
circITGB1 Regulates Adipocyte Proliferation and Differentiation via the miR-23a/ARRB1 Pathway

 ,
,
Abstract
1. Introduction
2. Results
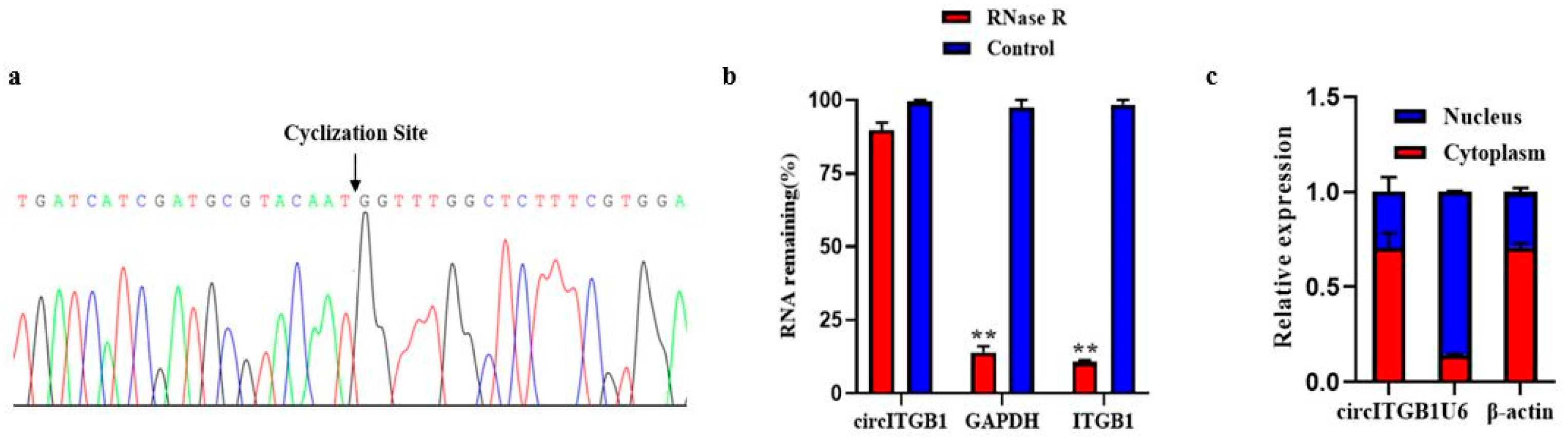
2.1. Characterization of circITGB1 in Adipocytes
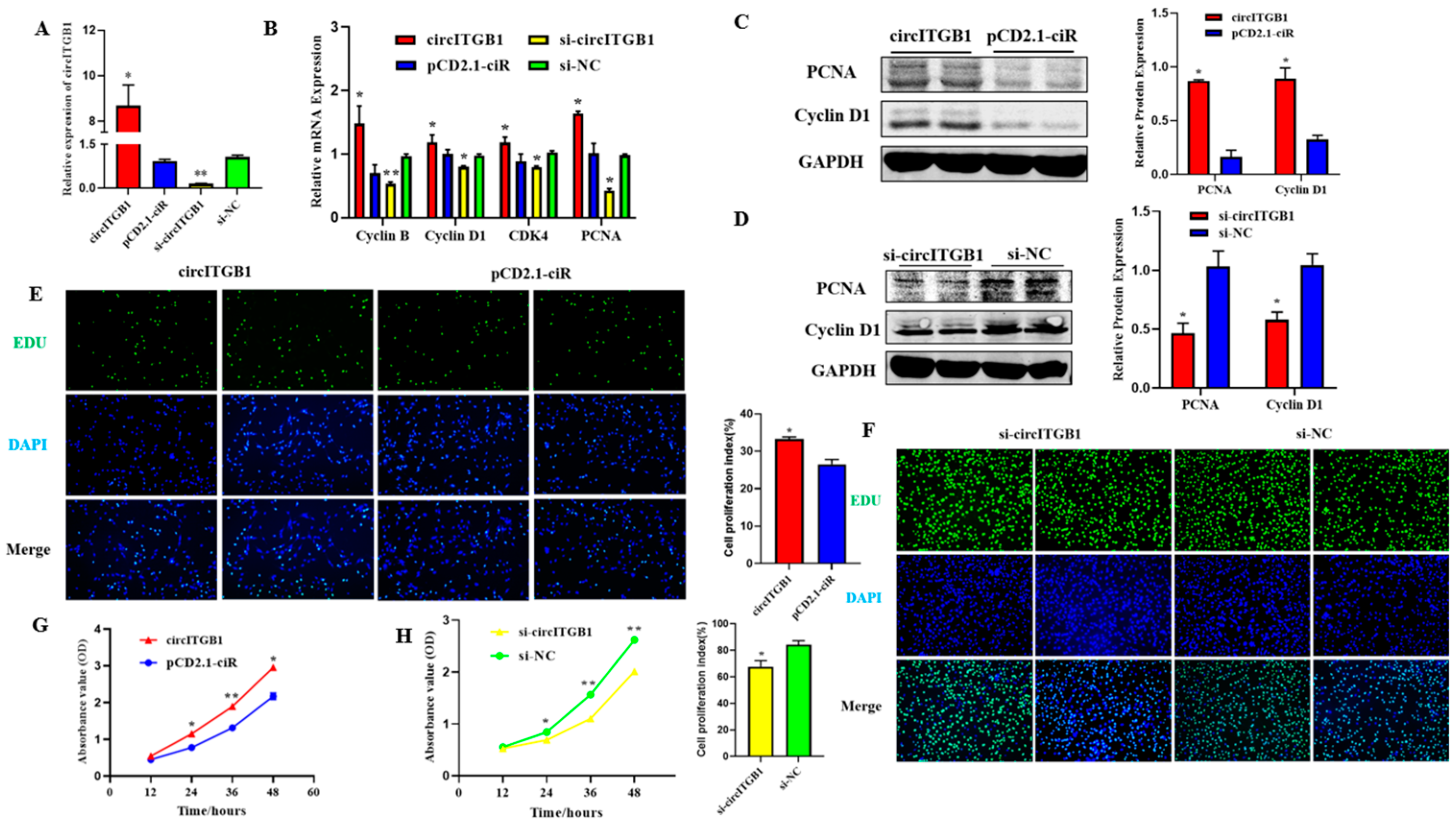
2.2. CircITGB1 Promotes Adipocytes Proliferation
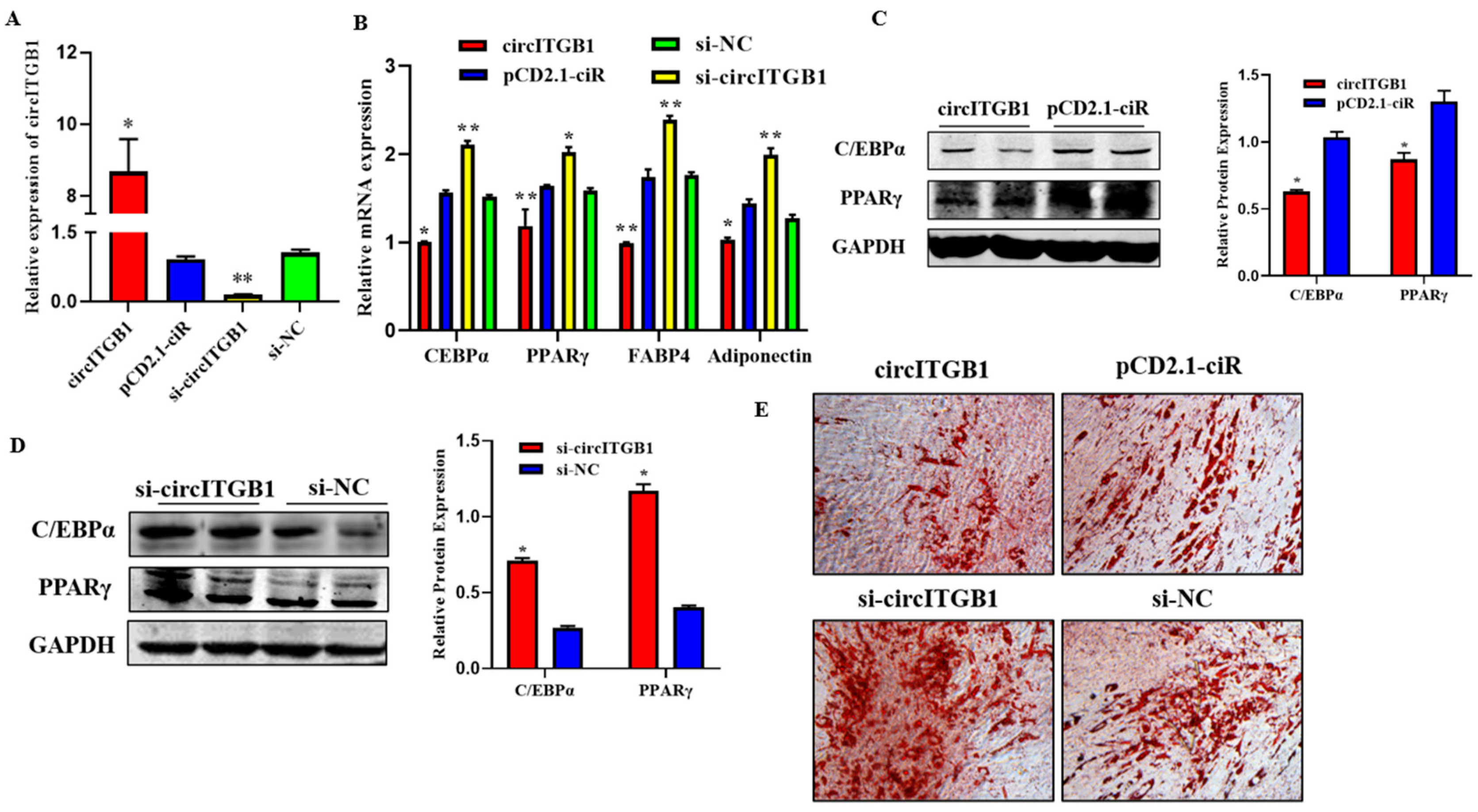
2.3. CircITGB1 Inhibits Adipocytes Differentiation
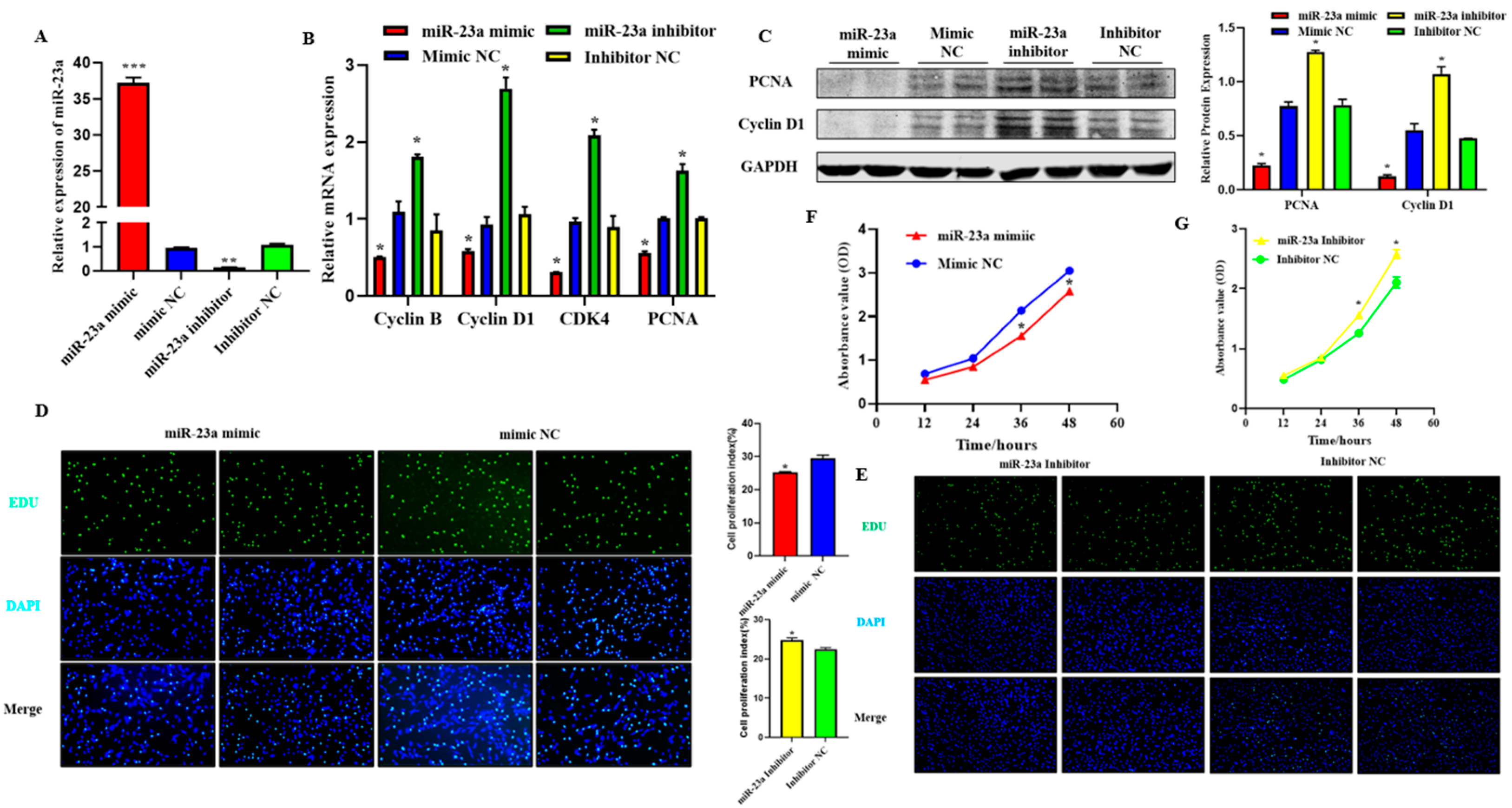
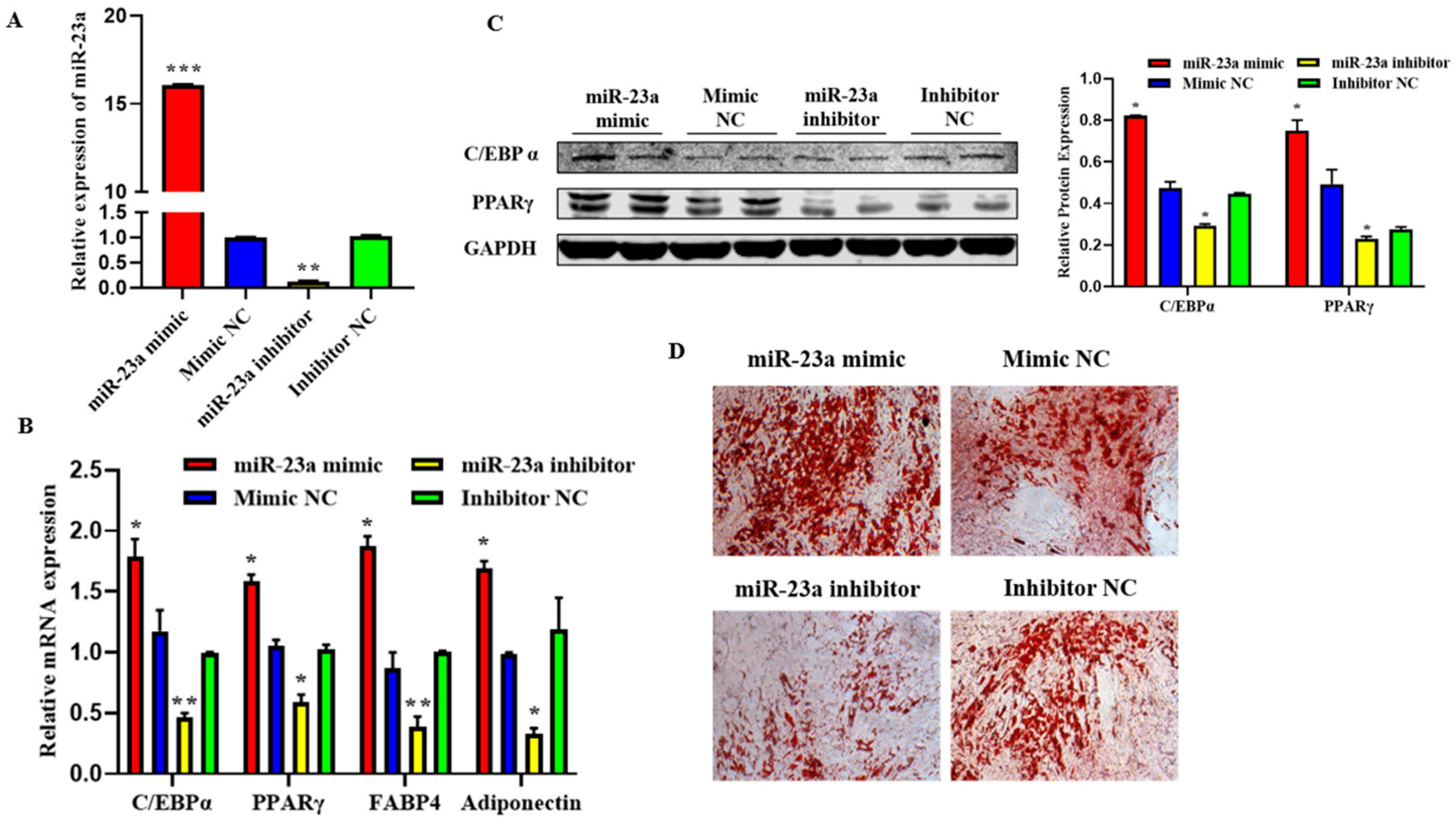
2.4. MiR-23a Inhibits Adipocytes Proliferation
2.5. MiR-23a Promotes Adipocytes Differentiation
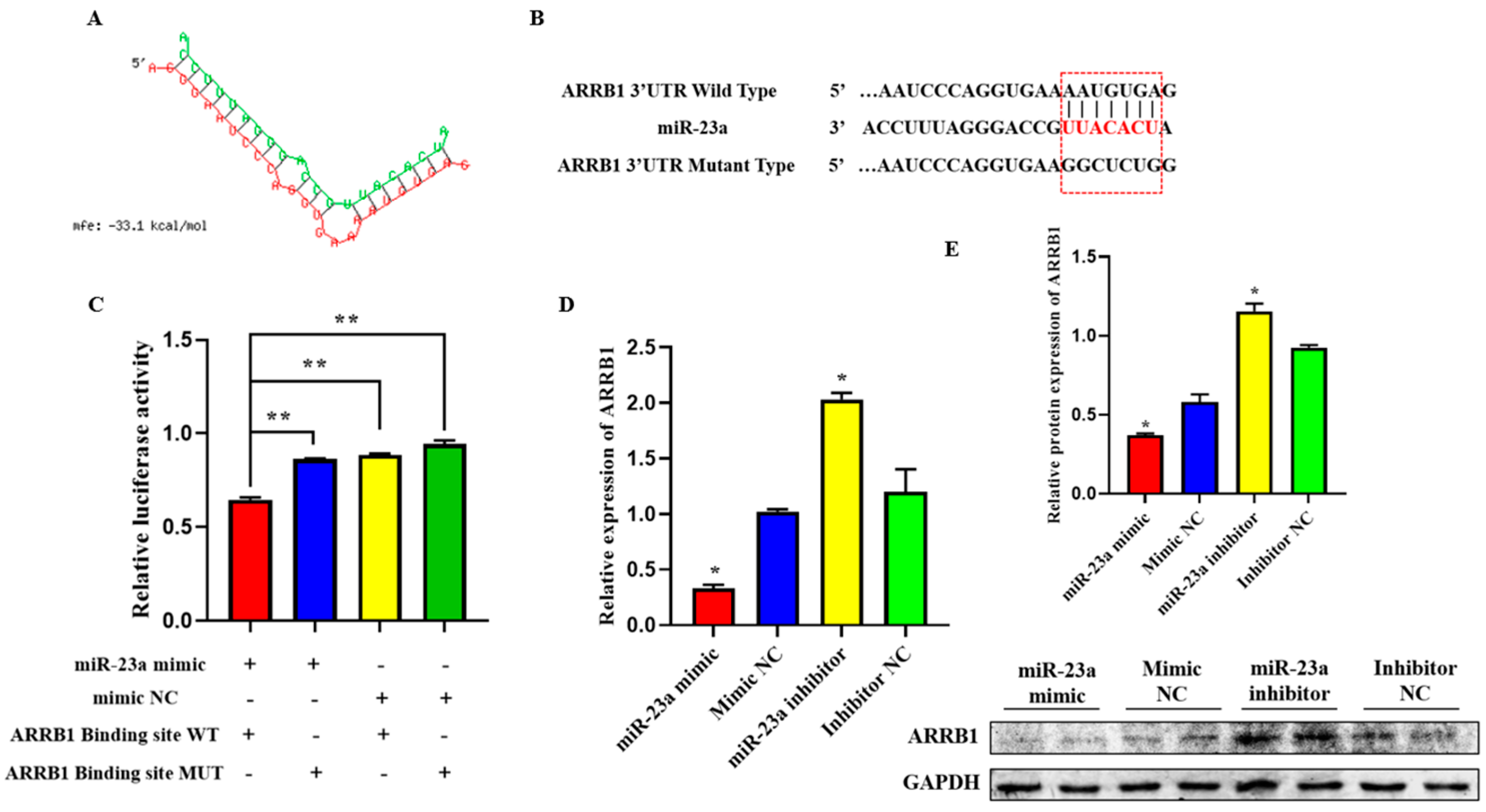
2.6. ARRB1 Is a Target of miR-23a
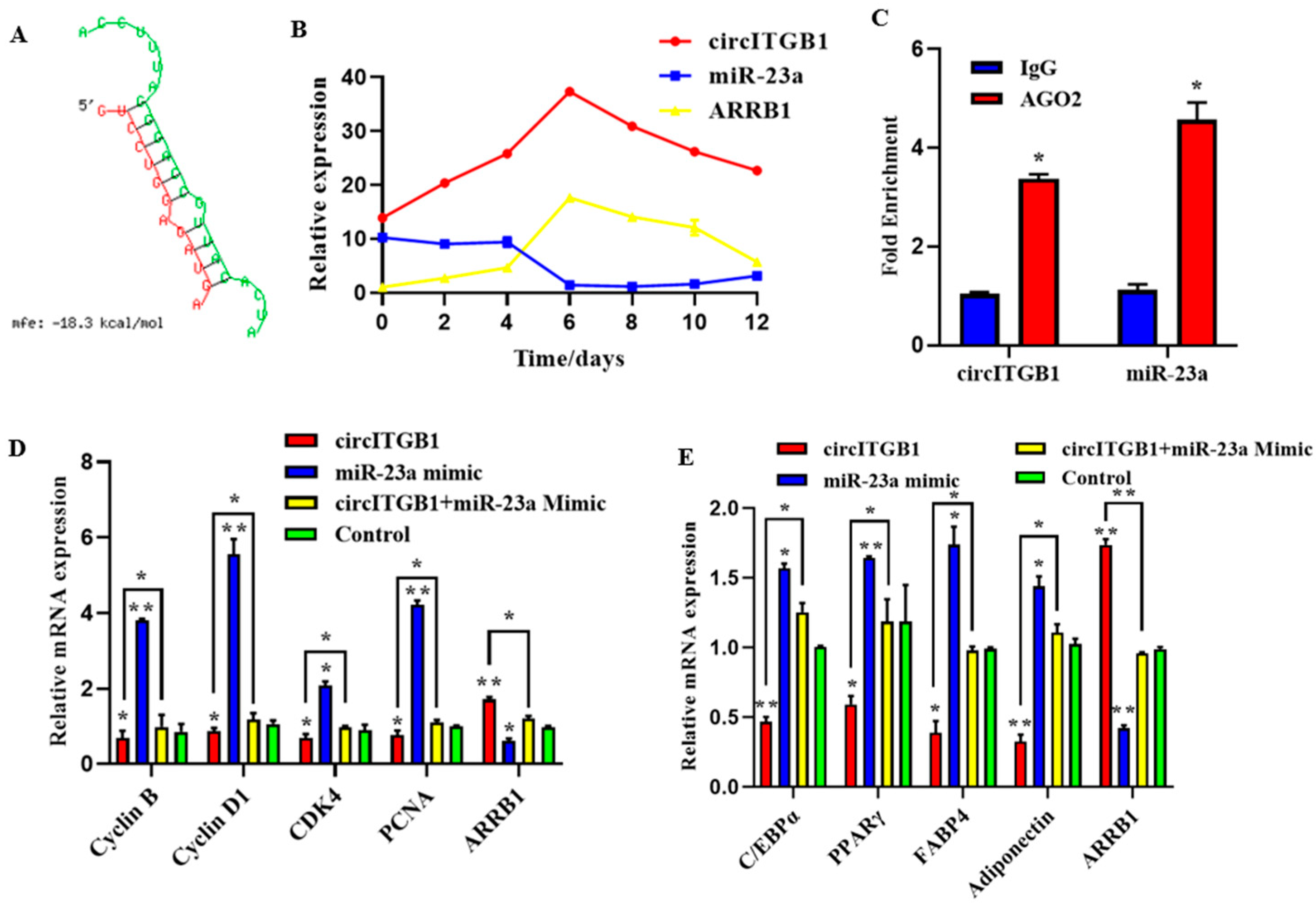
2.7. CircITGB1 Act as a ceRNA for miR-23a
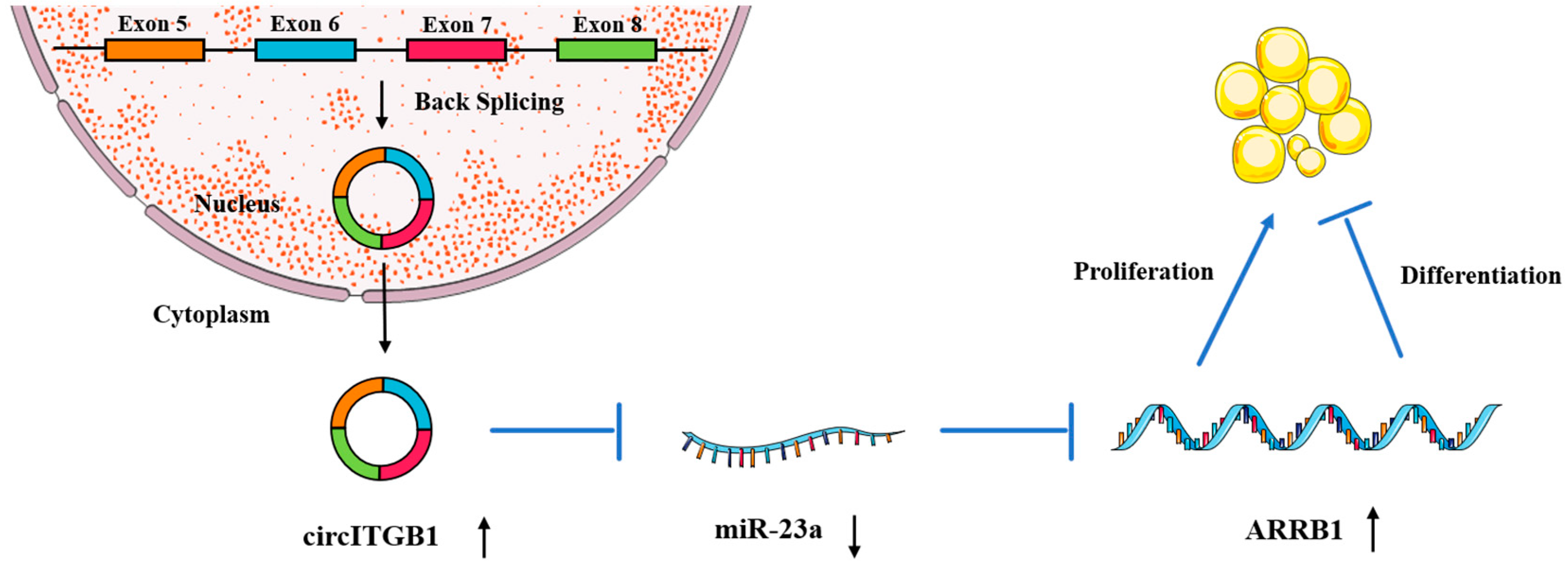
3. Discussion
4. Materials and Methods
4.1. Sample Collection
4.2. Cell Isolation and Culture
4.3. Plasmids Construction
4.4. Cell Transfection
4.5. RNA Exaction, cDNA Synthesis, and Quantitative Real-Time PCR
4.6. Primer
4.7. Validation of circITGB1
4.8. EdU and CCK-8 Analysis
4.9. Western Blotting
4.10. Oil Red Staining
4.11. Biding Relationship Prediction
4.12. Dual-Luciferase Reporter Assay
4.13. RIP Assay
4.14. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Wyatt, S.B.; Winters, K.P.; Dubbert, P.M. Overweight and obesity: Prevalence, consequences, and causes of a growing public health problem. Am. J. Med Sci. 2006, 331, 166–174. [Google Scholar] [CrossRef]
- Brown, R.E.; Sharma, A.M.; Ardern, C.I.; Mirdamadi, P.; Mirdamadi, P.; Kuk, J.L. Secular differences in the association between caloric intake, macronutrient intake, and physical activity with obesity. Obes. Res. Clin. Pract. 2016, 10, 243–255. [Google Scholar] [CrossRef]
- Mount, P.; Davies, M.; Choy, S.-W.; Cook, N.; Power, D.J.M. Obesity-related chronic kidney disease—The role of lipid metabolism. Metabolites 2015, 5, 720–732. [Google Scholar] [CrossRef]
