

Prostaglandin E2 Exposure Disrupts E-Cadherin/Caveolin-1-Mediated Tumor Suppression to Favor Caveolin-1-Enhanced Migration, Invasion, and Metastasis in Melanoma Models

 ,
,  , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
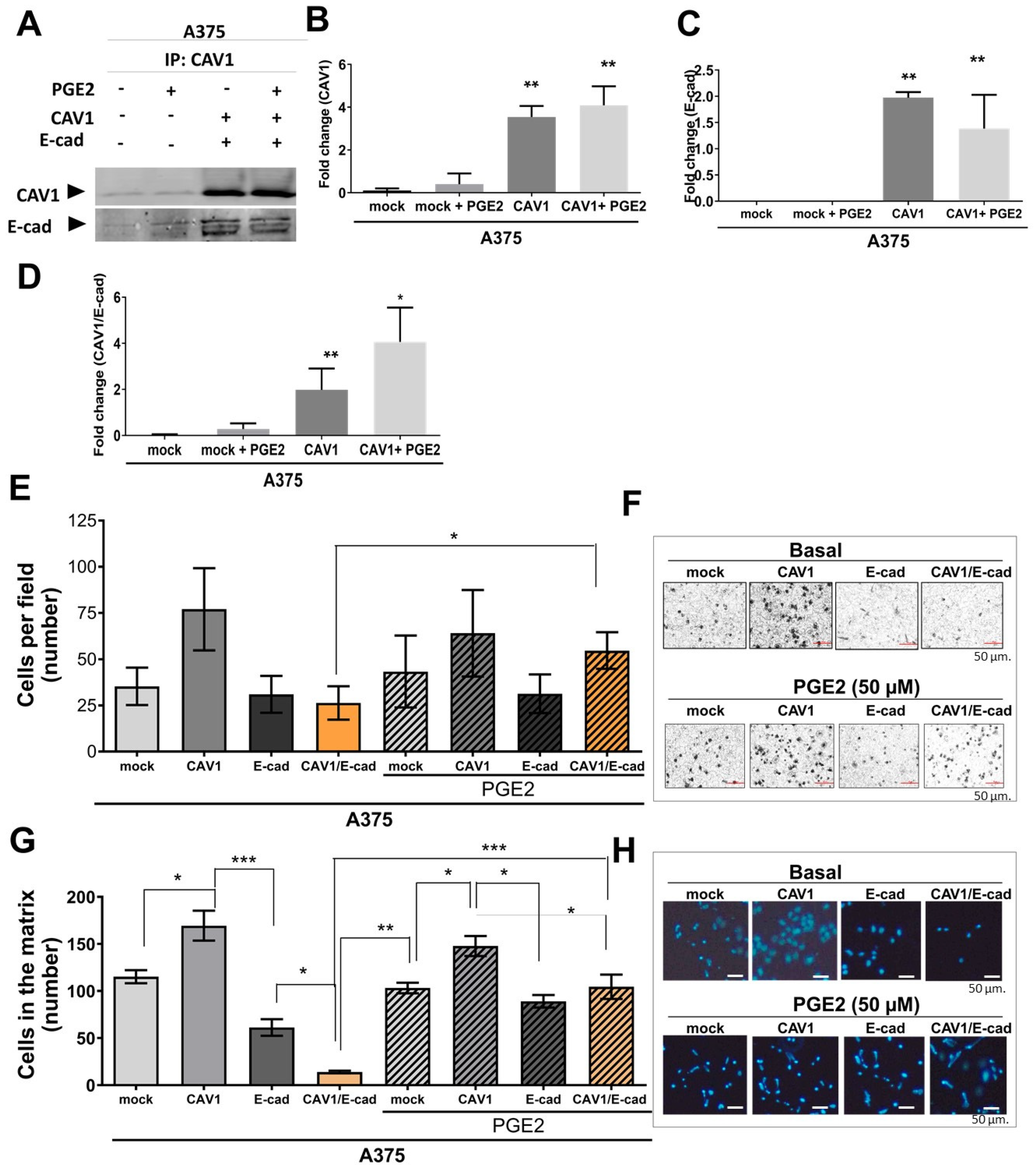
2.1. PGE2 Decreases CAV1/E-Cad Complex Formation and Increases the Phosphorylation of CAV1 on Tyrosine-14
2.2. Exposure of B16F10 (CAV1/E-Cad) Cells to PGE2 Restores CAV1-Enhanced Migration and Invasion
2.3. Tumor Suppression Due to CAV1/E-Cad Co-Expression in B16F10 Cells Ablated via Exposure to PGE2
2.4. Exposure to PGE2 Increases the Migration and Invasion of Human Melanoma Cells
2.5. PGE2 Increases the Colony-Forming Capacity of A375 (CAV1/E-Cad) Cells
2.6. PGE2 Increases the Metastatic Potential of Human Melanoma Cells
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Cell Culture
4.3. Western Blotting
4.4. CAV1 Immunoprecipitation
4.5. Migration and Invasion Assays
4.6. Animal Studies
4.7. Tumor Growth Assays
4.8. Metastasis Assays with Human A375 Melanoma Cells
4.9. Cytokine ELISA
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Neagu, M.; Constantin, C.; Caruntu, C.; Dumitru, C.; Surcel, M.; Zurac, S. Inflammation: A Key Process in Skin Tumorigenesis. Oncol. Lett. 2019, 17, 4068–4084. [Google Scholar] [CrossRef] [PubMed]
- Hölzel, M.; Tüting, T. Inflammation-Induced Plasticity in Melanoma Therapy and Metastasis. Trends Immunol. 2016, 37, 364–374. [Google Scholar] [CrossRef] [PubMed]
- Bald, T.; Quast, T.; Landsberg, J.; Rogava, M.; Glodde, N.; Lopez-Ramos, D.; Kohlmeyer, J.; Riesenberg, S.; van den Boorn-Konijnenberg, D.; Hömig-Hölzel, C.; et al. Ultraviolet-Radiation-Induced Inflammation Promotes Angiotropism and Metastasis in Melanoma. Nature 2014, 507, 109–113. [Google Scholar] [CrossRef]
- Arai, Y.; Matsuura, T.; Matsuura, M.; Fujiwara, M.; Okayasu, I.; Ito, S.; Arihiro, S. Prostaglandin E-Major Urinary Metabolite as a Biomarker for Inflammation in Ulcerative Colitis: Prostaglandins Revisited. Digestion 2016, 93, 32–39. [Google Scholar] [CrossRef]
- Aoki, T.; Narumiya, S. Prostaglandin E2-EP2 Signaling as a Node of Chronic Inflammation in the Colon Tumor Microenvironment. Inflamm. Regen. 2017, 37, 4. [Google Scholar] [CrossRef] [PubMed]
- Montrose, D.C.; Nakanishi, M.; Murphy, R.C.; Zarini, S.; McAleer, J.P.; Vella, A.T.; Rosenberg, D.W. The Role of PGE2 in Intestinal Inflammation and Tumorigenesis. Prostaglandins Other Lipid Mediat. 2015, 116–117, 26–36. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; DuBois, R.N. Eicosanoids and Cancer. Nat. Rev. Cancer 2010, 10, 181–193. [Google Scholar] [CrossRef]
