

Acute In Vivo Administration of Compound 21 Stimulates Akt and ERK1/2 Phosphorylation in Mouse Heart and Adipose Tissue
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
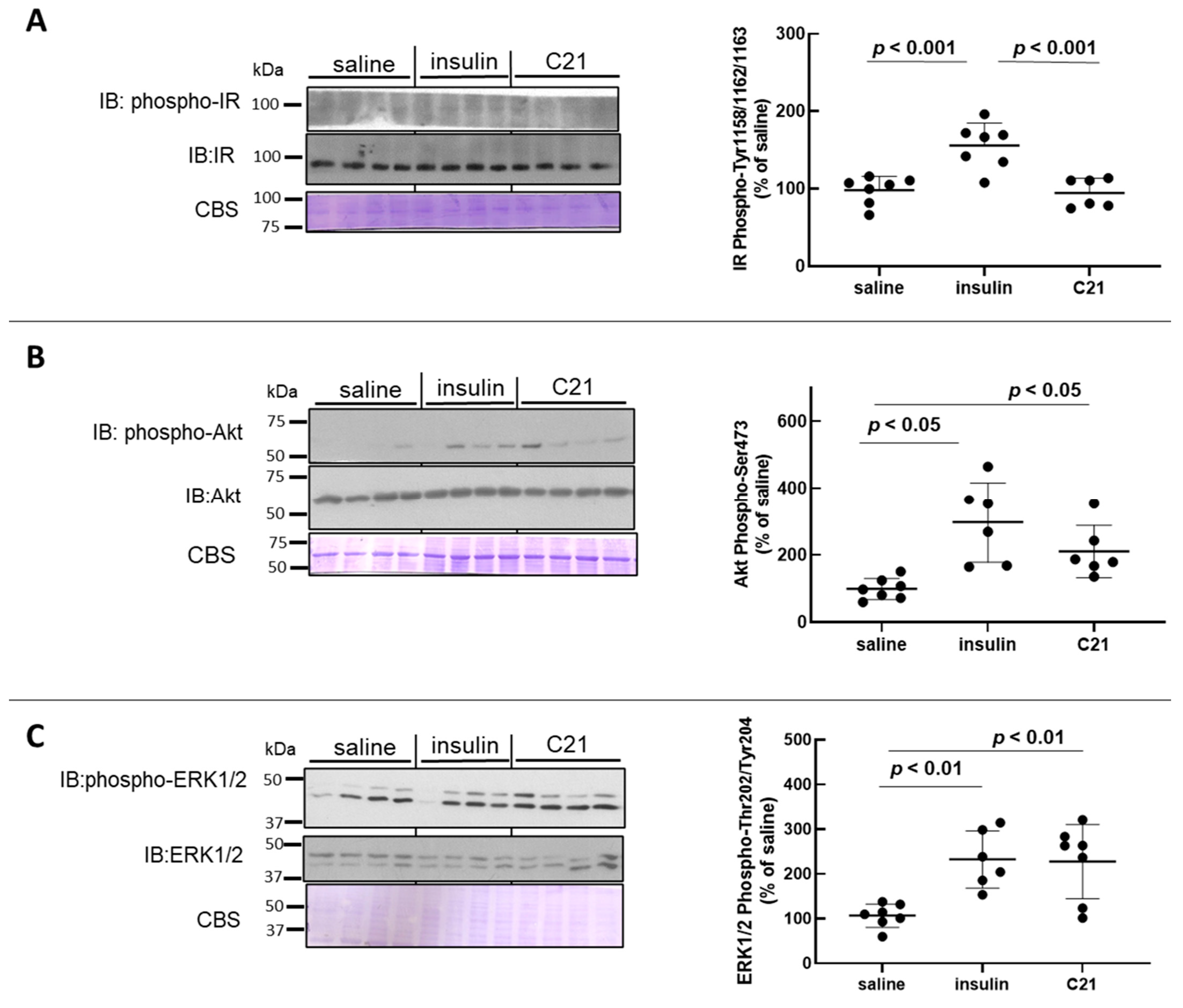
2.1. C21 Induces the Phosphorylation of Akt and ERK1/2 in Mouse White Adipose Tissue (WAT)
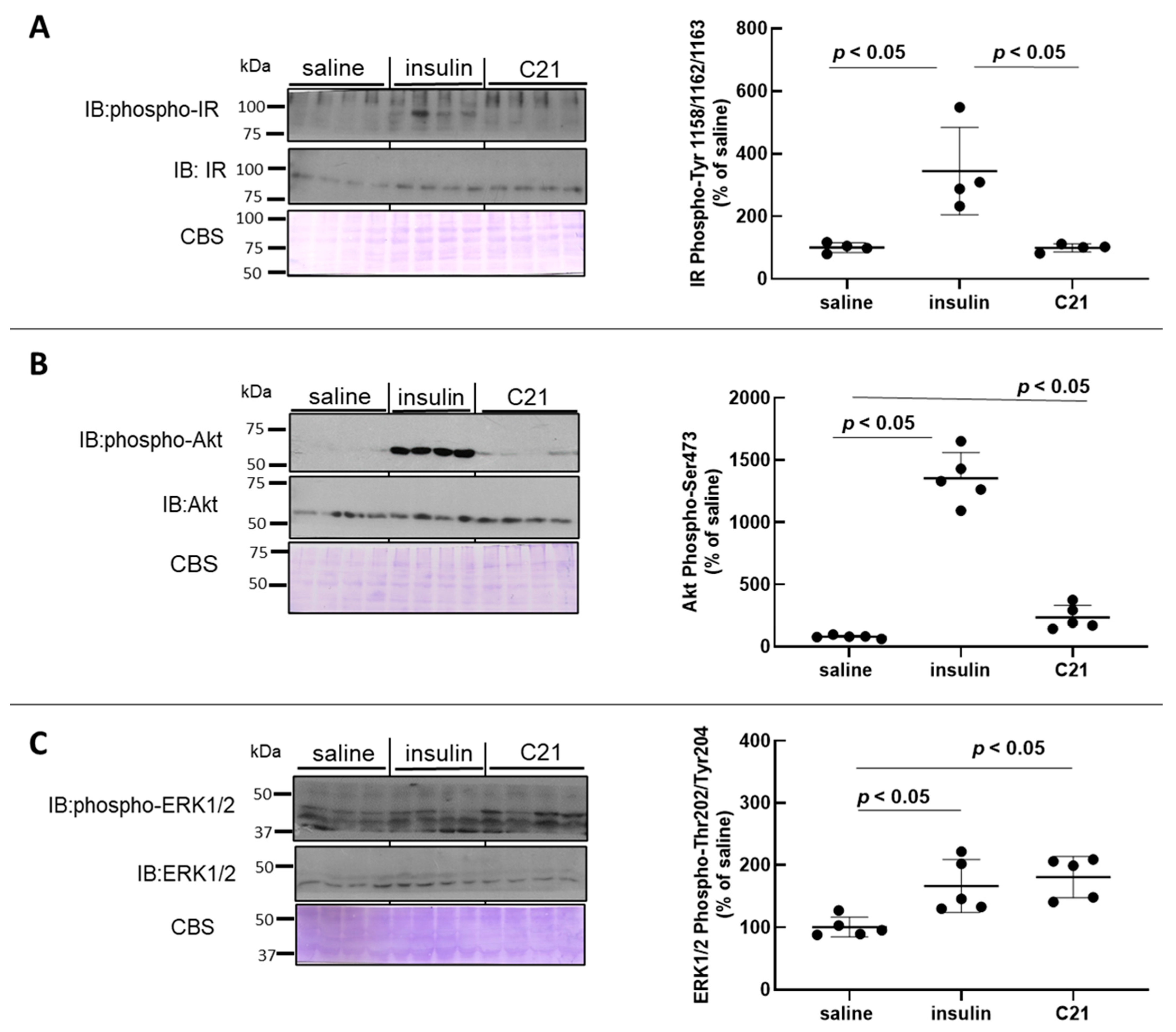
2.2. C21 Induces the Phosphorylation of Akt and ERK1/2 in Mouse Heart
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. In Vivo Administration of C21and Tissue Collection
4.3. Western Blot
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| Ang | Angiotensin |
| AT1R | Angiotensin II receptor type 1 |
| AT2R | Angiotensin II receptor type 2 |
| ATIP | AT2R-interacting protein |
| C21 | Compound 21 |
| GPCR | G protein-coupled receptor |
| IR | Insulin receptor |
| KO | Knockout |
| NO | Nitric oxide |
| PPAR | Peroxisome proliferator-activated receptor |
| NO | Nitric oxide |
| STZ | Streptozotocin |
| WAT | White adipose tissue |
References
- Folli, F.; Kahn, C.R.; Hansen, H.; Bouchie, J.L.; Feene, E.P. Angiotensin II inhibits insulin signaling in aortic smooth muscle cells at multiple levels. A potential role for serine phosphorylation in insulin/angiotensin II crosstalk. J. Clin. Investig. 1997, 100, 2158–2169. [Google Scholar] [CrossRef] [PubMed]
- Henriksen, E.J. Improvement of insulin sensitivity by antagonism of the renin-angiotensin system. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2007, 293, R974–R980. [Google Scholar] [CrossRef] [PubMed]
- Shiuchi, T.; Iwai, M.; Li, H.S.; Wu, L.; Min, L.J.; Li, J.M.; Okumura, M.; Cui, T.X.; Horiuchi, M. Angiotensin II type-1 receptor blocker valsartan enhances insulin sensitivity in skeletal muscles of diabetic mice. Hypertension 2004, 43, 1003–1010. [Google Scholar] [CrossRef] [PubMed]
- Favre, G.A.; Esnault, V.L.; Van Obberghen, E. Modulation of glucose metabolism by the renin-angiotensin-aldosterone system. Am. J. Physiol. Endocrinol. Metab. 2015, 308, E435–E449. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, M.C.; Giani, J.F.; Dominici, F.P.; Turyn, D.; Toblli, J.E. Long-term treatment with an angiotensin II receptor blocker decreases adipocyte size and improves insulin signaling in obese Zucker rats. J. Hypertens. 2009, 27, 2409–2420. [Google Scholar] [CrossRef] [PubMed]