- Toledo-Corral, C.M.; Alderete, T.L.; Hu, H.H.; Nayak, K.; Esplana, S.; Liu, T.; Goran, M.I.; Weigensberg, M.J. Metabolism, Ectopic fat deposition in prediabetic overweight and obese minority adolescents. J. Clin. Endocrinol. 2013, 98, 1115–1121. [Google Scholar] [CrossRef]
- Tedeschi, L.; Cannas, A.; Fox, D. A nutrition mathematical model to account for dietary supply and requirements of energy and other nutrients for domesticated small ruminants: The development and evaluation of the Small Ruminant Nutrition System. Small Rumin. Res. 2010, 89, 174–184. [Google Scholar] [CrossRef]
- Alberto, F.J.; Boyer, F.; Orozco-terWengel, P.; Streeter, I.; Servin, B.; de Villemereuil, P.; Benjelloun, B.; Librado, P.; Biscarini, F.; Colli, L.; et al. Convergent genomic signatures of domestication in sheep and goats. Nat. Commun. 2018, 9, 1–9. [Google Scholar] [CrossRef]
- Hammer, R.E.; Pursel, V.G.; Rexroad, C.E.; Wall, R.J.; Bolt, D.J.; Ebert, K.M.; Palmiter, R.D.; Brinster, R.L. Production of transgenic rabbits, sheep and pigs by microinjection. Nature 1985, 315, 680–683. [Google Scholar] [CrossRef]
- Shumbusho, F.; Raoul, J.; Astruc, J.-M.; Palhiere, I.; Lemarié, S.; Fugeray-Scarbel, A.; Elsen, J.M. Economic evaluation of genomic selection in small ruminants: A sheep meat breeding program. Animal 2016, 10, 1033–1041. [Google Scholar] [CrossRef]
- Wilkes, D.; Li, G.; Angeles, C.F.; Patterson, J.T.; Angeles, C.F. A large animal neuropathic pain model in sheep: A strategy for improving the predictability of preclinical models for therapeutic development. J. Pain Res. 2012, 5, 415. [Google Scholar] [CrossRef]
- Lu, M. Circular RNA: Functions, applications and prospects. ExRNA 2020, 2, 1–7. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef]
- Qu, S.; Yang, X.; Li, X.; Wang, J.; Gao, Y.; Shang, R.; Sun, W.; Dou, K.; Li, H. Circular RNA: A new star of noncoding RNAs. Cancer Lett. 2015, 365, 141–148. [Google Scholar] [CrossRef]
- Patop, I.L.; Wüst, S.; Kadener, S. Past, present, and future of circ RNA s. EMBO J. 2019, 38, e100836. [Google Scholar] [CrossRef]