- Wang, D.; Fu, L.; Sun, H.; Guo, L.; DuBois, R.N. Prostaglandin E2 Promotes Colorectal Cancer Stem Cell Expansion and Metastasis in Mice. Gastroenterology 2015, 149, 1884–1895.e4. [Google Scholar] [CrossRef]
- Stamatakis, K.; Jimenez-Martinez, M.; Jimenez-Segovia, A.; Chico-Calero, I.; Conde, E.; Galán-Martínez, J.; Ruiz, J.; Pascual, A.; Barrocal, B.; López-Pérez, R.; et al. Prostaglandins Induce Early Growth Response 1 Transcription Factor Mediated Microsomal Prostaglandin E2 Synthase Up-Regulation for Colorectal Cancer Progression. Oncotarget 2015, 6, 39941–39959. [Google Scholar] [CrossRef]
- Kim, S.-H.; Hashimoto, Y.; Cho, S.-N.; Roszik, J.; Milton, D.R.; Dal, F.; Kim, S.F.; Menter, D.G.; Yang, P.; Ekmekcioglu, S.; et al. Microsomal PGE2 Synthase-1 Regulates Melanoma Cell Survival and Associates with Melanoma Disease Progression. Pigment Cell Melanoma Res. 2016, 29, 297–308. [Google Scholar] [CrossRef]
- Zang, S.; Ni, M.; Lian, Y.; Zhang, Y.; Liu, J.; Huang, A. Expression of Microsomal Prostaglandin E2 Synthase-1 and Its Role in Human Hepatocellular Carcinoma. Hum. Pathol. 2013, 44, 1681–1687. [Google Scholar] [CrossRef] [PubMed]
- Finetti, F.; Travelli, C.; Ercoli, J.; Colombo, G.; Buoso, E.; Trabalzini, L. Prostaglandin E2 and Cancer: Insight into Tumor Progression and Immunity. Biology 2020, 9, 434. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Xu, X.; Jiang, M.; Bi, Y.; Xu, J.; Han, M. Lipopolysaccharide Induces Inflammation and Facilitates Lung Metastasis in a Breast Cancer Model via the Prostaglandin E2-EP2 Pathway. Mol. Med. Rep. 2015, 11, 4454–4462. [Google Scholar] [CrossRef]
- Long, M.D.; Herfarth, H.H.; Pipkin, C.A.; Porter, C.Q.; Sandler, R.S.; Kappelman, M.D. Increased Risk for Non-Melanoma Skin Cancer in Patients with Inflammatory Bowel Disease. Clin. Gastroenterol. Hepatol. 2010, 8, 268–274. [Google Scholar] [CrossRef]
- Kim, S.-H.; Jason, R.; Cho, S.; Ogata, D.; Milton, D.R.; Peng, W.; Menter, D.G.; Ekmekcioglu, S.; Grimm, E.A. The COX2 Effector Microsomal PGE2 Synthase-1 Is a Regulator of Immunosuppression in Cutaneous Melanoma. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2019, 25, 1650–1663. [Google Scholar] [CrossRef] [PubMed]
- Buckman, S.Y.; Gresham, A.; Hale, P.; Hruza, G.; Anast, J.; Masferrer, J.; Pentland, A.P. COX-2 Expression Is Induced by UVB Exposure in Human Skin: Implications for the Development of Skin Cancer. Carcinogenesis 1998, 19, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Kim, K.J. Upregulation of Prostaglandin E2 by Inducible Microsomal Prostaglandin E Synthase-1 in Colon Cancer. Ann. Coloproctology 2022, 38, 153–159. [Google Scholar] [CrossRef]
- Karpisheh, V.; Nikkhoo, A.; Hojjat-Farsangi, M.; Namdar, A.; Azizi, G.; Ghalamfarsa, G.; Sabz, G.; Yousefi, M.; Yousefi, B.; Jadidi-Niaragh, F. Prostaglandin E2 as a Potent Therapeutic Target for Treatment of Colon Cancer. Prostaglandins Other Lipid Mediat. 2019, 144, 106338. [Google Scholar] [CrossRef]
