
Transglutaminase Activity Is Conserved in Stratified Epithelia and Skin Appendages of Mammals and Birds
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
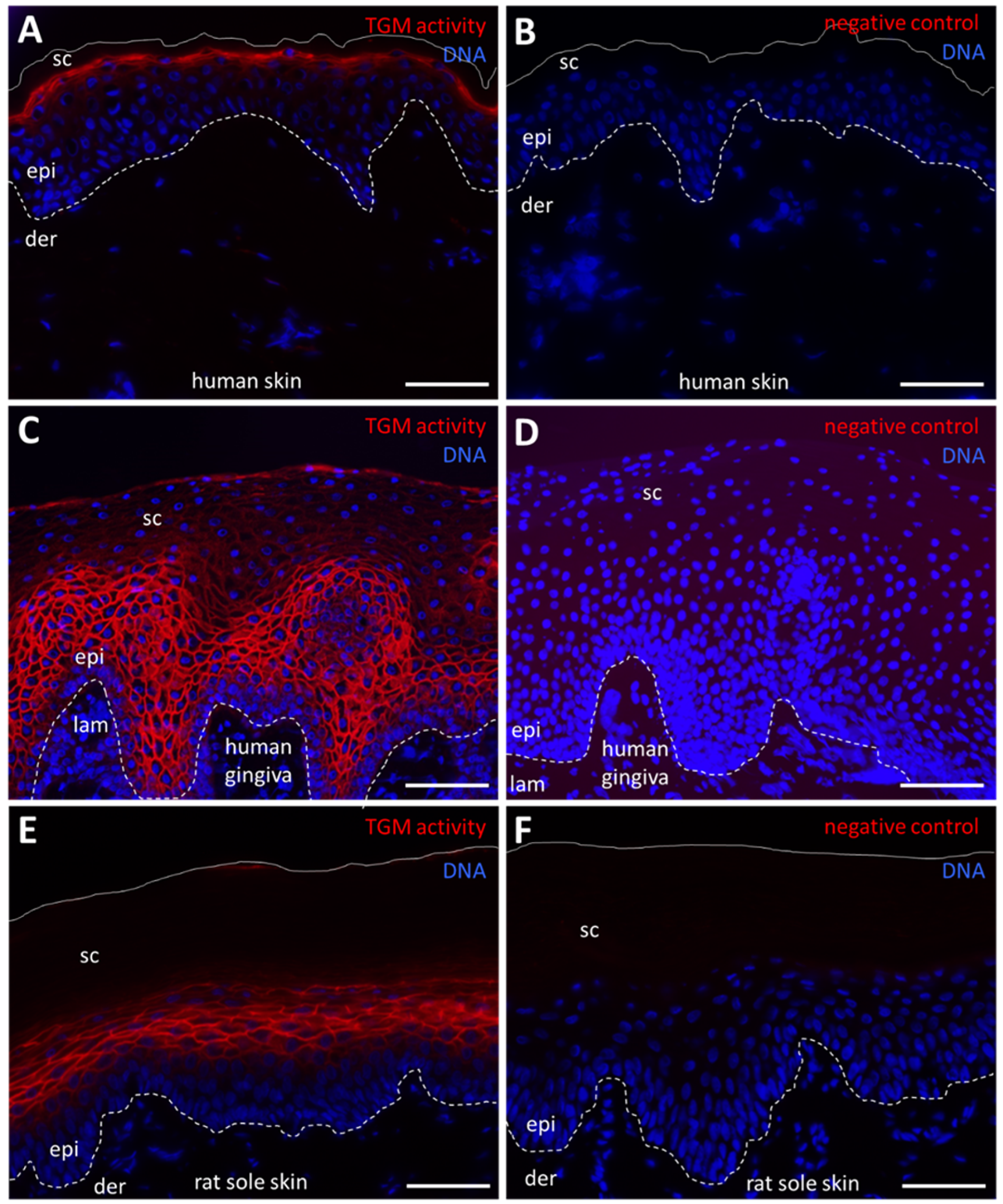
2.1. Fluorescence Labeling Detects TGM Activity in Terminally Differentiated Keratinocytes of the Epidermis and Oral Epithelium
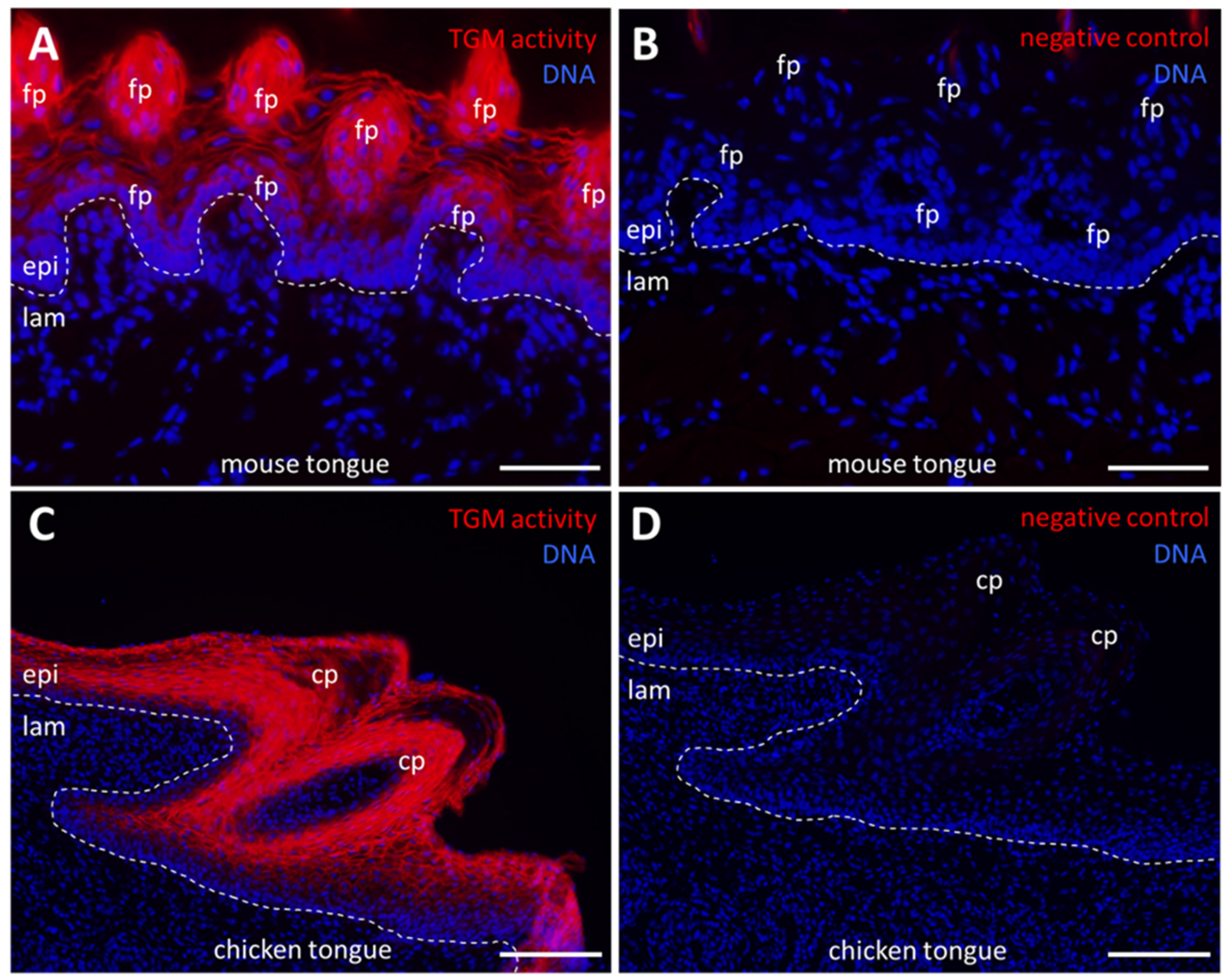
2.2. Cornifying Papillae of the Tongues of Mammals and Birds Contain Active Transglutaminases
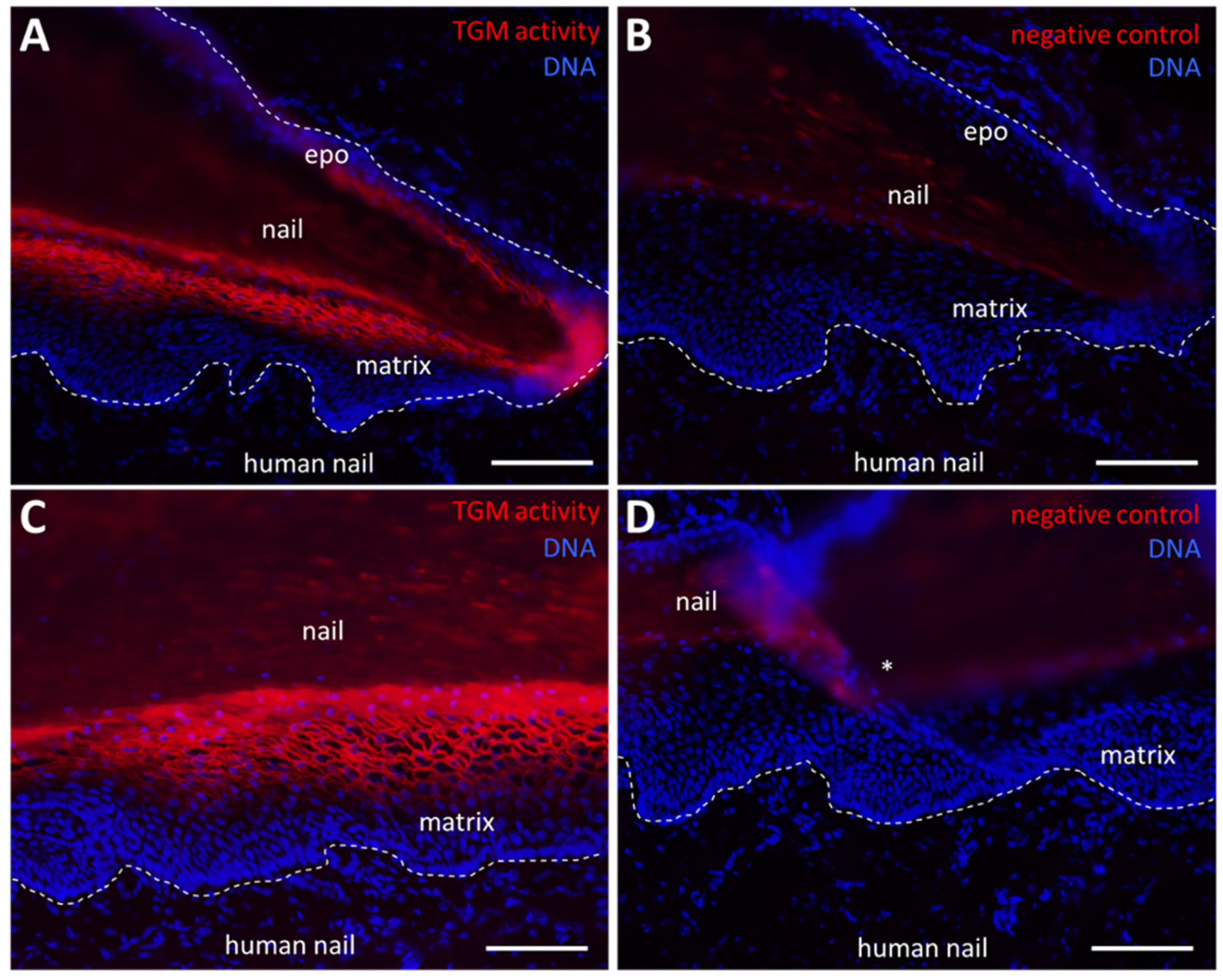
2.3. TGM Activity Is Present in the Nail Matrix
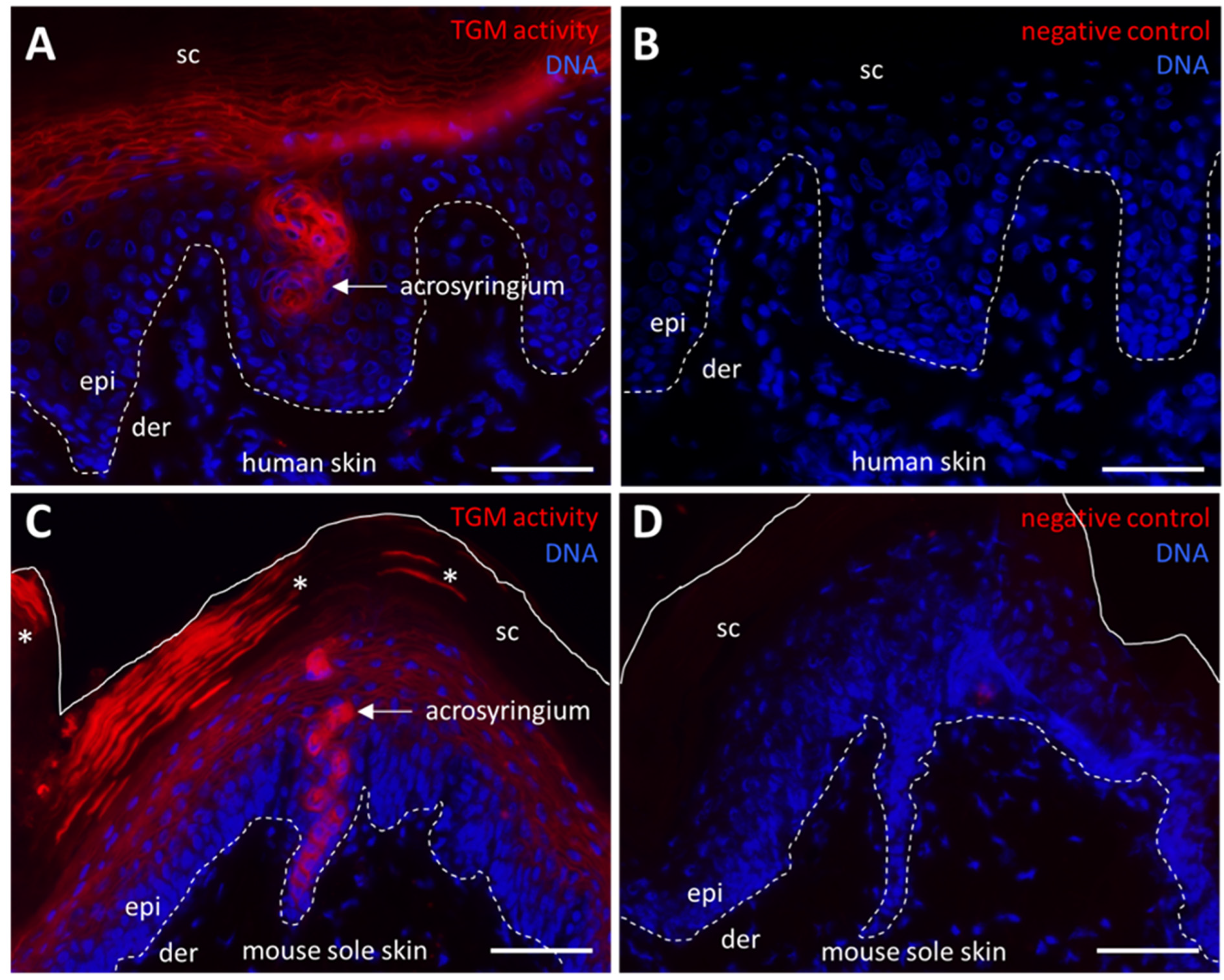
2.4. TGM Activity Contributes to Formation of the Arcosyringeal Duct of Sweat Glands
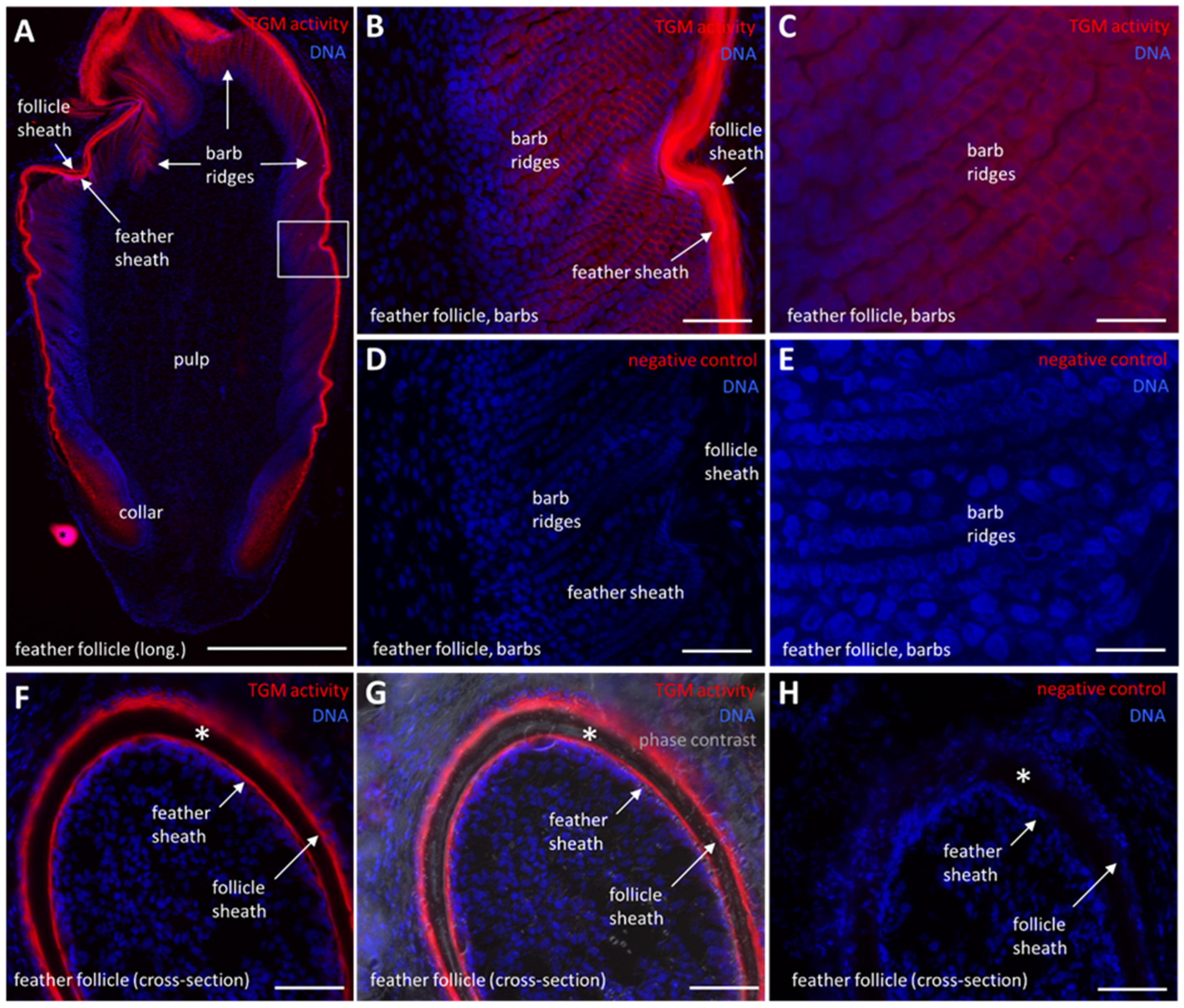
2.5. TGM Activity Is Present in Differentiated Keratinocytes of the Feather Follicle
3. Discussion
4. Materials and Methods
4.1. Tissue Samples
4.2. TGM Activity Labeling In Situ
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bereiter-Hahn, J.; Matoltsy, A.G.; Richards, K.S. (Eds.) Biology of the Integument. Vol 2. Vertebrates; Springer: New York, NY, USA, 1986. [Google Scholar]
- Chuong, C.M.; Nickoloff, B.J.; Elias, P.M.; Goldsmith, L.A.; Macher, E.; Maderson, P.A.; Sundberg, J.P.; Tagami, H.; Plonka, P.M.; Thestrup-Pederson, K.; et al. What is the ‘true’ function of skin? Exp. Dermatol. 2002, 11, 159–187. [Google Scholar] [CrossRef] [PubMed]
- Candi, E.; Schmidt, R.; Melino, G. The cornified envelope: A model of cell death in the skin. Nat. Rev. Mol. Cell. Biol. 2005, 6, 328–340. [Google Scholar] [CrossRef] [PubMed]
- Eckhart, L.; Lippens, S.; Tschachler, E.; Declercq, W. Cell death by cornification. Biochim. Biophys. Acta 2013, 1833, 3471–3480. [Google Scholar] [CrossRef] [PubMed]
- Harland, D.P.; Plowman, J.E. Development of hair fibres. Adv. Exp. Med. Biol. 2018, 1054, 109–154. [Google Scholar] [CrossRef] [PubMed]
