; Suryaprakash, S.; Kotrappa, M.; Mulyasasmita, W.; Topp, S.; Wu, J. Manufacturing Innovation to Drive Down Cell Therapy Costs. Trends Biotechnol. 2023, 41, 1216–1219. [Google Scholar] [CrossRef]
- Fang, Y.; Mo, F.; Shou, J.; Wang, H.; Luo, K.; Zhang, S.; Han, N.; Li, H.; Ye, S.; Zhou, Z.; et al. A Pan-Cancer Clinical Study of Personalized Neoantigen Vaccine Monotherapy in Treating Patients with Various Types of Advanced Solid Tumors. Clin. Cancer Res. 2020, 26, 4511–4520. [Google Scholar] [CrossRef]
- Chen, Z.; Zhang, S.; Han, N.; Jiang, J.; Xu, Y.; Ma, D.; Lu, L.; Guo, X.; Qiu, M.; Huang, Q.; et al. A Neoantigen-Based Peptide Vaccine for Patients with Advanced Pancreatic Cancer Refractory to Standard Treatment. Front. Immunol. 2021, 12, 691605. [Google Scholar] [CrossRef]
- Shou, J.; Mo, F.; Zhang, S.; Lu, L.; Han, N.; Liu, L.; Qiu, M.; Li, H.; Han, W.; Ma, D.; et al. Combination Treatment of Radiofrequency Ablation and Peptide Neoantigen Vaccination: Promising Modality for Future Cancer Immunotherapy. Front. Immunol. 2022, 13, 1000681. [Google Scholar] [CrossRef]
- Rojas, L.A.; Sethna, Z.; Soares, K.C.; Olcese, C.; Pang, N.; Patterson, E.; Lihm, J.; Ceglia, N.; Guasp, P.; Chu, A.; et al. Personalized Rna Neoantigen Vaccines Stimulate T Cells in Pancreatic Cancer. Nature 2023, 618, 144–150. [Google Scholar] [CrossRef]