- Xue, X.; Shah, Y.M. Hypoxia-Inducible Factor-2α Is Essential in Activating the COX2/MPGES-1/PGE2 Signaling Axis in Colon Cancer. Carcinogenesis 2013, 34, 163–169. [Google Scholar] [CrossRef]
- Sheng, J.; Sun, H.; Yu, F.B.; Li, B.; Zhang, Y.; Zhu, Y.T. The Role of Cyclooxygenase-2 in Colorectal Cancer. Int. J. Med. Sci. 2020, 17, 1095. [Google Scholar] [CrossRef]
- Mitsui, Y.; Tomonobu, N.; Watanabe, M.; Kinoshita, R.; Sumardika, I.W.; Youyi, C.; Murata, H.; Yamamoto, K.-I.; Sadahira, T.; Rodrigo, A.G.H.; et al. Upregulation of Mobility in Pancreatic Cancer Cells by Secreted S100A11 through Activation of Surrounding Fibroblasts. Oncol. Res. 2019, 27, 945–956. [Google Scholar] [CrossRef]
- Schaeuble, K.; Cannelle, H.; Favre, S.; Huang, H.-Y.; Oberle, S.G.; Speiser, D.E.; Zehn, D.; Luther, S.A. Attenuation of Chronic Antiviral T-Cell Responses through Constitutive COX2-Dependent Prostanoid Synthesis by Lymph Node Fibroblasts. PLoS Biol. 2019, 17, e3000072. [Google Scholar] [CrossRef]
- Castellone, M.D.; Teramoto, H.; Williams, B.O.; Druey, K.M.; Gutkind, J.S. Prostaglandin E2 Promotes Colon Cancer Cell Growth through a Gs-Axin-Beta-Catenin Signaling Axis. Science 2005, 310, 1504–1510. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Jung, C.; Liu, C.; Sheng, H. Prostaglandin E2 Stimulates the Beta-Catenin/T Cell Factor-Dependent Transcription in Colon Cancer. J. Biol. Chem. 2005, 280, 26565–26572. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Li, C.; Li, M.; Tong, X.; Hu, X.; Yang, X.; Yan, X.; He, L.; Wan, C. CYP2S1 Depletion Enhances Colorectal Cell Proliferation Is Associated with PGE2-Mediated Activation of β-Catenin Signaling. Exp. Cell Res. 2015, 331, 377–386. [Google Scholar] [CrossRef]
- Fröhlich, J.; Rose, K.; Hecht, A. Transcriptional Activity Mediated by β-CATENIN and TCF/LEF Family Members Is Completely Dispensable for Survival and Propagation of Multiple Human Colorectal Cancer Cell Lines. Sci. Rep. 2023, 13, 287. [Google Scholar] [CrossRef] [PubMed]
- Shao, J.; Lee, S.B.; Guo, H.; Evers, B.M.; Sheng, H. Prostaglandin E2 Stimulates the Growth of Colon Cancer Cells via Induction of Amphiregulin. Cancer Res. 2003, 63, 5218–5223. [Google Scholar]
- Dufour, M.; Faes, S.; Dormond-Meuwly, A.; Demartines, N.; Dormond, O. PGE2-Induced Colon Cancer Growth Is Mediated by MTORC1. Biochem. Biophys. Res. Commun. 2014, 451, 587–591. [Google Scholar] [CrossRef]
- Sheng, H.; Shao, J.; Morrow, J.D.; Beauchamp, R.D.; DuBois, R.N. Modulation of Apoptosis and Bcl-2 Expression by Prostaglandin E2 in Human Colon Cancer Cells. Cancer Res. 1998, 58, 362–366. [Google Scholar]