- Steckelings, U.M.; Widdop, R.E.; Sturrock, E.D.; Lubbe, L.; Hussain, T.; Kaschina, E.; Unger, T.; Hallberg, A.; Carey, R.M.; Sumners, C. The angiotensin AT2 receptor: From a binding site to a novel therapeutic target. Pharmacol. Rev. 2022, 74, 1051–1135. [Google Scholar] [CrossRef]
- Fatima, N.; Patel, S.N.; Hussain, T. Angiotensin II type 2 receptor: A target for protection against hypertension, metabolic dysfunction, and organ remodeling. Hypertension 2021, 77, 1845–1856. [Google Scholar] [CrossRef]
- Chai, W.; Wang, W.; Dong, Z.; Cao, W.; Liu, Z. Angiotensin II receptors modulate muscle microvascular and metabolic responses to insulin in vivo. Diabetes 2011, 60, 2939–2946. [Google Scholar] [CrossRef]
- Chai, W.; Wang, W.; Liu, J.; Barrett, E.J.; Carey, R.M.; Cao, W.; Liu, Z. Angiotensin II type 1 and type 2 receptors regulate basal skeletal muscle microvascular volume and glucose use. Hypertension 2010, 55, 523–530. [Google Scholar] [CrossRef]
- Muñoz, M.C.; Burghi, V.; Miquet, J.G.; Cervino, I.A.; Quiroga, D.T.; Mazziotta, L.; Dominici, F.P. Chronic blockade of the AT2 receptor with PD123319 impairs insulin signaling in C57BL/6 mice. Peptides 2017, 88, 37–45. [Google Scholar] [CrossRef]
- Ohshima, K.; Mogi, M.; Jing, F.; Iwanami, J.; Tsukuda, K.; Min, L.-J.; Ogimoto, A.; Dahlöf, B.; Steckelings, U.M.; Unger, T.; et al. Direct angiotensin II type 2 receptor stimulation ameliorates insulin resistance in type 2 diabetes mice with PPARγ activation. PLoS ONE 2012, 7, e483. [Google Scholar] [CrossRef] [PubMed]
- Shum, M.; Pinard, S.; Guimond, M.-O.; Labbé, S.M.; Roberge, C.; Baillargeon, J.-P.; Langlois, M.F.; Alterman, M.; Wallinder, C.; Hallberg, A.; et al. Angiotensin II type 2 receptor promotes adipocyte differentiation and restores adipocyte size in high fat/high-fructose diet-induced insulin resistance in rats. Am. J. Physiol. Endocrinol. Metab. 2013, 304, E197–E210. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.; Yu, L.; Gao, L. Activation of angiotensin type 2 receptors partially ameliorates streptozotocin-induced diabetes in male rats by islet protection. Endocrinology 2014, 155, 793–804. [Google Scholar] [CrossRef] [PubMed]
- Shao, C.; Zucker, I.H.; Gao, L. Angiotensin type 2 receptor in pancreatic islets of adult rats: A novel insulinotropic mediator. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E1281–E1291. [Google Scholar] [CrossRef]