- Ishitsuka, Y.; Roop, D.R. The epidermis: Redox governor of health and diseases. Antioxidants 2021, 11, 47. [Google Scholar] [CrossRef] [PubMed]
- Alibardi, L. Vertebrate keratinization evolved into cornification mainly due to transglutaminase and sulfhydryl oxidase activities on epidermal proteins: An immunohistochemical survey. Anat. Rec. 2022, 305, 333–358. [Google Scholar] [CrossRef] [PubMed]
- Russell, L.J.; DiGiovanna, J.J.; Rogers, G.R.; Steinert, P.M.; Hashem, N.; Compton, J.G.; Bale, S.J. Mutations in the gene for transglutaminase 1 in autosomal recessive lamellar ichthyosis. Nat. Genet. 1995, 9, 279–283. [Google Scholar] [CrossRef]
- Basmanav, F.B.Ü.; Cau, L.; Tafazzoli, A.; Méchin, M.C.; Wolf, S.; Romano, M.T.; Valentin, F.; Wiegmann, H.; Huchenq, A.; Kandil, R.; et al. Mutations in three genes encoding proteins involved in hair shaft formation cause uncombable hair syndrome. Am. J. Hum. Genet. 2016, 99, 1292–1304. [Google Scholar] [CrossRef] [Green Version]
- Cassidy, A.J.; van Steensel, M.A.; Steijlen, P.M.; van Geel, M.; van der Velden, J.; Morley, S.M.; Terrinoni, A.; Melino, G.; Candi, E.; McLean, W.H. A homozygous missense mutation in TGM5 abolishes epidermal transglutaminase 5 activity and causes acral peeling skin syndrome. Am. J. Hum. Genet. 2005, 77, 909–917. [Google Scholar] [CrossRef] [Green Version]
- Sachslehner, A.P.; Surbek, M.; Lachner, J.; Paudel, S.; Eckhart, L. Identification of chicken transglutaminase 1 and in situ localization of transglutaminase activity in avian skin and esophagus. Genes 2021, 12, 1565. [Google Scholar] [CrossRef]
- Hohl, D.; Aeschlimann, D.; Huber, M. In vitro and rapid in situ transglutaminase assays for congenital ichthyoses--a comparative study. J. Investig. Dermatol. 1998, 110, 268–271. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghunath, M.; Hennies, H.C.; Velten, F.; Wiebe, V.; Steinert, P.M.; Reis, A.; Traupe, H. A novel in situ method for the detection of deficient transglutaminase activity in the skin. Arch. Dermatol. Res. 1998, 290, 621–627. [Google Scholar] [CrossRef] [PubMed]