- Mori, A.; Saito, M.; Sakamoto, K.; Narita, M.; Nakahara, T.; Ishii, K. Stimulation of Prostanoid IP and EP(2) Receptors Dilates Retinal Arterioles and Increases Retinal and Choroidal Blood Flow in Rats. Eur. J. Pharmacol. 2007, 570, 135–141. [Google Scholar] [CrossRef]
- Krysan, K.; Dalwadi, H.; Sharma, S.; Põld, M.; Dubinett, S. Cyclooxygenase 2-Dependent Expression of Survivin Is Critical for Apoptosis Resistance in Non-Small Cell Lung Cancer. Cancer Res. 2004, 64, 6359–6362. [Google Scholar] [CrossRef] [PubMed]
- Krysan, K.; Merchant, F.H.; Zhu, L.; Dohadwala, M.; Luo, J.; Lin, Y.; Heuze-Vourc’h, N.; Põld, M.; Seligson, D.; Chia, D.; et al. COX-2-Dependent Stabilization of Survivin in Non-Small Cell Lung Cancer. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 2004, 18, 206–208. [Google Scholar] [CrossRef]
- Zhang, T.; Otevrel, T.; Gao, Z.; Gao, Z.; Ehrlich, S.M.; Fields, J.Z.; Boman, B.M. Evidence That APC Regulates Survivin Expression: A Possible Mechanism Contributing to the Stem Cell Origin of Colon Cancer. Cancer Res. 2001, 61, 8664–8667. [Google Scholar] [PubMed]
- Torres, V.A.; Tapia, J.C.; Rodríguez, D.A.; Párraga, M.; Lisboa, P.; Montoya, M.; Leyton, L.; Quest, A.F.G. Caveolin-1 Controls Cell Proliferation and Cell Death by Suppressing Expression of the Inhibitor of Apoptosis Protein Survivin. J. Cell Sci. 2006, 119, 1812–1823. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, D.A.; Tapia, J.C.; Fernandez, J.G.; Torres, V.A.; Muñoz, N.; Galleguillos, D.; Leyton, L.; Quest, A.F.G. Caveolin-1-Mediated Suppression of Cyclooxygenase-2 via a Beta-Catenin-Tcf/Lef-Dependent Transcriptional Mechanism Reduced Prostaglandin E2 Production and Survivin Expression. Mol. Biol. Cell 2009, 20, 2297–2310. [Google Scholar] [CrossRef] [PubMed]
- Venza, I.; Visalli, M.; Oteri, R.; Beninati, C.; Teti, D.; Venza, M. Genistein Reduces Proliferation of EP3-Expressing Melanoma Cells through Inhibition of PGE2-Induced IL-8 Expression. Int. Immunopharmacol. 2018, 62, 86–95. [Google Scholar] [CrossRef]
- Vaid, M.; Singh, T.; Prasad, R.; Kappes, J.C.; Katiyar, S.K. Therapeutic Intervention of Proanthocyanidins on the Migration Capacity of Melanoma Cells Is Mediated through PGE2 Receptors and β-Catenin Signaling Molecules. Am. J. Cancer Res. 2015, 5, 3325–3338. [Google Scholar]
- Bender, F.C.; Reymond, M.A.; Bron, C.; Quest, A.F. Caveolin-1 Levels Are down-Regulated in Human Colon Tumors, and Ectopic Expression of Caveolin-1 in Colon Carcinoma Cell Lines Reduces Cell Tumorigenicity. Cancer Res. 2000, 60, 5870–5878. [Google Scholar]