- Burris, H.A.; Patel, M.R.; Cho, D.C.; Clarke, J.M.; Gutierrez, M.; Zaks, T.Z.; Frederick, J.; Hopson, K.; Mody, K.; Binanti-Berube, A.; et al. A Phase I Multicenter Study to Assess the Safety, Tolerability, and Immunogenicity of Mrna-4157 Alone in Patients with Resected Solid Tumors and in Combination with Pembrolizumab in Patients with Unresectable Solid Tumors. J. Clin. Oncol. 2019, 37, 2523. [Google Scholar] [CrossRef]
- Carlino, M.S.; Khattak, A.; Weber, J.S.; Meniawy, T.; Sullivan, R.J.; Taylor, M.H.; Kim, K.B.; McKean, M.; Faries, M.B.; Cowey, C.L.; et al. Minimal Residual Disease by Circulating Tumor DNA as a Biomarker of Recurrence Free Survival in Resected High-Risk Melanoma Patients Treated with Mrna-4157/V940, a Personalized Cancer Vaccine, and Pembrolizumab. J. Clin. Oncol. 2023, 41, LBA9515. [Google Scholar] [CrossRef]
- Khattak, A.; Weber, J.S.; Meniawy, T.; Taylor, M.H.; Ansstas, G.; Kim, K.B.; McKean, M.; Long, G.V.; Sullivan, R.J.; Faries, M.B.; et al. Distant Metastasis-Free Survival Results from the Randomized, Phase 2 Mrna-4157-P201/Keynote-942 Trial. J. Clin. Oncol. 2023, 41, LBA9503. [Google Scholar] [CrossRef]
- Dolgin, E. Personalized Cancer Vaccines Pass First Major Clinical Test. Nat. Rev. Drug Discov. 2023, 22, 607–609. [Google Scholar] [CrossRef]
- Tahtinen, S.; Tong, A.J.; Himmels, P.; Oh, J.; Paler-Martinez, A.; Kim, L.; Wichner, S.; Oei, Y.; McCarron, M.J.; Freund, E.C.; et al. Il-1 and Il-1ra Are Key Regulators of the Inflammatory Response to Rna Vaccines. Nat. Immunol. 2022, 23, 532–542. [Google Scholar] [CrossRef]
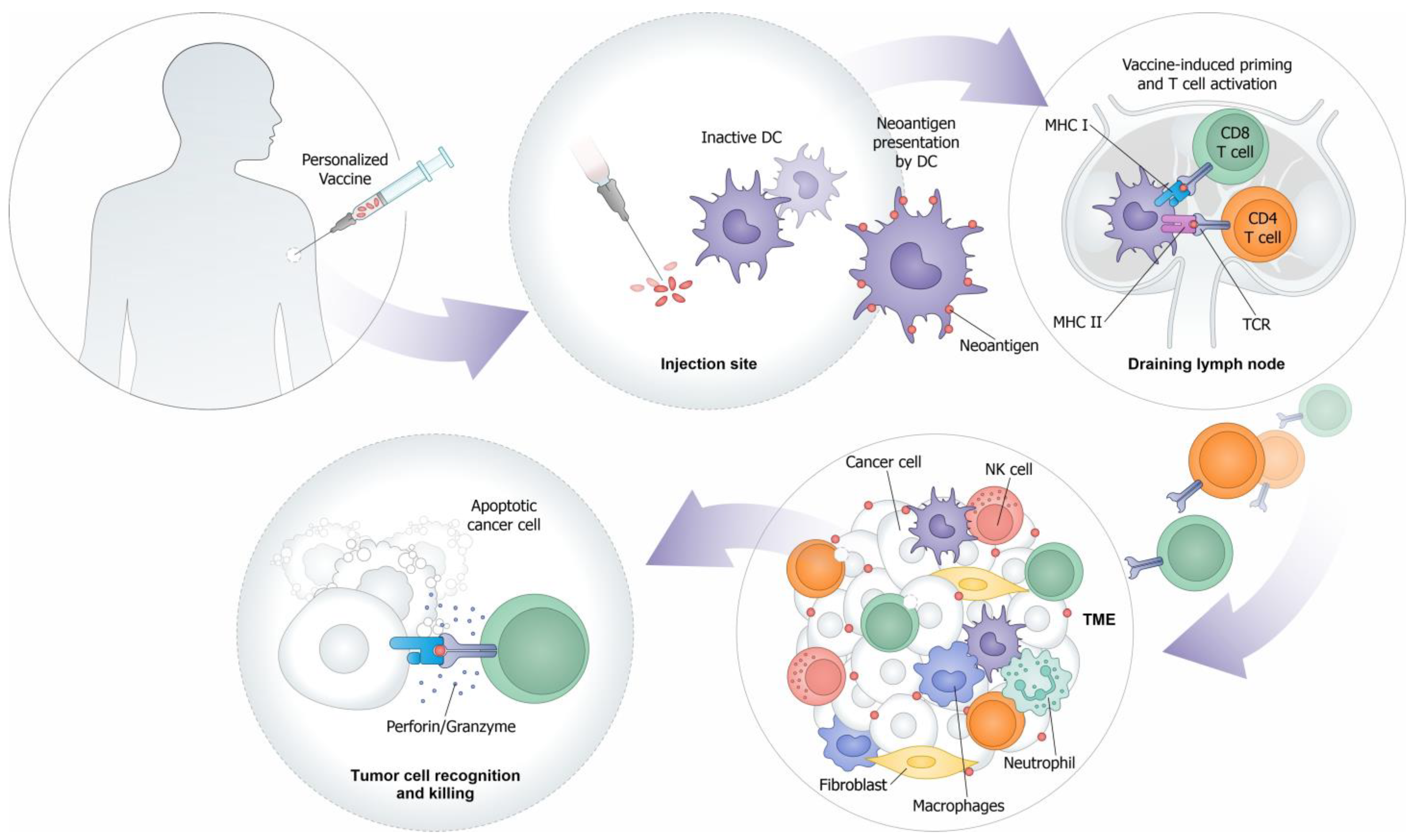