- Koulis, C.; Chow, B.S.M.; McKelvey, M.; Steckelings, U.M.; Unger, T.; Thallas-Bonke, V.; Thomas, M.C.; Cooper, M.E.; Jandeleit-Dahm, K.A.; Allen, T.J. AT2R agonist, compound 21, is reno-protective against type 1 diabetic nephropathy. Hypertension 2015, 65, 1073–1081. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Wang, Y.; Li, X.Y.; Leung, P.S. Angiotensin II type 2 receptor activation with Compound 21 augments islet function and regeneration in streptozotocin-induced neonatal rats and human pancreatic progenitor cells. Pancreas 2017, 46, 395–404. [Google Scholar] [CrossRef]
- Nag, S.; Khan, M.A.; Samuel, P.; Ali, Q.; Hussain, T. Chronic angiotensin AT2R activation prevents high-fat diet-induced adiposity and obesity in female mice independent of estrogen. Metabolism 2015, 64, 814–825. [Google Scholar] [CrossRef]
- Nag, S.; Patel, S.; Mani, S.; Hussain, T. Role of angiotensin type 2 receptor in improving lipid metabolism and preventing adiposity. Mol. Cell. Biochem. 2019, 461, 195–204. [Google Scholar] [CrossRef]
- Quiroga, D.T.; Muñoz, M.C.; Gil, C.; Pffeifer, M.; Toblli, J.E.; Steckelings, U.M.; Giani, J.F.; Dominici, F.P. Chronic administration of the angiotensin type 2 receptor agonist C21 improves insulin sensitivity in C57BL/6 mice. Physiol. Rep. 2018, 6, e13824. [Google Scholar] [CrossRef]
- Dominici, F.P.; Veiras, L.C.; Shen, J.Z.Y.; Bernstein, E.A.; Quiroga, D.T.; Steckelings, U.M.; Bernstein, K.E.; Giani, J.F. Activation of AT2 receptors prevents diabetic complications in female db/db mice by NO-mediated mechanisms. Br. J. Pharmacol. 2020, 177, 4766–4781. [Google Scholar] [CrossRef]
- Samuel, P.; Khan, M.A.; Nag, S.; Inagami, T.; Hussain, T. Angiotensin AT2 receptor contributes towards gender bias in weight gain. PLoS ONE 2013, 8, e48425. [Google Scholar] [CrossRef] [PubMed]
- Quiroga, D.T.; Miquet, J.G.; Gonzalez, L.; Sotelo, A.I.; Muñoz, M.C.; Geraldes, P.M.; Giani, J.F.; Dominici, F.P. Mice lacking angiotensin type 2 receptor exhibit a sex specific attenuation of insulin sensitivity. Mol. Cell. Endocrinol. 2019, 498, 110587. [Google Scholar] [CrossRef] [PubMed]
- Peluso, A.A.; Bertelsen, J.B.; Andersen, K.; Mortsensen, T.P.; Hansen, P.B.; Sumners, C.; Bader, M.; Santos, R.A.; Steckelings, U.M. Identification of protein phosphatase involvement in the AT2 receptor-induced activation of endothelial nitric oxide synthase. Clin. Sci. 2018, 132, 777–790. [Google Scholar] [CrossRef] [PubMed]