- Lobos-González, L.; Aguilar, L.; Diaz, J.; Diaz, N.; Urra, H.; Torres, V.A.; Silva, V.; Fitzpatrick, C.; Lladser, A.; Hoek, K.S.; et al. E-Cadherin Determines Caveolin-1 Tumor Suppression or Metastasis Enhancing Function in Melanoma Cells. Pigment Cell Melanoma Res. 2013, 26, 555–570. [Google Scholar] [CrossRef]
- Lobos-Gonzalez, L.; Aguilar-Guzmán, L.; Fernandez, J.G.; Muñoz, N.; Hossain, M.; Bieneck, S.; Silva, V.; Burzio, V.; Sviderskaya, E.V.; Bennett, D.C.; et al. Caveolin-1 Is a Risk Factor for Postsurgery Metastasis in Preclinical Melanoma Models. Melanoma Res. 2014, 24, 108–119. [Google Scholar] [CrossRef]
- Bender, F.; Montoya, M.; Monardes, V.; Leyton, L.; Quest, A.F.G. Caveolae and Caveolae-like Membrane Domains in Cellular Signaling and Disease: Identification of Downstream Targets for the Tumor Suppressor Protein Caveolin-1. Biol. Res. 2002, 35, 151–167. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.-Y.; Wu, J.-X.; Wu, J.-S.; Pan, Y.-T.; Jin, R. Caveolin-1, E-Cadherin and β-Catenin in Gastric Carcinoma, Precancerous Tissues and Chronic Non-Atrophic Gastritis. Chin. J. Cancer Res. Chung-Kuo Yen Cheng Yen Chiu 2012, 24, 23–28. [Google Scholar] [CrossRef] [PubMed]
- Fernández, J.G.; Rodríguez, D.A.; Valenzuela, M.; Calderon, C.; Urzúa, U.; Munroe, D.; Rosas, C.; Lemus, D.; Díaz, N.; Wright, M.C.; et al. Survivin Expression Promotes VEGF-Induced Tumor Angiogenesis via PI3K/Akt Enhanced β-Catenin/Tcf-Lef Dependent Transcription. Mol. Cancer 2014, 13, 209. [Google Scholar] [CrossRef]
- Torres, V.A.; Tapia, J.C.; Rodriguez, D.A.; Lladser, A.; Arredondo, C.; Leyton, L.; Quest, A.F.G. E-Cadherin Is Required for Caveolin-1-Mediated down-Regulation of the Inhibitor of Apoptosis Protein Survivin via Reduced Beta-Catenin-Tcf/Lef-Dependent Transcription. Mol. Cell. Biol. 2007, 27, 7703–7717. [Google Scholar] [CrossRef] [PubMed]
- Diaz, J.; Mendoza, P.; Ortiz, R.; Díaz, N.; Leyton, L.; Stupack, D.; Quest, A.F.; Torres, V.A. Rab5 Is Required in Metastatic Cancer Cells for Caveolin-1-Enhanced Rac1 Activation, Migration and Invasion. J. Cell Sci. 2014, 127, 2401–2406. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, R.; Díaz, J.; Díaz, N.; Lobos-Gonzalez, L.; Cárdenas, A.; Contreras, P.; Díaz, M.I.; Otte, E.; Cooper-White, J.; Torres, V.; et al. Extracellular Matrix-Specific Caveolin-1 Phosphorylation on Tyrosine 14 Is Linked to Augmented Melanoma Metastasis but Not Tumorigenesis. Oncotarget 2016, 7, 40571–40593. [Google Scholar] [CrossRef]
- Ortiz, R.; Díaz, J.; Díaz-Valdivia, N.; Martínez, S.; Simón, L.; Contreras, P.; Lobos-González, L.; Guerrero, S.; Leyton, L.; Quest, A.F. Src-Family Kinase Inhibitors Block Early Steps of Caveolin-1-Enhanced Lung Metastasis by Melanoma Cells. Biochem. Pharmacol. 2020, 177, 113941. [Google Scholar] [CrossRef]
