
Resveratrol Analogs and Prodrugs Differently Affect the Survival of Breast Cancer Cells Impairing Estrogen/Estrogen Receptor α/Neuroglobin Pathway

 ,
,  , , ,
, , ,  ,
,  ,
,  , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
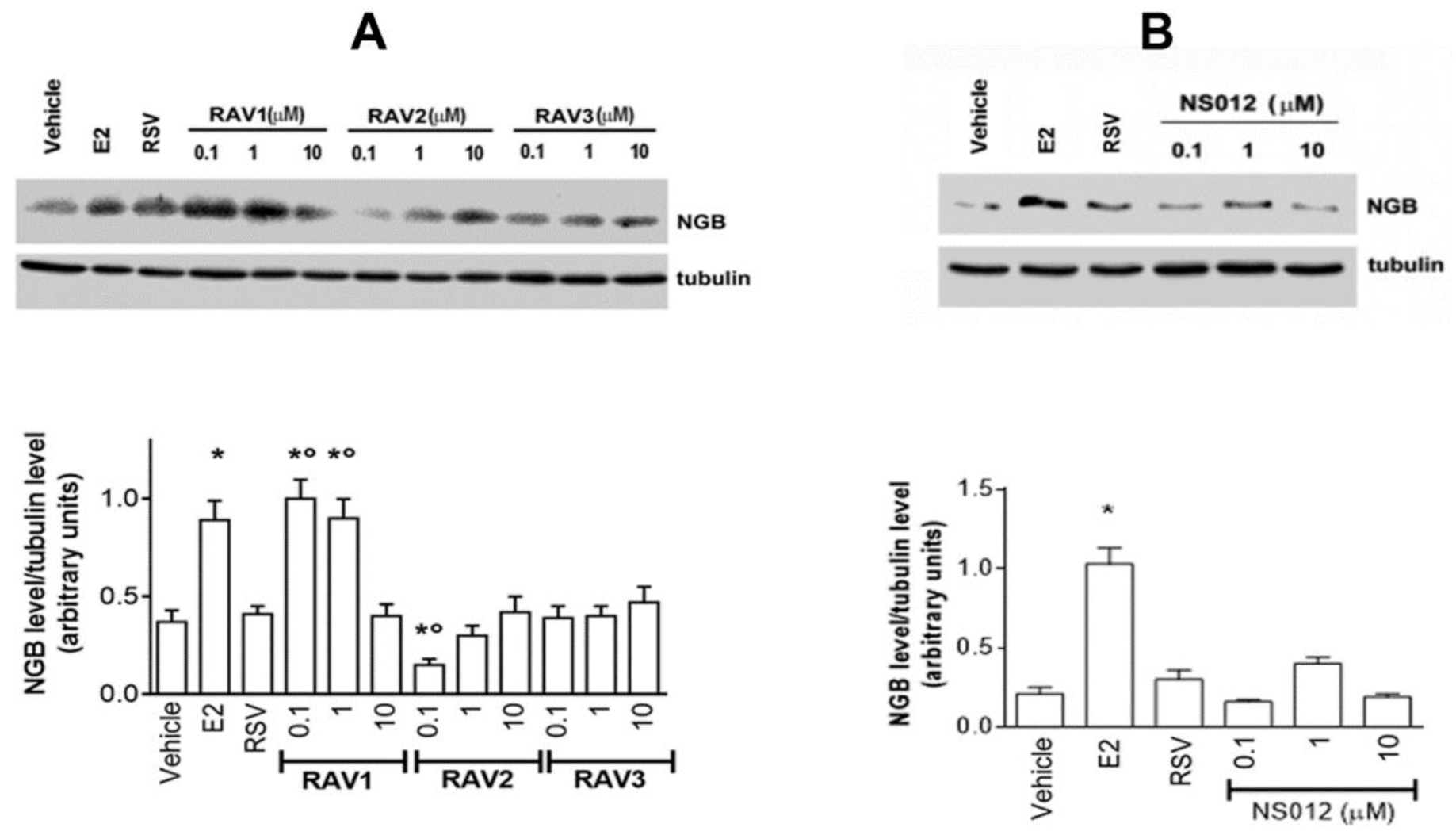
2.1. Effect of Resveratrol Analogs on NGB Levels in MCF-7 Breast Cancer Cells
2.2. Effect of Gold Nanoparticle-Conjugated RSV on NGB Levels Modulation
2.3. Involvement of ERα and ERα Signaling in NP-R Effects
2.4. Functional Outcomes of NP-R
2.5. Cellular Uptake and Internalization of NP-R
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Synthesis of RSV Analogues
4.2.1. General Procedure for the Synthesis of Derivatives RAV1, RAV2, RAV3, and NS012
4.2.2. (E)-1-(6-methoxynaphthalen-2-yl)ethan-1-one O-(benzo[d][1,3]dioxol-5-ylmethyl) Oxime (RAV1)
4.2.3. (E)-benzo[d][1,3]dioxole-4-carbaldehyde O-(benzo[d][1,3]dioxol-5-ylmethyl) Oxime (RAV2)
4.2.4. (E)-2-hydroxy-4-iodobenzaldehyde O-(benzo[d][1,3]dioxol-5-ylmethyl) Oxime (RAV3)
4.2.5. (E)-4-hydroxybenzaldehyde O-(benzo[d][1,3]dioxol-5-ylmethyl) Oxime (NS012)
4.3. Synthesis and Purification of Gold NP and NP-R
4.4. Cell Culture
4.5. Western Blot
4.6. Electron Microscopy
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA. Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Faneyte, I.F.; Schrama, J.G.; Peterse, J.L.; Remijnse, P.L.; Rodenhuis, S.; van de Vijver, M.J. Breast Cancer Response to Neoadjuvant Chemotherapy: Predictive Markers and Relation with Outcome. Br. J. Cancer 2003, 88, 406–412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tokuda, E.; Seino, Y.; Arakawa, A.; Saito, M.; Kasumi, F.; Hayashi, S.; Yamaguchi, Y. Estrogen Receptor-α Directly Regulates Sensitivity to Paclitaxel in Neoadjuvant Chemotherapy for Breast Cancer. Breast Cancer Res. Treat. 2012, 133, 427–436. [Google Scholar] [CrossRef] [PubMed]
- Conforti, R.; Boulet, T.; Tomasic, G.; Taranchon, E.; Arriagada, R.; Spielmann, M.; Ducourtieux, M.; Soria, J.C.; Tursz, T.; Delaloge, S.; et al. Breast Cancer Molecular Subclassification and Estrogen Receptor Expression to Predict Efficacy of Adjuvant Anthracyclines-Based Chemotherapy: A Biomarker Study from Two Randomized Trials. Ann. Oncol. Off. J. Eur. Soc. Med. Oncol. 2007, 18, 1477–1483. [Google Scholar] [CrossRef]
- Fiocchetti, M.; Nuzzo, M.T.; Totta, P.; Acconcia, F.; Ascenzi, P.; Marino, M. Neuroglobin, a pro-Survival Player in Estrogen Receptor α-Positive Cancer Cells. Cell Death Dis. 2014, 5, e1449. [Google Scholar] [CrossRef] [Green Version]