- Peluso, A.A.; Kempf, S.J.; Verano-Braga, T.; Rodrigues-Ribeiro, L.; Johansen, L.E.; Hansen, M.R.; Kitlen, G.; Haugaard, A.H.; Sumners, C.; Ditzel, H.J.; et al. Quantitative phosphoproteomics of the angiotensin AT2-Receptor signaling network identifies HDAC1 (histone-deacetylase-1) and p53 as mediators of antiproliferation and apoptosis. Hypertension 2022, 79, 2530–2541. [Google Scholar] [CrossRef] [PubMed]
- Hiromi, H.; Katsutoshi, Y.; Masaoki, T.; Hiroshi, O. Angiotensin type 2 receptor-mediated phosphorylation of eNOS in the aortas of mice with 2-kidney, 1-clip hypertension. Hypertension 2005, 45, 967–973. [Google Scholar]
- Than, A.; Xu, S.; Li, R.; Leow, M.K.-S.; Sun, L.; Chen, P. Angiotensin type 2 receptor activation promotes browning of white adipose tissue and brown adipogenesis. Signal Transduct. Target Ther. 2017, 2, 17022. [Google Scholar] [CrossRef] [PubMed]
- Caruso-Neves, C.; Kwon, S.-H.; Guggino, W.B. Albumin endocytosis in proximal tubule cells is modulated by angiotensin II through an AT2 receptor-mediated protein kinase B activation. Proc. Natl. Acad. Sci. USA 2005, 102, 17513–17518. [Google Scholar] [CrossRef]
- Guimarães-Nobre, C.C.; Mendonça-Reis, E.; Passinho-da-Costa, L.; Miranda-Alves, L.; Berto-Junior, H.C. Signaling pathway in the osmotic resistance induced by angiotensin II AT2 receptor activation in human erythrocytes. Rep. Biochem. Mol. Biol. 2021, 10, 314–326. [Google Scholar] [CrossRef]
- Kulkarni, K.; Patel, S.; Ali, R.; Hussain, P. Angiotensin II type 2 receptor activation preserves megalin in the kidney and prevents proteinuria in high salt diet fed rats. Sci. Rep. 2023, 13, 4277. [Google Scholar] [CrossRef]
- Stroth, U.; Blume, A.; Mielke, K.; Unger, T. Angiotensin AT2 receptor stimulates ERK1 and ERK2 in quiescent but inhibits ERK in NGF-stimulated PC12W cells. Brain Res. Mol. Brain Res. 2000, 78, 175–180. [Google Scholar] [CrossRef]
- Yoshida, T.; Huq, T.S.; Delafontaine, P. Angiotensin Type 2 receptor signaling in satellite cells potentiates skeletal muscle regeneration. J. Biol. Chem. 2014, 289, 26239–26248. [Google Scholar] [CrossRef] [PubMed]
- Hagihara, G.N.; Lobato, N.S.; Filgueira, F.P.; Akamine, E.H.; Aragão, D.S.; Casarini, D.E.; Carvalho, M.H.C.; Fortes, Z.B. Upregulation of ERK1/2-eNOS via AT2 receptors decreases the contractile response to angiotensin II in resistance mesenteric arteries from obese rats. PLoS ONE 2014, 9, e106029. [Google Scholar] [CrossRef] [PubMed]