- Wu, J.; Qi, X.; Liu, L.; Hu, X.; Liu, J.; Yang, J.; Yang, J.; Lu, L.; Zhang, Z.; Ma, S. Emerging epigenetic regulation of circular RNAs in human cancer. Mol. Ther.-Nucleic Acids 2019, 16, 589–596. [Google Scholar] [CrossRef]
- Huang, K.; Shi, X.E.; Wang, J.; Yao, Y.; Peng, Y.; Chen, X.; Li, X.; Yang, G. Upregulated microRNA-106a promotes porcine preadipocyte proliferation and differentiation by targeting different genes. Genes 2019, 10, 805. [Google Scholar] [CrossRef]
- Fan, Y.; Gan, M.; Tan, Y.; Chen, L.; Shen, L.; Niu, L.; Liu, Y.; Tang, G.; Jiang, Y.; Li, X. Mir-152 regulates 3T3-L1 preadipocyte proliferation and differentiation. Molecules 2019, 24, 3379. [Google Scholar] [CrossRef]
- Acharya, A.; Berry, D.C.; Zhang, H.; Jiang, Y.; Jones, B.T.; Hammer, R.E.; Graff, J.M.; Mendell, J.T. miR-26 suppresses adipocyte progenitor differentiation and fat production by targeting Fbxl19. Genes Dev. 2019, 33, 1367–1380. [Google Scholar] [CrossRef]
- Li, X.; Ballantyne, L.L.; Yu, Y.; Funk, C.D. Perivascular adipose tissue–derived extracellular vesicle miR-221-3p mediates vascular remodeling. FASEB J. 2019, 33, 12704–12722. [Google Scholar] [CrossRef]
- Chen, T.; Zhang, Y.; Liu, Y.; Zhu, D.; Yu, J.; Li, G.; Sun, Z.; Wang, W.; Jiang, H.; Hong, Z. MiR-27a promotes insulin resistance and mediates glucose metabolism by targeting PPAR-γ-mediated PI3K/AKT signaling. Aging (Albany NY) 2019, 11, 7510. [Google Scholar] [CrossRef]
- Lin, X.; Luo, J.; Zhang, L.; Zhu, J. MicroRNAs synergistically regulate milk fat synthesis in mammary gland epithelial cells of dairy goats. Gene Expr. 2013, 16, 1. [Google Scholar] [CrossRef]
- Zheng, T.; Chen, H. Resveratrol ameliorates the glucose uptake and lipid metabolism in gestational diabetes mellitus mice and insulin-resistant adipocytes via miR-23a-3p/NOV axis. Mol. Immunol. 2021, 137, 163–173. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Chen, Y.; Guo, L.; Jiang, T.; Lin, Z. miR-23a/b regulates the balance between osteoblast and adipocyte differentiation in bone marrow mesenchymal stem cells. Bone Res. 2016, 4, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Xiao, C.; Wei, T.; Liu, L.X.; Liu, J.Q.; Wang, C.X.; Yuan, Z.Y.; Ma, H.H.; Jin, H.G.; Zhang, L.C.; Cao, Y. Whole-Transcriptome analysis of preadipocyte and adipocyte and construction of regulatory networks to investigate lipid metabolism in sheep. Front. Genet. 2021, 12, 662143. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Li, H.; Yang, J.; Shen, X.; Song, C.; Yang, Z.; Wang, X.; Huang, Y.; Lan, X.; Lei, C. circRNA profiling reveals an abundant circFUT10 that promotes adipocyte proliferation and inhibits adipocyte differentiation via sponging let-7. Mol. Ther.-Nucleic Acids 2020, 20, 491–501. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, H.; Li, Y.; Mao, R.; Yang, H.; Zhang, Y.; Zhang, Y.; Guo, P.; Zhan, D.; Zhang, T. Circular RNA SAMD4A controls adipogenesis in obesity through the miR-138-5p/EZH2 axis. Theranostics 2020, 10, 4705. [Google Scholar] [CrossRef]
- Fink-Gremmels, J.; Malekinejad, H. Technology, Clinical effects and biochemical mechanisms associated with exposure to the mycoestrogen zearalenone. Anim. Feed. Sci. Technol. 2007, 137, 326–341. [Google Scholar] [CrossRef]
- Zhuang, L.-N.; Hu, W.-X.; Xin, S.-M.; Zhao, J.; Pei, G. β-arrestin-1 protein represses adipogenesis and inflammatory responses through its interaction with peroxisome proliferator-activated receptor-γ (PPARγ). J. Biol. Chem. 2011, 286, 28403–28413. [Google Scholar] [CrossRef]
- Salzman, J.; Chen, R.E.; Olsen, M.N.; Wang, P.L.; Brown, P.O. Cell-type specific features of circular RNA expression. PLoS Genet. 2013, 9, e1003777. [Google Scholar] [CrossRef]
- Liang, D.; Tatomer, D.C.; Luo, Z.; Wu, H.; Yang, L.; Chen, L.-L.; Cherry, S.; Wilusz, J.E. The output of protein-coding genes shifts to circular RNAs when the pre-mRNA processing machinery is limiting. Mol. Cell 2017, 68, 940–954.e3. [Google Scholar] [CrossRef]