- Fiocchetti, M.; Cipolletti, M.; Leone, S.; Ascenzi, P.; Marino, M. Neuroglobin Overexpression Induced by the 17β-Estradiol-Estrogen Receptor-α Pathway Reduces the Sensitivity of MCF-7 Breast Cancer Cell to Paclitaxel. IUBMB Life 2016, 68, 645–651. [Google Scholar] [CrossRef]
- Shaito, A.; Posadino, A.M.; Younes, N.; Hasan, H.; Halabi, S.; Alhababi, D.; Al-Mohannadi, A.; Abdel-Rahman, W.M.; Eid, A.H.; Nasrallah, G.K.; et al. Potential Adverse Effects of Resveratrol: A Literature Review. Int. J. Mol. Sci. 2020, 21, 2084. [Google Scholar] [CrossRef] [Green Version]
- Bowers, J.L.; Tyulmenkov, V.V.; Jernigan, S.C.; Klinge, C.M. Resveratrol Acts as a Mixed Agonist/Antagonist for Estrogen Receptors Alpha and Beta. Endocrinology 2000, 141, 3657–3667. [Google Scholar] [CrossRef]
- Cipolletti, M.; Montalesi, E.; Nuzzo, M.T.; Fiocchetti, M.; Ascenzi, P.; Marino, M. Potentiation of Paclitaxel Effect by Resveratrol in Human Breast Cancer Cells by Counteracting the 17β-Estradiol/Estrogen Receptor α/Neuroglobin Pathway. J. Cell. Physiol. 2019, 234, 3147–3157. [Google Scholar] [CrossRef]
- Almeida, L.; Vaz-da-Silva, M.; Falcão, A.; Soares, E.; Costa, R.; Loureiro, A.I.; Fernandes-Lopes, C.; Rocha, J.-F.; Nunes, T.; Wright, L.; et al. Pharmacokinetic and Safety Profile of Trans-Resveratrol in a Rising Multiple-Dose Study in Healthy Volunteers. Mol. Nutr. Food Res. 2009, 53 (Suppl. 1), S7–S15. [Google Scholar] [CrossRef]
- Sergides, C.; Chirilă, M.; Silvestro, L.; Pitta, D.; Pittas, A. Bioavailability and Safety Study of Resveratrol 500 Mg Tablets in Healthy Male and Female Volunteers. Exp. Ther. Med. 2016, 11, 164–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soukup, S.T.; Helppi, J.; Müller, D.R.; Zierau, O.; Watzl, B.; Vollmer, G.; Diel, P.; Bub, A.; Kulling, S.E. Phase II Metabolism of the Soy Isoflavones Genistein and Daidzein in Humans, Rats and Mice: A Cross-Species and Sex Comparison. Arch. Toxicol. 2016, 90, 1335–1347. [Google Scholar] [CrossRef] [PubMed]
- Teng, H.; Chen, L. Polyphenols and Bioavailability: An Update. Crit. Rev. Food Sci. Nutr. 2019, 59, 2040–2051. [Google Scholar] [CrossRef] [PubMed]
- Montalesi, E.; Cipolletti, M.; Cracco, P.; Fiocchetti, M.; Marino, M. Divergent Effects of Daidzein and Its Metabolites on Estrogen-Induced Survival of Breast Cancer Cells. Cancers 2020, 12, 167. [Google Scholar] [CrossRef] [Green Version]
- Intagliata, S.; Modica, M.N.; Santagati, L.M.; Montenegro, L. Strategies to Improve Resveratrol Systemic and Topical Bioavailability: An Update. Antioxidants 2019, 8, 244. [Google Scholar] [CrossRef] [Green Version]
- Venditti, I.; Iucci, G.; Fratoddi, I.; Cipolletti, M.; Montalesi, E.; Marino, M.; Secchi, V.; Battocchio, C. Direct Conjugation of Resveratrol on Hydrophilic Gold Nanoparticles: Structural and Cytotoxic Studies for Biomedical Applications. Nanomaterials 2020, 10, 1898. [Google Scholar] [CrossRef]