- Urra, H.; Torres, V.A.; Ortiz, R.J.; Lobos, L.; Díaz, M.I.; Díaz, N.; Härtel, S.; Leyton, L.; Quest, A.F.G. Caveolin-1-Enhanced Motility and Focal Adhesion Turnover Require Tyrosine-14 but Not Accumulation to the Rear in Metastatic Cancer Cells. PLoS ONE 2012, 7, e33085. [Google Scholar] [CrossRef]
- Borowicz, S.; Van Scoyk, M.; Avasarala, S.; Rathinam, M.K.K.; Tauler, J.; Bikkavilli, R.K.; Winn, R.A. The soft agar colony formation assay. JoVE J. Vis. Exp. 2014, 92, e51998. [Google Scholar]
- Quest, A.F.G.; Lobos-González, L.; Nuñez, S.; Sanhueza, C.; Fernández, J.-G.; Aguirre, A.; Rodríguez, D.; Leyton, L.; Torres, V. The Caveolin-1 Connection to Cell Death and Survival. Curr. Mol. Med. 2013, 13, 266–281. [Google Scholar] [CrossRef]
- Li, S.; Seitz, R.; Lisanti, M.P. Phosphorylation of Caveolin by Src Tyrosine Kinases. The Alpha-Isoform of Caveolin Is Selectively Phosphorylated by v-Src in Vivo. J. Biol. Chem. 1996, 271, 3863–3868. [Google Scholar] [CrossRef] [PubMed]
- Cao, H.; Courchesne, W.E.; Mastick, C.C. A Phosphotyrosine-Dependent Protein Interaction Screen Reveals a Role for Phosphorylation of Caveolin-1 on Tyrosine 14: Recruitment of C-Terminal Src Kinase. J. Biol. Chem. 2002, 277, 8771–8774. [Google Scholar] [CrossRef] [PubMed]
- Núñez-Wehinger, S.; Ortiz, R.J.; Díaz, N.; Díaz, J.; Lobos-González, L.; Quest, A.F.G. Caveolin-1 in Cell Migration and Metastasis. Curr. Mol. Med. 2014, 14, 255–274. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.I.; Lakshmikanthan, V.; Frilot, N.; Daaka, Y. Prostaglandin E2 Promotes Lung Cancer Cell Migration via EP4-BetaArrestin1-c-Src Signalsome. Mol. Cancer Res. MCR 2010, 8, 569–577. [Google Scholar] [CrossRef]
- Linge, A.; Morishima, N.; Kasper, M.; Barth, K. Bleomycin Induces Caveolin-1 and -2 Expression in Epithelial Lung Cancer A549 Cells. Anticancer Res. 2007, 27, 1343–1351. [Google Scholar]
- Díaz-Valdivia, N.I.; Díaz, J.; Contreras, P.; Campos, A.; Rojas-Celis, V.; Burgos-Ravanal, R.A.; Lobos-González, L.; Torres, V.A.; Perez, V.I.; Frei, B.; et al. The Non-Receptor Tyrosine Phosphatase Type 14 Blocks Caveolin-1-Enhanced Cancer Cell Metastasis. Oncogene 2020, 39, 3693–3709. [Google Scholar] [CrossRef]
- Webb, D.J.; Zhang, H.; Horwitz, A.F. Cell Migration: An Overview. Methods Mol. Biol. 2005, 294, 3–11. [Google Scholar]
- Felicetti, F.; Parolini, I.; Bottero, L.; Fecchi, K.; Errico, M.C.; Raggi, C.; Biffoni, M.; Spadaro, F.; Lisanti, M.P.; Sargiacomo, M.; et al. Caveolin-1 Tumor-Promoting Role in Human Melanoma. Int. J. Cancer 2009, 125, 1514–1522. [Google Scholar] [CrossRef]
- Goetz, J.G.; Joshi, B.; Lajoie, P.; Strugnell, S.S.; Scudamore, T.; Kojic, L.D.; Nabi, I.R. Concerted Regulation of Focal Adhesion Dynamics by Galectin-3 and Tyrosine-Phosphorylated Caveolin-1. J. Cell Biol. 2008, 180, 1261–1275. [Google Scholar] [CrossRef]