- Chen, L.-L. The biogenesis and emerging roles of circular RNAs. Nat. Rev. Mol. Cell Biol. 2016, 17, 205–211. [Google Scholar] [CrossRef]
- Zhang, Y.; Xue, W.; Li, X.; Zhang, J.; Chen, S.; Zhang, J.-L.; Yang, L.; Chen, L.-L. The biogenesis of nascent circular RNAs. Cell Rep. 2016, 15, 611–624. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; You, S.; Yao, Y.; Liu, Z.-J.; Hazi, W.; Li, C.-Y.; Zhang, X.-Y.; Hou, X.-X.; Wei, J.-C.; Li, X.-Y. Expression profiles of circular RNAs in sheep skeletal muscle. Asian-Australas. J. Anim. Sci. 2018, 31, 1550. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Zhou, H.; Hickford, J.G.; Gong, H.; Wang, J.; Hu, J.; Liu, X.; Li, S.; Zhao, M.; Luo, Y. Identification and characterization of circular RNA in lactating mammary glands from two breeds of sheep with different milk production profiles using RNA-Seq. Genomics 2020, 112, 2186–2193. [Google Scholar] [CrossRef] [PubMed]
- Liu, A.; Chen, X.; Liu, M.; Zhang, L.; Ma, X.; Tian, S. Differential expression and functional analysis of CircRNA in the ovaries of low and high fecundity hanper sheep. Animals 2021, 11, 1863. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, C.; Wei, J.; Ni, W.; Xu, Y.; Yao, R.; Zhang, M.; Li, H.; Liu, L.; Dang, H. Comprehensive expression profiling analysis of pituitary indicates that circRNA participates in the regulation of sheep estrus. Genes 2019, 10, 90. [Google Scholar] [CrossRef]
- Fu, D.; Shi, Y.; Liu, J.-B.; Wu, T.-M.; Jia, C.-Y.; Yang, H.-Q.; Zhang, D.-D.; Yang, X.-L.; Wang, H.-M.; Ma, Y.-S. Targeting long non-coding RNA to therapeutically regulate gene expression in cancer. Mol. Ther.-Nucleic Acids 2020, 21, 712–724. [Google Scholar] [CrossRef]
- Zhou, C.; Duan, S.J.C. The role of long non-coding RNA NNT-AS1 in neoplastic disease. Cancers 2020, 12, 3086. [Google Scholar] [CrossRef]
- Huang, Y.; Zhang, C.; Xiong, J.; Ren, H.J.G. Diseases, Emerging important roles of circRNAs in human cancer and other diseases. Genes Dis. 2021, 8, 412–423. [Google Scholar] [CrossRef]