- Fiocchetti, M.; Cipolletti, M.; Ascenzi, P.; Marino, M. Dissecting the 17β-Estradiol Pathways Necessary for Neuroglobin Anti-Apoptotic Activity in Breast Cancer. J. Cell. Physiol. 2018, 233, 5087–5103. [Google Scholar] [CrossRef]
- Wu, X.; Hawse, J.R.; Subramaniam, M.; Goetz, M.P.; Ingle, J.N.; Spelsberg, T.C. The Tamoxifen Metabolite, Endoxifen, Is a Potent Antiestrogen That Targets Estrogen Receptor Alpha for Degradation in Breast Cancer Cells. Cancer Res. 2009, 69, 1722–1727. [Google Scholar] [CrossRef] [Green Version]
- Marino, M.; Galluzzo, P.; Ascenzi, P. Estrogen Signaling Multiple Pathways to Impact Gene Transcription. Curr. Genomics 2006, 7, 497–508. [Google Scholar] [CrossRef] [Green Version]
- Marino, M.; Acconcia, F.; Bresciani, F.; Weisz, A.; Trentalance, A. Distinct Nongenomic Signal Transduction Pathways Controlled by 17beta-Estradiol Regulate DNA Synthesis and Cyclin D(1) Gene Transcription in HepG2 Cells. Mol. Biol. Cell 2002, 13, 3720–3729. [Google Scholar] [CrossRef]
- Ascenzi, P.; di Masi, A.; Leboffe, L.; Fiocchetti, M.; Nuzzo, M.T.; Brunori, M.; Marino, M. Neuroglobin: From Structure to Function in Health and Disease. Mol. Aspects Med. 2016, 52, 1–48. [Google Scholar] [CrossRef]
- Mitra, S.; Das, R.; Emran, T.B.; Labib, R.K.; Islam, F.; Sharma, R.; Ahmad, I.; Nainu, F.; Chidambaram, K.; Alhumaydhi, F.A.; et al. Diallyl Disulfide: A Bioactive Garlic Compound with Anticancer Potential. Front. Pharmacol. 2022, 13, 943967. [Google Scholar] [CrossRef] [PubMed]
- Küpeli Akkol, E.; Genç, Y.; Karpuz, B.; Sobarzo-Sánchez, E.; Capasso, R. Coumarins and Coumarin-Related Compounds in Pharmacotherapy of Cancer. Cancers 2020, 12, 1959. [Google Scholar] [CrossRef] [PubMed]
- Virgili, F.; Marino, M. Regulation of Cellular Signals from Nutritional Molecules: A Specific Role for Phytochemicals, beyond Antioxidant Activity. Free Radic. Biol. Med. 2008, 45, 1205–1216. [Google Scholar] [CrossRef] [PubMed]
- Alamolhodaei, N.S.; Tsatsakis, A.M.; Ramezani, M.; Hayes, A.W.; Karimi, G. Resveratrol as MDR Reversion Molecule in Breast Cancer: An Overview. Food Chem. Toxicol. 2017, 103, 223–232. [Google Scholar] [CrossRef] [Green Version]
- Bishayee, A.; Politis, T.; Darvesh, A.S. Resveratrol in the Chemoprevention and Treatment of Hepatocellular Carcinoma. Cancer Treat. Rev. 2010, 36, 43–53. [Google Scholar] [CrossRef]
- Goswami, S.K.; Das, D.K. Resveratrol and Chemoprevention. Cancer Lett. 2009, 284, 1–6. [Google Scholar] [CrossRef]