- Thacher, S.M.; Rice, R.H. Keratinocyte-specific transglutaminase of cultured human epidermal cells: Relation to cross-linked envelope formation and terminal differentiation. Cell 1985, 40, 685–695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dabija-Wolter, G.; Bakken, V.; Cimpan, M.R.; Johannessen, A.C.; Costea, D.E. In vitro reconstruction of human junctional and sulcular epithelium. J. Oral Pathol. Med. 2013, 42, 396–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jäger, K.; Fischer, H.; Tschachler, E.; Eckhart, L. Terminal differentiation of nail matrix keratinocytes involves up-regulation of DNase1L2 but is independent of caspase-14 expression. Differentiation 2007, 75, 939–946. [Google Scholar] [CrossRef]
- Ta, B.M.; Gallagher, G.T.; Chakravarty, R.; Rice, R.H. Keratinocyte transglutaminase in human skin and oral mucosa: Cytoplasmic localization and uncoupling of differentiation markers. J. Cell Sci. 1990, 95, 631–638. [Google Scholar] [CrossRef]
- Pereira, D.; Sequeira, I. A scarless healing tale: Comparing homeostasis and wound healing of oral mucosa with skin and oesophagus. Front. Cell. Dev. Biol. 2021, 9, 682143. [Google Scholar] [CrossRef]
- Skieresz-Szewczyk, K.; Jackowiak, H.; Skrzypski, M. Alpha-keratin, keratin-associated proteins and transglutaminase 1 are present in the ortho- and parakeratinized epithelium of the avian tongue. Cells 2022, 11, 1899. [Google Scholar] [CrossRef]
- Dhouailly, D.; Xu, C.; Manabe, M.; Schermer, A.; Sun, T.T. Expression of hair-related keratins in a soft epithelium: Subpopulations of human and mouse dorsal tongue keratinocytes express keratin markers for hair-, skin- and esophageal-types of differentiation. Exp. Cell Res. 1989, 181, 141–158. [Google Scholar] [CrossRef]