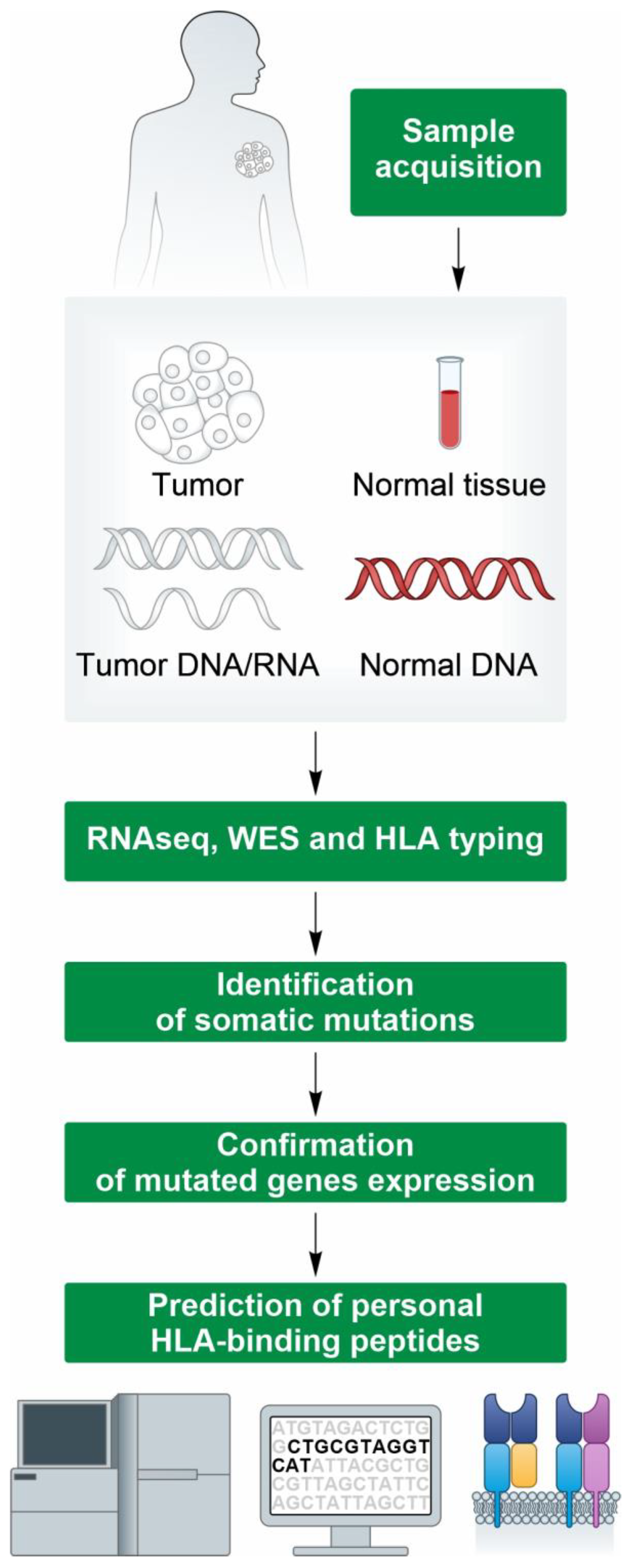
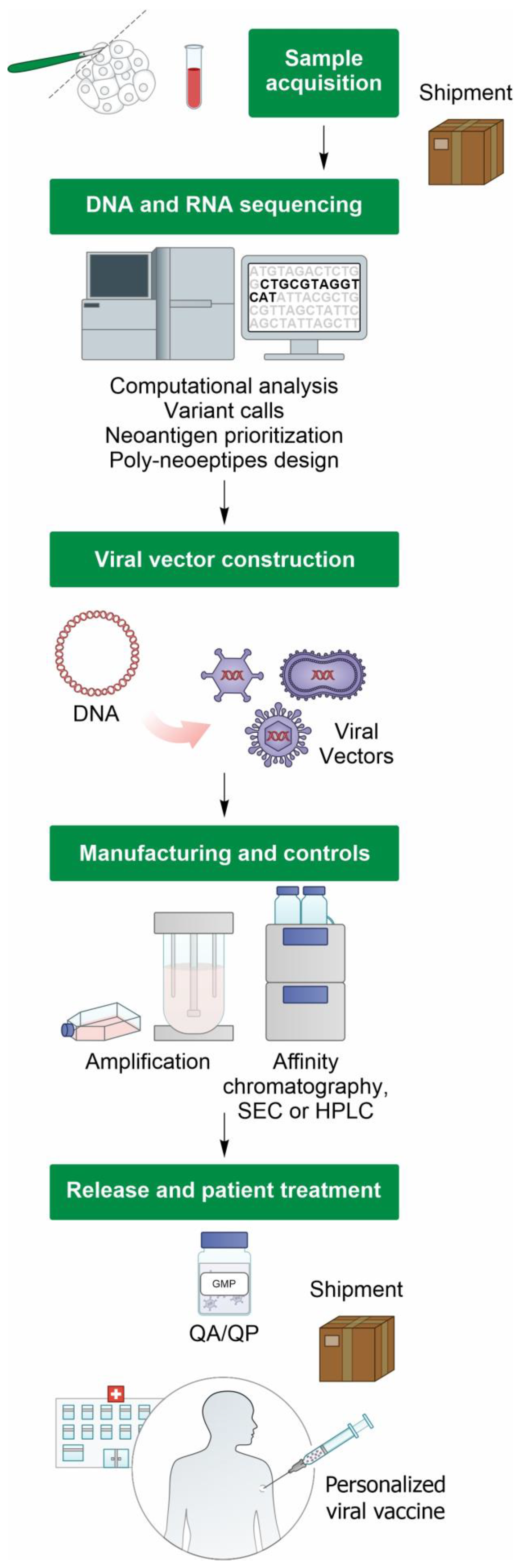

{kind=link}
{kind=link}
{kind=link}
| Vaccine | Company | Platform | Number of Neoantigens | Phase | Main Indication | Adjuvant | Route of Administration | Trial Number | Status | Immunotherapy | Vaccine Doses |
|---|---|---|---|---|---|---|---|---|---|---|---|
| iNEO-Vac-P01 | Hangzhou Neoantigen Therapeutic | Peptide | 5–20 | I | Advanced Pancreatic cancer | GM-CSF | Subcutaneous | NCT03645148 | Completed | aPD1 | 7 |
| Advanced Pancreatic cancer | GM-CSF | NCT03662815 | NA | 7 | |||||||
| Pancreatic cancer | GM-CSF | NCT04810910 | Recruiting | 7 | |||||||
| Esophagus cancer | GM-CSF | NCT05307835 | Recruiting | 7 | |||||||
| EVX-01 | Evaxion Biotech/Merck | Peptide | Up to 10 | II | Metastatic melanoma | Intramuscular | NCT05309421 | Recruiting | aPD1 | 10 | |
| EVX-02 | Evaxion Biotech | DNA | Up to 13 | II | Advanced melanoma | Intramuscular | NCT04455503 | Active, not recruiting | aPD1 | 8 | |
| VB10.NEO | Nykode/Genentech | DNA | Up to 40 | I | Advanced solid tumors (melanoma, NSCLC, RCC, UC, HNSCC, TNBC, gastric/GEJ cancer, cervical, anal, or MSI-high tumors) | Intramuscular | NCT05018273 | Recruiting | aPDL1 | 15 | |
| II | Melanoma, NSCLC, RCC, and SCCHN | Intramuscular | NCT03548467 | Completed | Bempegaldesleukin-NKTR-214 | 14 | |||||
| GNOS-PV02 | Genos Therapeutics | DNA | Up to 40 | II | Metastatic liver cancer | plasmid encoded IL-12 (INO-9012) | Intradermal injection and electroporation | NCT04251117 | Active, not recruiting | aPD1 | NA |