- Kundu, J.K.; Surh, Y.-J. Cancer Chemopreventive and Therapeutic Potential of Resveratrol: Mechanistic Perspectives. Cancer Lett. 2008, 269, 243–261. [Google Scholar] [CrossRef]
- Ciccone, L.; Nencetti, S.; Socci, S.; Orlandini, E. Neuroglobin and Neuroprotection: The Role of Natural and Synthetic Compounds in Neuroglobin Pharmacological Induction. Neural Regen. Res. 2021, 16, 2353–2358. [Google Scholar] [CrossRef]
- Ramírez-Garza, S.L.; Laveriano-Santos, E.P.; Marhuenda-Muñoz, M.; Storniolo, C.E.; Tresserra-Rimbau, A.; Vallverdú-Queralt, A.; Lamuela-Raventós, R.M. Health Effects of Resveratrol: Results from Human Intervention Trials. Nutrients 2018, 10, 1892. [Google Scholar] [CrossRef]
- Cory, H.; Passarelli, S.; Szeto, J.; Tamez, M.; Mattei, J. The Role of Polyphenols in Human Health and Food Systems: A Mini-Review. Front. Nutr. 2018, 5, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fulda, S.; Debatin, K.-M. Sensitization for Anticancer Drug-Induced Apoptosis by the Chemopreventive Agent Resveratrol. Oncogene 2004, 23, 6702–6711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Lorenzo, C.; Colombo, F.; Biella, S.; Stockley, C.; Restani, P. Polyphenols and Human Health: The Role of Bioavailability. Nutrients 2021, 13, 273. [Google Scholar] [CrossRef] [PubMed]
- Bohara, R.A.; Tabassum, N.; Singh, M.P.; Gigli, G.; Ragusa, A.; Leporatti, S. Recent Overview of Resveratrol’s Beneficial Effects and Its Nano-Delivery Systems. Molecules 2022, 27, 5154. [Google Scholar] [CrossRef] [PubMed]
- Savjani, K.T.; Gajjar, A.K.; Savjani, J.K. Drug Solubility: Importance and Enhancement Techniques. ISRN Pharm. 2012, 2012, 195727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pecyna, P.; Wargula, J.; Murias, M.; Kucinska, M. More Than Resveratrol: New Insights into Stilbene-Based Compounds. Biomolecules 2020, 10, 1111. [Google Scholar] [CrossRef]
- Chen, Y.; Ke, G.; Ma, Y.; Zhu, Z.; Liu, M.; Liu, Y.; Yan, H.; Yang, C.J. A Synthetic Light-Driven Substrate Channeling System for Precise Regulation of Enzyme Cascade Activity Based on DNA Origami. J. Am. Chem. Soc. 2018, 140, 8990–8996. [Google Scholar] [CrossRef]
- Carr, J.A.; Wang, H.; Abraham, A.; Gullion, T.; Lewis, J.P. L-Cysteine Interaction with Au55 Nanoparticle. J. Phys. Chem. C 2012, 116, 25816–25823. [Google Scholar] [CrossRef]
- Sarangi, S.N.; Hussain, A.M.P.; Sahu, S.N. Strong UV Absorption and Emission from L-Cysteine Capped Monodispersed Gold Nanoparticles. Appl. Phys. Lett. 2009, 95, 073109. [Google Scholar] [CrossRef]