- Eckhart, L.; Ehrlich, F. Evolution of trichocyte keratins. Adv. Exp. Med. Biol. 2018, 1054, 33–45. [Google Scholar] [CrossRef]
- Mlitz, V.; Strasser, B.; Jaeger, K.; Hermann, M.; Ghannadan, M.; Buchberger, M.; Alibardi, L.; Tschachler, E.; Eckhart, L. Trichohyalin-like proteins have evolutionarily conserved roles in the morphogenesis of skin appendages. J. Investig. Dermatol. 2014, 134, 2685–2692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, H.; Szabo, S.; Scherz, J.; Jaeger, K.; Rossiter, H.; Buchberger, M.; Ghannadan, M.; Hermann, M.; Theussl, H.C.; Tobin, D.J.; et al. Essential role of the keratinocyte-specific endonuclease DNase1L2 in the removal of nuclear DNA from hair and nails. J. Investig. Dermatol. 2011, 131, 1208–1215. [Google Scholar] [CrossRef] [Green Version]
- Rice, R.H.; Crumrine, D.; Hohl, D.; Munro, C.S.; Elias, P.M. Cross-linked envelopes in nail plate in lamellar ichthyosis. Br. J. Dermatol. 2003, 149, 1050–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rice, R.H.; Crumrine, D.; Uchida, Y.; Gruber, R.; Elias, P.M. Structural changes in epidermal scale and appendages as indicators of defective TGM1 activity. Arch. Dermatol. Res. 2005, 297, 127–133. [Google Scholar] [CrossRef] [PubMed]
- Rice, R.H.; Wong, V.J.; Pinkerton, K.E. Ultrastructural visualization of cross-linked protein features in epidermal appendages. J. Cell Sci. 1994, 107, 1985–1992. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.H.; Yu, M.; Wu, P.; Jiang, T.X.; Yu, H.S.; Widelitz, R.B.; Chuong, C.M. Sculpting skin appendages out of epidermal layers via temporally and spatially regulated apoptotic events. J. Investig. Dermatol. 2004, 122, 1348–1355. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, W.L.; Wu, H.; Chiu, Y.K.; Wang, S.; Jiang, T.X.; Luo, Z.L.; Lin, Y.C.; Li, A.; Hsu, J.T.; Huang, H.L.; et al. The making of a flight feather: Bio-architectural principles and adaptation. Cell 2019, 179, 1409–1423.e17. [Google Scholar] [CrossRef]