- Xia, L.; Song, M.; Sun, M.; Wang, F.; Yang, C. Circular RNAs as biomarkers for cancer. Circ. RNAs 2018, 1087, 171–187. [Google Scholar]
- Li, Q.; Hagberg, C.E.; Silva Cascales, H.; Lang, S.; Hyvönen, M.T.; Salehzadeh, F.; Chen, P.; Alexandersson, I.; Terezaki, E.; Harms, M.J.; et al. Obesity and hyperinsulinemia drive adipocytes to activate a cell cycle program and senesce. Nat. Med. 2021, 27, 1941–1953. [Google Scholar] [CrossRef]
- Lauvai, J.; Schumacher, M.; Finco, F.D.B.A.; Graeve, L. Bacaba phenolic extract attenuates adipogenesis by down-regulating PPARγ and C/EBPα in 3T3-L1 cells. NFS J. 2017, 9, 8–14. [Google Scholar] [CrossRef]
- Pan, Y.; Jing, J.; Qiao, L.; Liu, J.; Zhao, J.; An, L.; Li, B.; Wang, W.; Liang, C.; Liu, W. miR-124-3p affects the formation of intramuscular fat through alterations in branched chain amino acid consumption in sheep. Biochem. Biophys. Res. Commun. 2018, 495, 1769–1774. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Pan, Y.; Ren, D.; Zhao, Y.; Li, B.; Qiao, L.; Liu, W. Regulation of SCD1 gene expression by miR-200c and miR-429 in ovine subcutaneous adipocytes. Acta Vet. Et Zootech. Sin. 2019, 50, 1347–1357. [Google Scholar]
- Zhou, X.; Yang, P.-C. MicroRNA: A small molecule with a big biological impact. MicroRNA (Shariqah United Arab. Emir.) 2012, 1, 1. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Wang, L.; Raza, S.H.A.; Wang, X.; Wang, G.; Liang, C.; Cheng, G.; Li, B.; Zan, L. MiR-33a plays a crucial role in the proliferation of bovine preadipocytes. Adipocyte 2021, 10, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Dai, Y.-M.; Ji, C.-B.; Yang, L.; Shi, C.-M.; Xu, G.-F.; Pang, L.-X.; Huang, F.-Y.; Zhang, C.-M.; Guo, X.-R. MiR-146b is a regulator of human visceral preadipocyte proliferation and differentiation and its expression is altered in human obesity. Mol. Cell. Endocrinol. 2014, 393, 65–74. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Du, J.; Wang, L.; Niu, L.; Zhao, Y.; Tang, G.; Jiang, Y.; Shuai, S.; Bai, L.; Li, X. MicroRNA-143a-3p modulates preadipocyte proliferation and differentiation by targeting MAPK7. Biomed. Pharmacother. 2018, 108, 531–539. [Google Scholar] [CrossRef]