| GRANITE | Gritsone bio | Viral DNA ChAd68 and samRNA | Up to 20 | II | Colon cancer | Intramuscular | NCT05456165 | Terminated | aPDL1 and aCTLA4 | GRT-C901 2 doses and GRT-R902 4 doses | |
| NSCLC, colorectal cancer, gastroesophageal adenocarcinoma, and urothelial carcinoma | Intramuscular | NCT03639714 | Completed | aPD1 and aCTLA4 | GRT-C901 2 doses and GRT-R902 4 doses | ||||||
| III | Metastatic colorectal cancer | Intramuscular | NCT05141721 | Active, not recruiting | aPD1, aCTLA4, aVEGF | GRT-C901 2 doses and GRT-R902 4 doses | |||||
| NOUS-PEV | Nouscom | Viarl DNA GAd and viral DNA MVA | Up to 30 | I | Metastatic NSCLC and melanoma | Intramuscular | NCT04990479 | Active, not recruiting | aPD1 | Gad 1 dose and MVA 3 doses | |
| RO7198457 | BioNTech/Genentech | Unmodified mRNA | Up to 20 | I | Melanoma, NSCLC, bladder cancer, colorectal cancer, TNBC, RCC, and head and neck cancer | Intravenous | NCT03289962 | Active, not recruiting | aPDL1 | at least 8 | |
| I | Pancreatic cancer | Intravenous | NCT04161755 | Active, not recruiting | aPDL1 | 10 | |||||
| II | Colon cancer | Intravenous | NCT04486378 | Recruiting | 15 | ||||||
| III | Advanced melanoma | Intravenous | NCT03815058 | Active, not recruiting | aPD1 | at least 8 | |||||
| mRNA 4157 | Moderna/Merck | Modified mRNA | Up to 34 | I | NSCLC, SCLC, melanoma, bladder urothelial carcinoma, HPV-ve HNSCC, and MSI-high tumors | Intramuscular | NCT03313778 | Active, not recruiting | aPD1 | 9 | |