- Prum, R.O. Development and evolutionary origin of feathers. J. Exp. Zool. 1999, 285, 291–306. [Google Scholar] [CrossRef]
- Sawyer, R.H.; Rogers, L.; Washington, L.; Glenn, T.C.; Knapp, L.W. Evolutionary origin of the feather epidermis. Dev. Dyn. 2005, 232, 256–267. [Google Scholar] [CrossRef]
- Wagner, G.P. Homology, Genes and Evolutionary Innovation; Princeton University Press: Princeton, NJ, USA, 2014. [Google Scholar]
- Alibardi, L. Review: Cornification, morphogenesis and evolution of feathers. Protoplasma 2017, 254, 1259–1281. [Google Scholar] [CrossRef]
- Strasser, B.; Mlitz, V.; Hermann, M.; Rice, R.H.; Eigenheer, R.A.; Alibardi, L.; Tschachler, E.; Eckhart, L. Evolutionary origin and diversification of epidermal barrier proteins in amniotes. Mol. Biol. Evol. 2014, 31, 3194–3205. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehrlich, F.; Lachner, J.; Hermann, M.; Tschachler, E.; Eckhart, L. Convergent evolution of cysteine-rich keratins in hard skin appendages of terrestrial vertebrates. Mol. Biol. Evol. 2020, 37, 982–993. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murphy, G.F.; Flynn, T.C.; Rice, R.H.; Pinkus, G.S. Involucrin expression in normal and neoplastic human skin: A marker for keratinocyte differentiation. J. Investig. Dermatol. 1984, 82, 453–457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ishida-Yamamoto, A.; Hohl, D.; Roop, D.R.; Iizuka, H.; Eady, R.A.J. Loricrin immunoreactivity in human skin: Localization to specific granules (L-granules) in acrosyringia. Arch. Dermatol. Res. 1993, 285, 491–498. [Google Scholar] [CrossRef]
- Rerknimitr, P.; Tanizaki, H.; Yamamoto, Y.; Amano, W.; Nakajima, S.; Nakashima, C.; Nonomura, Y.; Wititsuwannakul, J.; Miyachi, Y.; Otsuka, A.; et al. Decreased filaggrin level may lead to sweat duct obstruction in filaggrin mutant mice. J. Investig. Dermatol. 2017, 137, 248–251. [Google Scholar] [CrossRef] [Green Version]