- Park, A.; Lee, Y.; Kim, M.S.; Kang, Y.J.; Park, Y.J.; Jung, H.; Kim, T.D.; Lee, H.G.; Choi, I.; Yoon, S.R. Prostaglandin E2 Secreted by Thyroid Cancer Cells Contributes to Immune Escape Through the Suppression of Natural Killer (NK) Cell Cytotoxicity and NK Cell Differentiation. Front. Immunol. 2018, 9, 1859. [Google Scholar] [CrossRef]
- Zhou, M.; Wang, A.; Yin, B.; Wu, D.; Han, S.; Zhang, W.; Liu, J.; Sun, K. SND1 Promotes the Proliferation of Osteosarcoma Cells by Upregulating COX-2/PGE2 Expression via Activation of NF-κB. Oncol. Rep. 2019, 41, 579–589. [Google Scholar] [CrossRef] [PubMed]
- Cheng, S.Y.; Zhang, H.; Zhang, M.; Xia, S.K.; Bai, X.M.; Zhang, L.; Ma, J.; Rong, R.; Wang, Y.P.; Du, M.Z.; et al. Prostaglandin E2 Receptor EP2 Mediates Snail Expression in Hepatocellular Carcinoma Cells. Oncol. Rep. 2014, 31, 2099–2106. [Google Scholar] [CrossRef] [PubMed]
- O’Callaghan, G.; Houston, A. Prostaglandin E2 and the EP Receptors in Malignancy: Possible Therapeutic Targets? Br. J. Pharmacol. 2015, 172, 5239–5250. [Google Scholar] [CrossRef] [PubMed]
- Dohadwala, M.; Yang, S.-C.; Luo, J.; Sharma, S.; Batra, R.K.; Huang, M.; Lin, Y.; Goodglick, L.; Krysan, K.; Fishbein, M.C.; et al. Cyclooxygenase-2-Dependent Regulation of E-Cadherin: Prostaglandin E(2) Induces Transcriptional Repressors ZEB1 and Snail in Non-Small Cell Lung Cancer. Cancer Res. 2006, 66, 5338–5345. [Google Scholar] [CrossRef]
- Kawamori, T.; Uchiya, N.; Sugimura, T.; Wakabayashi, K. Enhancement of Colon Carcinogenesis by Prostaglandin E2 Administration. Carcinogenesis 2003, 24, 985–990. [Google Scholar] [CrossRef]
- Wang, D.; Buchanan, F.G.; Wang, H.; Dey, S.K.; DuBois, R.N. Prostaglandin E2 Enhances Intestinal Adenoma Growth via Activation of the Ras-Mitogen-Activated Protein Kinase Cascade. Cancer Res. 2005, 65, 1822–1829. [Google Scholar] [CrossRef]
- Smuđ Orehovec, S.; Dujmović, A.; Mijatović, D.; Mance, M.; Šarčević, B. Immunohistochemical Expression of Matrix Metalloproteinase-1 and Cyclooxygenase-2 in Cutaneous Squamous Cell and Basal Cell Carcinoma. Acta Dermatovenerol. Croat. 2021, 291, 8–20. [Google Scholar]
- Kiraly, A.J.; Soliman, E.; Jenkins, A.; Van Dross, R.T. Apigenin inhibits COX-2, PGE2, and EP1 and also initiates terminal differentiation in the epidermis of tumor bearing mice. Prostaglandins Leukot. Essent. Fatty Acids 2016, 104, 44–53. [Google Scholar] [CrossRef]
- Rundhaug, J.E.; Simper, M.S.; Surh, I.; Fischer, S.M. The role of the EP receptors for prostaglandin E2 in skin and skin cancer. Cancer Metastasis Rev. 2011, 30, 465–480. [Google Scholar] [CrossRef]