- Zhang, Z.; Wu, S.; Muhammad, S.; Ren, Q.; Sun, C. miR-103/107 promote ER stress-mediated apoptosis via targeting the Wnt3a/β-catenin/ATF6 pathway in preadipocytes. J. Lipid Res. 2018, 59, 843–853. [Google Scholar] [CrossRef]
- Du, J.; Zhang, P.; Gan, M.; Zhao, X.; Xu, Y.; Li, Q.; Jiang, Y.; Tang, G.; Li, M.; Wang, J. MicroRNA-204-5p regulates 3T3-L1 preadipocyte proliferation, apoptosis and differentiation. Gene 2018, 668, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Mirra, P.; Desiderio, A.; Spinelli, R.; Nigro, C.; Longo, M.; Parrillo, L.; D’Esposito, V.; Carissimo, A.; Hedjazifar, S.; Smith, U. Adipocyte precursor cells from first degree relatives of type 2 diabetic patients feature changes in hsa-mir-23a-5p,-193a-5p, and-193b-5p and insulin-like growth factor 2 expression. FASEB J. 2021, 35, e21357. [Google Scholar] [CrossRef]
- Wang, C.; Zeng, X.; Zhou, Z.; Zhao, J.; Pei, G. β-arrestin-1 contributes to brown fat function and directly interacts with PPARα and PPARγ. Sci. Rep. 2016, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Pan, Y.; Jing, J.; Qiao, L.; Liu, J.; An, L.; Li, B.; Ren, D.; Liu, W. MiRNA-seq reveals that miR-124-3p inhibits adipogenic differentiation of the stromal vascular fraction in sheep via targeting C/EBPα. Domest. Anim. Endocrinol. 2018, 65, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Krupp, G. Stringent RNA Quality Control Using the Agilent 2100 Bioanalyzer; Agilent Technologies: Waldbronn, Germany, 2005. [Google Scholar]
- Krüger, J.; Rehmsmeier, M. RNAhybrid: microRNA target prediction easy, fast and flexible. Nucleic Acids Res. 2006, 34 (Suppl. 2), W451–W454. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) | Length (bp) |
|---|---|---|
| Cyclin B | CGTACTCCGTCTCCAGCC | 261 |