- Takeda, M.; Nomura, T.; Sugiyama, T.; Miyauchi, T.; Suzuki, S.; Fujita, Y.; Shimizu, H. Compound heterozygous missense mutations p.Leu207Pro and p.Tyr544Cys in TGM1 cause a severe form of lamellar ichthyosis. J. Dermatol. 2018, 45, 1463–1467. [Google Scholar] [CrossRef]
- Freedman, J.C.; Parry, T.J.; Zhang, P.; Majumdar, A.; Krishnan, S.; Regula, L.K.; O’Malley, M.; Coghlan, S.; Yogesha, S.D.; Ramasamy, S.; et al. Preclinical evaluation of a modified herpes simplex virus type 1 vector encoding human TGM1 for the treatment of autosomal recessive congenital ichthyosis. J. Investig. Dermatol. 2021, 141, 874–882.e6. [Google Scholar] [CrossRef]
- Basset, J.; Marchal, L.; Hovnanian, A. EGFR signaling is overactive in pachyonychia congenita: Effective treatment with oral erlotinib. J. Investig. Dermatol. 2022, 143, 294–304.e8. [Google Scholar] [CrossRef]
- Schuppan, D.; Mäki, M.; Lundin, K.E.A.; Isola, J.; Friesing-Sosnik, T.; Taavela, J.; Popp, A.; Koskenpato, J.; Langhorst, J.; Hovde, Ø.; et al. A randomized trial of a transglutaminase 2 inhibitor for celiac disease. N. Engl. J. Med. 2021, 385, 35–45. [Google Scholar] [CrossRef]
- Paolella, G.; Sposito, S.; Romanelli, A.M.; Caputo, I. Type 2 transglutaminase in coeliac disease: A key player in pathogenesis, diagnosis and therapy. Int. J. Mol. Sci. 2022, 23, 7513. [Google Scholar] [CrossRef]
- Basmanav, F.B.; Cesarato, N.; Kumar, S.; Borisov, O.; Kokordelis, P.; Ralser, D.J.; Wehner, M.; Axt, D.; Xiong, X.; Thiele, H.; et al. Assessment of the genetic spectrum of uncombable hair syndrome in a cohort of 107 individuals. JAMA Dermatol. 2022, 158, 1245–1253. [Google Scholar] [CrossRef] [PubMed]