| II | High-risk melanoma | Intramuscular | NCT03897881 | Recruiting | aPD1 | 9 | |||||
| III | High-risk melanoma | Intramuscular | NCT05933577 | Recruiting | aPD1 | 9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seclì, L.; Leoni, G.; Ruzza, V.; Siani, L.; Cotugno, G.; Scarselli, E.; D’Alise, A.M. Personalized Cancer Vaccines Go Viral: Viral Vectors in the Era of Personalized Immunotherapy of Cancer. Int. J. Mol. Sci. 2023, 24, 16591. https://doi.org/10.3390/ijms242316591
Seclì L, Leoni G, Ruzza V, Siani L, Cotugno G, Scarselli E, D’Alise AM. Personalized Cancer Vaccines Go Viral: Viral Vectors in the Era of Personalized Immunotherapy of Cancer. International Journal of Molecular Sciences. 2023; 24(23):16591. https://doi.org/10.3390/ijms242316591
Chicago/Turabian StyleSeclì, Laura, Guido Leoni, Valentino Ruzza, Loredana Siani, Gabriella Cotugno, Elisa Scarselli, and Anna Morena D’Alise. 2023. "Personalized Cancer Vaccines Go Viral: Viral Vectors in the Era of Personalized Immunotherapy of Cancer" International Journal of Molecular Sciences 24, no. 23: 16591. https://doi.org/10.3390/ijms242316591
APA StyleSeclì, L., Leoni, G., Ruzza, V., Siani, L., Cotugno, G., Scarselli, E., & D’Alise, A. M. (2023). Personalized Cancer Vaccines Go Viral: Viral Vectors in the Era of Personalized Immunotherapy of Cancer. International Journal of Molecular Sciences, 24(23), 16591. https://doi.org/10.3390/ijms242316591