- Sampson, A.K.; Irvine, J.C.; Shihata, W.A.; Dragoljevic, D.; Lumsden, N.; Huet, O.; Barnes, T.; Unger, T.; Steckelings, U.M.; Jennings, G.L.; et al. Compound 21, a selective agonist of angiotensin AT2 receptors, prevents endothelial inflammation and leukocyte adhesion in vitro and in vivo. Br. J. Pharmacol. 2016, 173, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Castoldi, G.; Carletti, R.; Ippolito, S.; Stella, A.; Zerbini, G.; Pelucchi, S.; Zatti, G.; di Gioia, C.R.T. Angiotensin type 2 and Mas receptor activation prevents myocardial fibrosis and hypertrophy through the reduction of inflammatory cell infiltration and local sympathetic activity in angiotensin II-dependent hypertension. Int. J. Mol. Sci. 2021, 22, 13678. [Google Scholar] [CrossRef] [PubMed]
- Fredgart, M.H.; Leurgans, T.M.; Stenelo, M.; Nybo, M.; Bloksgaard, M.; Lindblad, L.; De Mey, J.G.R.; Steckelings, U.M. The angiotensin AT2-receptor agonist compound 21 is an antagonist for the thromboxane TP-receptor—Implications for preclinical studies and future clinical use. Peptides 2023, 164, 170990. [Google Scholar] [CrossRef] [PubMed]
- Forrester, S.J.; Booz, G.W.; Sigmund, C.D.; Coffman, T.M.; Kawai, T.; Rizzo, V.; Scalia, R.; Eguchi, S. Angiotensin II signal transduction: An update on mechanisms of physiology and pathophysiology. Physiol. Rev. 2018, 98, 1627–1738. [Google Scholar] [CrossRef] [PubMed]
- Sumners, C.; Peluso, A.A.; Haugaard, A.H.; Bertelsen, J.B.; Steckelings, U.M. Anti-fibrotic mechanisms of angiotensin AT2-receptor stimulation. Acta Physiol. 2019, 227, e13280. [Google Scholar] [CrossRef]
- Hilger, D.; Masureel, M.; Kobilka, B.K. Structure and dynamics of GPCR signaling complexes. Nat. Struct. Mol. Biol. 2018, 25, 4–12. [Google Scholar] [CrossRef]
- Shchepinova, M.M.; Hanyaloglu, A.C.; Frost, G.S.; Tate, E.W. Chemical biology of non-canonical G protein-coupled receptor signaling: Toward advanced therapeutics. Curr. Opin. Chem. Biol. 2020, 56, 98–110. [Google Scholar] [CrossRef]
- Turu, G.; Szidonya, L.; Gáborik, Z.; Buday, L.; Spät, A.; Clark, A.J.L.; Hunyady, L. Differential beta-arrestin binding of AT1 and AT2 angiotensin receptors. FEBS Lett. 2006, 580, 41–45. [Google Scholar] [CrossRef]
- Colin, M.; Delaitre, D.; Foulquier, S.; Dupuis, F. The AT1/AT2 receptor equilibrium is a cornerstone of the regulation of the renin angiotensin system beyond the cardiovascular system. Molecules 2023, 28, 5481. [Google Scholar] [CrossRef] [PubMed]
- Goutaudier, R.; Coizet, V.; Carcenac, C.; Carnicella, S. Compound 21, a two-edged sword with both DREADD-selective and off-target outcomes in rats. PLoS ONE 2020, 15, e0238156. [Google Scholar] [CrossRef] [PubMed]