- Piro, M.C.; Ventura, A.; Smirnov, A.; Saggini, A.; Lena, A.M.; Mauriello, A.; Bianchi, L.; Melino, G.; Candi, E. Transglutaminase 3 reduces the severity of psoriasis in imiquimod-treated mouse skin. Int. J. Mol. Sci. 2020, 21, 1566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ling, S.; Xu, B.; Luo, Y.; Fang, X.; Liu, X.; Wang, A.; Zhou, Y.; Zhang, S.; Zong, W.; Li, W.; et al. Transglutaminase 3 attenuates skin inflammation in psoriasis by inhibiting NF-κB activation through phosphorylated STAT3-TET3 signaling. J. Investig. Dermatol. 2022, 142, 2968–2977.e10. [Google Scholar] [CrossRef]
- Alshehri, F.S.M.; Whyte, C.S.; Mutch, N.J. Factor XIII-A: An indispensable "factor" in haemostasis and wound healing. Int. J. Mol. Sci. 2021, 22, 3055. [Google Scholar] [CrossRef] [PubMed]
- Steinert, P.M.; Chung, S.I.; Kim, S.Y. Inactive zymogen and highly active proteolytically processed membrane-bound forms of the transglutaminase 1 enzyme in human epidermal keratinocytes. Biochem. Biophys. Res. Commun. 1996, 221, 101–106. [Google Scholar] [CrossRef]
- Kim, H.C.; Nemes, Z.; Idler, W.W.; Hyde, C.C.; Steinert, P.M.; Ahvazi, B. Crystallization and preliminary X-ray analysis of human transglutaminase 3 from zymogen to active form. J. Struct. Biol. 2001, 135, 73–77. [Google Scholar] [CrossRef] [PubMed]
- Murata, T.; Honda, T.; Egawa, G.; Yamamoto, Y.; Ichijo, R.; Toyoshima, F.; Dainichi, T.; Kabashima, K. Transient elevation of cytoplasmic calcium ion concentration at a single cell level precedes morphological changes of epidermal keratinocytes during cornification. Sci. Rep. 2018, 8, 6610. [Google Scholar] [CrossRef] [Green Version]
- Agis, H.; Blei, J.; Watzek, G.; Gruber, R. Is zoledronate toxic to human periodontal fibroblasts? J. Dent. Res. 2010, 89, 40–45. [Google Scholar] [CrossRef]
- Hamburger, V.; Hamilton, H.L. A series of normal stages in the development of the chick embryo. J. Morphol. 1951, 88, 49–92. [Google Scholar] [CrossRef]