- Inada, M.; Takita, M.; Yokoyama, S.; Watanabe, K.; Tominari, T.; Matsumoto, C.; Hirata, M.; Maru, Y.; Maruyama, T.; Sugimoto, Y.; et al. Direct Melanoma Cell Contact Induces Stromal Cell Autocrine Prostaglandin E2-EP4 Receptor Signaling that Drives Tumor Growth, Angiogenesis, and Metastasis. J. Biol. Chem. 2015, 290, 29781–29793. [Google Scholar] [CrossRef]
- Lobos-González, L.; Silva, V.; Araya, M.; Restovic, F.; Echenique, J.; Oliveira-Cruz, L.; Fitzpatrick, C.; Briones, M.; Villegas, J.; Villota, C.; et al. Targeting Antisense Mitochondrial NcRNAs Inhibits Murine Melanoma Tumor Growth and Metastasis through Reduction in Survival and Invasion Factors. Oncotarget 2016, 7, 58331. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, C.; Bendek, M.F.; Briones, M.; Farfán, N.; Silva, V.A.; Nardocci, G.; Montecino, M.; Boland, A.; Deleuze, J.-F.; Villegas, J.; et al. Mitochondrial NcRNA Targeting Induces Cell Cycle Arrest and Tumor Growth Inhibition of MDA-MB-231 Breast Cancer Cells through Reduction of Key Cell Cycle Progression Factors. Cell Death Dis. 2019, 10, 423. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lobos-González, L.; Oróstica, L.; Díaz-Valdivia, N.; Rojas-Celis, V.; Campos, A.; Duran-Jara, E.; Farfán, N.; Leyton, L.; Quest, A.F.G. Prostaglandin E2 Exposure Disrupts E-Cadherin/Caveolin-1-Mediated Tumor Suppression to Favor Caveolin-1-Enhanced Migration, Invasion, and Metastasis in Melanoma Models. Int. J. Mol. Sci. 2023, 24, 16947. https://doi.org/10.3390/ijms242316947
Lobos-González L, Oróstica L, Díaz-Valdivia N, Rojas-Celis V, Campos A, Duran-Jara E, Farfán N, Leyton L, Quest AFG. Prostaglandin E2 Exposure Disrupts E-Cadherin/Caveolin-1-Mediated Tumor Suppression to Favor Caveolin-1-Enhanced Migration, Invasion, and Metastasis in Melanoma Models. International Journal of Molecular Sciences. 2023; 24(23):16947. https://doi.org/10.3390/ijms242316947
Chicago/Turabian StyleLobos-González, Lorena, Lorena Oróstica, Natalia Díaz-Valdivia, Victoria Rojas-Celis, America Campos, Eduardo Duran-Jara, Nicole Farfán, Lisette Leyton, and Andrew F. G. Quest. 2023. "Prostaglandin E2 Exposure Disrupts E-Cadherin/Caveolin-1-Mediated Tumor Suppression to Favor Caveolin-1-Enhanced Migration, Invasion, and Metastasis in Melanoma Models" International Journal of Molecular Sciences 24, no. 23: 16947. https://doi.org/10.3390/ijms242316947
APA StyleLobos-González, L., Oróstica, L., Díaz-Valdivia, N., Rojas-Celis, V., Campos, A., Duran-Jara, E., Farfán, N., Leyton, L., & Quest, A. F. G. (2023). Prostaglandin E2 Exposure Disrupts E-Cadherin/Caveolin-1-Mediated Tumor Suppression to Favor Caveolin-1-Enhanced Migration, Invasion, and Metastasis in Melanoma Models. International Journal of Molecular Sciences, 24(23), 16947. https://doi.org/10.3390/ijms242316947