- Bertelà, F.; Marsotto, M.; Meneghini, C.; Burratti, L.; Maraloiu, V.-A.; Iucci, G.; Venditti, I.; Prosposito, P.; D’Ezio, V.; Persichini, T.; et al. Biocompatible Silver Nanoparticles: Study of the Chemical and Molecular Structure, and the Ability to Interact with Cadmium and Arsenic in Water and Biological Properties. Nanomaterials 2021, 11, 2540. [Google Scholar] [CrossRef]
- Krieg, A.J.; Krieg, S.A.; Ahn, B.S.; Shapiro, D.J. Interplay between Estrogen Response Element Sequence and Ligands Controls in Vivo Binding of Estrogen Receptor to Regulated Genes. J. Biol. Chem. 2004, 279, 5025–5034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augereau, P.; Miralles, F.; Cavaillès, V.; Gaudelet, C.; Parker, M.; Rochefort, H. Characterization of the Proximal Estrogen-Responsive Element of Human Cathepsin D Gene. Mol. Endocrinol. Baltim. Md 1994, 8, 693–703. [Google Scholar] [CrossRef] [Green Version]
- Cipolletti, M.; Solar Fernandez, V.; Montalesi, E.; Marino, M.; Fiocchetti, M. Beyond the Antioxidant Activity of Dietary Polyphenols in Cancer: The Modulation of Estrogen Receptors (ERs) Signaling. Int. J. Mol. Sci. 2018, 19, 2624. [Google Scholar] [CrossRef] [Green Version]
- Li, S.; Penninckx, S.; Karmani, L.; Heuskin, A.-C.; Watillon, K.; Marega, R.; Zola, J.; Corvaglia, V.; Genard, G.; Gallez, B.; et al. LET-Dependent Radiosensitization Effects of Gold Nanoparticles for Proton Irradiation. Nanotechnology 2016, 27, 455101. [Google Scholar] [CrossRef]
- Tremi, I.; Havaki, S.; Georgitsopoulou, S.; Lagopati, N.; Georgakilas, V.; Gorgoulis, V.G.; Georgakilas, A.G. A Guide for Using Transmission Electron Microscopy for Studying the Radiosensitizing Effects of Gold Nanoparticles In Vitro. Nanomaterials 2021, 11, 859. [Google Scholar] [CrossRef]
- Wu, Y.; Zhang, Q.; Ruan, Z.; Yin, Y. Intrinsic Effects of Gold Nanoparticles on Proliferation and Invasion Activity in SGC-7901 Cells. Oncol. Rep. 2016, 35, 1457–1462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, J.; Feliciano, T.J.; Li, W.; Lee, A.; Odom, T.W. Gold Nanoparticle Size and Shape Effects on Cellular Uptake and Intracellular Distribution of SiRNA Nanoconstructs. Bioconjug. Chem. 2017, 28, 1791–1800. [Google Scholar] [CrossRef]
- Balsamo, A.; Coletta, I.; Domiano, P.; Guglielmotti, A.; Landolfi, C.; Mancini, F.; Milanese, C.; Orlandini, E.; Rapposelli, S.; Pinza, M.; et al. (E)-[2-(4-Methylsulphonylphenyl)-1-Cyclopentenyl-1-Methyliden](Arylmethyloxy)Amines. Methyleneaminoxymethyl (MAOM) Analogues of Diarylcyclopentenyl Cyclooxygenase-2 Inhibitors: Synthesis and Biological Properties. Eur. J. Med. Chem. 2002, 37, 391–398. [Google Scholar] [CrossRef]
- Ciccone, L.; Nencetti, S.; Tonali, N.; Fruchart-Gaillard, C.; Shepard, W.; Nuti, E.; Camodeca, C.; Rossello, A.; Orlandini, E. Monoaryl derivatives as transthyretin fibril formation inhibitors: Design, synthesis, biological evaluation and structural analysis. Bioorg. Med. Chem. 2020, 28, 115673. [Google Scholar] [CrossRef]