- Bosnyak, S.; Welungoda, I.K.; Hallberg, A.; Alterman, M.; Widdop, R.E.; Jones, E.S. Stimulation of angiotensin AT2 receptors by the nonpeptide agonist, Compound 21, evokes vasodepressor effects in conscious spontaneously hypertensive rats. Br. J. Pharmacol. 2010, 159, 709–716. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Yuan, Z.; Wang, N.; Carey, R.M.; Aylor, K.W.; Chen, L.; Zhou, X.; Liu, Z. Direct activation of angiotensin II type 2 receptors enhances muscle microvascular perfusion, oxygenation, and insulin delivery in male rats. Endocrinology 2018, 159, 685–695. [Google Scholar] [CrossRef]
- Riches, A.C.; Sharp, J.G.; Brynmor Thomas, D.; Vaughan Smith, S. Blood volume determination in the mouse. J. Physiol. 1973, 228, 279–284. [Google Scholar] [CrossRef]
- Welinder, C.; Ekblad, L. Coomassie staining as loading control in Western blot analysis. J. Proteome Res. 2011, 10, 1416–1419. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quiroga, D.T.; Narvaéz Pardo, J.A.; Zubiría, M.G.; Barrales, B.; Muñoz, M.C.; Giovambattista, A.; Dominici, F.P. Acute In Vivo Administration of Compound 21 Stimulates Akt and ERK1/2 Phosphorylation in Mouse Heart and Adipose Tissue. Int. J. Mol. Sci. 2023, 24, 16839. https://doi.org/10.3390/ijms242316839
Quiroga DT, Narvaéz Pardo JA, Zubiría MG, Barrales B, Muñoz MC, Giovambattista A, Dominici FP. Acute In Vivo Administration of Compound 21 Stimulates Akt and ERK1/2 Phosphorylation in Mouse Heart and Adipose Tissue. International Journal of Molecular Sciences. 2023; 24(23):16839. https://doi.org/10.3390/ijms242316839
Chicago/Turabian StyleQuiroga, Diego T., Jorge A. Narvaéz Pardo, María G. Zubiría, Benjamín Barrales, Marina C. Muñoz, Andrés Giovambattista, and Fernando P. Dominici. 2023. "Acute In Vivo Administration of Compound 21 Stimulates Akt and ERK1/2 Phosphorylation in Mouse Heart and Adipose Tissue" International Journal of Molecular Sciences 24, no. 23: 16839. https://doi.org/10.3390/ijms242316839
APA StyleQuiroga, D. T., Narvaéz Pardo, J. A., Zubiría, M. G., Barrales, B., Muñoz, M. C., Giovambattista, A., & Dominici, F. P. (2023). Acute In Vivo Administration of Compound 21 Stimulates Akt and ERK1/2 Phosphorylation in Mouse Heart and Adipose Tissue. International Journal of Molecular Sciences, 24(23), 16839. https://doi.org/10.3390/ijms242316839