| AGCCAGTCAATCAGGATGGC | ||
| Cyclin D1 | GATGCCAACCTCCTCAACGA | 211 |
| GGAAGCGGTCCAGGTAGTTC | ||
| PCNA | ATCAGCTCAAGTGGCGTGAA | 213 |
| TGCCAAGGTGTCCGCATTAT | ||
| CDK4 | CCAATGTTGTCCGGCTGATG | 150 |
| CCTTGATCGTTTCGGCTGG | ||
| C/EBPα | TCCGTGGACAAGAACAGCAA | 137 |
| TCATTGTCACTGGTCAGCTCC | ||
| PPARγ | CGTACTCCGTCTCCAGCC | 234 |
| AGCCAGTCAATCAGGATGGC | ||
| FABP4 | AAACTGGGATGGGAAATCAACC | 261 |
| TGCTCTCTCGTAAACTCTGGTAGC | ||
| Adiponectin | ATCCCCGGGCTGTACTACTT | 129 |
| CTGGTCCACGTTCTGGTTCT | ||
| circITGB1 | TGCGTACAATGGTTTGGCTC | 185 |
| ATGCGCTGCTTACCAACAAG | ||
| ITGB1 | GTGGATCCCTTGTCCCACTG | 158 |
| ACCACACCTGCTACAATCGG | ||
| GAPDH | ACAGTCAAGGCAGAGAACGG | 98 |
| CCAGCATCACCCCACTTGAT | ||
| U6 | AGCCTTCAAATCACTGGCTACA | 197 |
| AGTACCTGCTTACCCATACCT | ||
| miR-23a | GGCGATCACATTGCCAGGGATTTCCA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yue, X.; Fan, M.; Liang, Y.; Qiao, L.; Liu, J.; Pan, Y.; Yang, K.; Liu, W. circITGB1 Regulates Adipocyte Proliferation and Differentiation via the miR-23a/ARRB1 Pathway. Int. J. Mol. Sci. 2023, 24, 1976. https://doi.org/10.3390/ijms24031976
Yue X, Fan M, Liang Y, Qiao L, Liu J, Pan Y, Yang K, Liu W. circITGB1 Regulates Adipocyte Proliferation and Differentiation via the miR-23a/ARRB1 Pathway. International Journal of Molecular Sciences. 2023; 24(3):1976. https://doi.org/10.3390/ijms24031976
Chicago/Turabian StyleYue, Xiaoyv, Mengdan Fan, Yv Liang, Liying Qiao, Jianhua Liu, Yangyang Pan, Kaijie Yang, and Wenzhong Liu. 2023. "circITGB1 Regulates Adipocyte Proliferation and Differentiation via the miR-23a/ARRB1 Pathway" International Journal of Molecular Sciences 24, no. 3: 1976. https://doi.org/10.3390/ijms24031976
APA StyleYue, X., Fan, M., Liang, Y., Qiao, L., Liu, J., Pan, Y., Yang, K., & Liu, W. (2023). circITGB1 Regulates Adipocyte Proliferation and Differentiation via the miR-23a/ARRB1 Pathway. International Journal of Molecular Sciences, 24(3), 1976. https://doi.org/10.3390/ijms24031976