- Ciccone, L.; Petrarolo, G.; Barsuglia, F.; Fruchart-Gaillard, C.; Cassar Lajeunesse, E.; Adewumi, A.T.; Soliman, M.E.S.; La Motta, C.; Orlandini, E.; Nencetti, S. Nature-Inspired O-Benzyl Oxime-Based Derivatives as New Dual-Acting Agents Tar-geting Aldose Reductase and Oxidative Stress. Biomolecules 2022, 12, 44847. [Google Scholar] [CrossRef]
- Ciccone, L.; Piragine, E.; Brogi, S.; Camodeca, C.; Fucci, R.; Calderone, V.; Nencetti, S.; Martelli, A.; Orlandini, E. Resveratrol-like Compounds as SIRT1 Activators. Int. J. Mol. Sci. 2022, 23, 15105. [Google Scholar] [CrossRef] [PubMed]
- Venditti, I.; Testa, G.; Sciubba, F.; Carlini, L.; Porcaro, F.; Meneghini, C.; Mobilio, S.; Battocchio, C.; Fratoddi, I. Hydrophilic Metal Nanoparticles Functionalized by 2-Diethylaminoethanethiol: A Close Look at the Metal–Ligand Interaction and Interface Chemical Structure. J. Phys. Chem. C 2017, 121, 8002–8013. [Google Scholar] [CrossRef]
- De Marinis, E.; Ascenzi, P.; Pellegrini, M.; Galluzzo, P.; Bulzomi, P.; Arevalo, M.A.; Garcia-Segura, L.M.; Marino, M. 17β-Estradiol--a New Modulator of Neuroglobin Levels in Neurons: Role in Neuroprotection against H2O2-Induced Toxicity. Neurosignals 2010, 18, 223–235. [Google Scholar] [CrossRef] [PubMed]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Montalesi, E.; Cracco, P.; Acconcia, F.; Fiocchetti, M.; Iucci, G.; Battocchio, C.; Orlandini, E.; Ciccone, L.; Nencetti, S.; Muzzi, M.; et al. Resveratrol Analogs and Prodrugs Differently Affect the Survival of Breast Cancer Cells Impairing Estrogen/Estrogen Receptor α/Neuroglobin Pathway. Int. J. Mol. Sci. 2023, 24, 2148. https://doi.org/10.3390/ijms24032148
Montalesi E, Cracco P, Acconcia F, Fiocchetti M, Iucci G, Battocchio C, Orlandini E, Ciccone L, Nencetti S, Muzzi M, et al. Resveratrol Analogs and Prodrugs Differently Affect the Survival of Breast Cancer Cells Impairing Estrogen/Estrogen Receptor α/Neuroglobin Pathway. International Journal of Molecular Sciences. 2023; 24(3):2148. https://doi.org/10.3390/ijms24032148
Chicago/Turabian StyleMontalesi, Emiliano, Patrizio Cracco, Filippo Acconcia, Marco Fiocchetti, Giovanna Iucci, Chiara Battocchio, Elisabetta Orlandini, Lidia Ciccone, Susanna Nencetti, Maurizio Muzzi, and et al. 2023. "Resveratrol Analogs and Prodrugs Differently Affect the Survival of Breast Cancer Cells Impairing Estrogen/Estrogen Receptor α/Neuroglobin Pathway" International Journal of Molecular Sciences 24, no. 3: 2148. https://doi.org/10.3390/ijms24032148