- Mallet, C.; Souci, L.; Ledevin, M.; Georgeault, S.; Larcher, T.; Denesvre, C. Establishment of a culture model for the prolonged maintenance of chicken feather follicles structure in vitro. PLoS ONE 2022, 17, e0271448. [Google Scholar] [CrossRef]
- Cau, L.; Pendaries, V.; Lhuillier, E.; Thompson, P.R.; Serre, G.; Takahara, H.; Méchin, M.-C.; Simon, M. Lowering relative hu-midity level increases epidermal protein deimination and drives human filaggrin breakdown. J. Dermatol. Sci. 2017, 86, 106–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sachslehner, A.P.; Surbek, M.; Golabi, B.; Geiselhofer, M.; Jäger, K.; Hess, C.; Kuchler, U.; Gruber, R.; Eckhart, L. Transglutaminase Activity Is Conserved in Stratified Epithelia and Skin Appendages of Mammals and Birds. Int. J. Mol. Sci. 2023, 24, 2193. https://doi.org/10.3390/ijms24032193
Sachslehner AP, Surbek M, Golabi B, Geiselhofer M, Jäger K, Hess C, Kuchler U, Gruber R, Eckhart L. Transglutaminase Activity Is Conserved in Stratified Epithelia and Skin Appendages of Mammals and Birds. International Journal of Molecular Sciences. 2023; 24(3):2193. https://doi.org/10.3390/ijms24032193
Chicago/Turabian StyleSachslehner, Attila Placido, Marta Surbek, Bahar Golabi, Miriam Geiselhofer, Karin Jäger, Claudia Hess, Ulrike Kuchler, Reinhard Gruber, and Leopold Eckhart. 2023. "Transglutaminase Activity Is Conserved in Stratified Epithelia and Skin Appendages of Mammals and Birds" International Journal of Molecular Sciences 24, no. 3: 2193. https://doi.org/10.3390/ijms24032193
APA StyleSachslehner, A. P., Surbek, M., Golabi, B., Geiselhofer, M., Jäger, K., Hess, C., Kuchler, U., Gruber, R., & Eckhart, L. (2023). Transglutaminase Activity Is Conserved in Stratified Epithelia and Skin Appendages of Mammals and Birds. International Journal of Molecular Sciences, 24(3), 2193. https://doi.org/10.3390/ijms24032193