Abstract
The emergence of chlorophyll-containing light-harvesting complexes (LHCs) was a crucial milestone in the evolution of photosynthetic eukaryotic organisms. Light-harvesting chlorophyll-binding proteins form complexes in proximity to the reaction centres of photosystems I and II and serve as an antenna, funnelling the harvested light energy towards the reaction centres, facilitating photochemical quenching, thereby optimizing photosynthesis. It is now generally accepted that the LHC proteins evolved from LHC-like proteins, a diverse family of proteins containing up to four transmembrane helices. Interestingly, LHC-like proteins do not participate in light harvesting to elevate photosynthesis activity under low light. Instead, they protect the photosystems by dissipating excess energy and taking part in non-photochemical quenching processes. Although there is evidence that LHC-like proteins are crucial factors of photoprotection, the roles of only a few of them, mainly the stress-related psbS and lhcSR, are well described. Here, we summarize the knowledge gained regarding the evolution and function of the various LHC-like proteins, with emphasis on those strongly related to photoprotection. We further suggest LHC-like proteins as candidates for improving photosynthesis in significant food crops and discuss future directions in their research.
1. In Addition to Functioning in Light Harvesting, the Light-Harvesting Proteins Are Important for Protection from Photoinhibition
Light-harvesting chlorophyll-binding complexes (LHCs) evolved in eukaryotic photosynthetic organisms to enhance the absorption cross-sections of photosystems I and II (PSI and PSII). LHCI and LHCII form megacomplexes with PSI and PSII (PSI-LHCI and PSII-LHCII) in thylakoid membranes and harvest light energy via the bound chlorophylls a and b. The harvested light energy is then directed towards the photosystem’s reaction centres (RCs) to drive charge separation and photosynthesis (Figure 1).
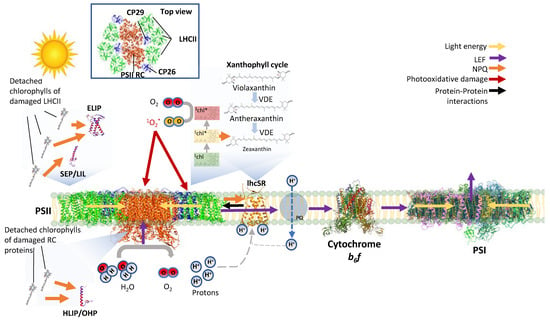
Figure 1.
Photosynthesis during HL and suggested photoprotective roles of LHC-like proteins. Light is harvested in the LHC and directed to PSII RC, where it excites a specialized chlorophyll-pair (P680) and initiates charge separation and linear electron flow (LEF). Electrons are transferred from PSII-RC via QA to QB that then join the plastoquinone (PQ) pool. From PQ, the electrons move to cytochrome b6f, PSI, and ferredoxin NADPH reductase (FNR) (not shown). FNR reduces NADP+ to NADPH, which is further utilized for carbon-fixation reaction via the Calvin–Benson–Bassham cycle. During HL, PSI and PSII are saturated with energy. The continuous photosynthesis facilitates a proton gradient between the lumen of the thylakoid and the stroma of the chloroplast due to the splitting of water and import of protons via the PQ pool and cytochrome b6f. This lumen acidification leads to protonation of protonable residues in psbS (mostly plants and mosses) and lhcSR (algae and mosses). In response, the pigment-less psbS interacts and activates NPQ in the LHCII subunit CP29. lhcSR binds lutein and chlorophyll and can act as a sinkhole for excess energy in the LHC. Thus, it can receive and scavenge excess energy from LHCII. When the light energy exceeds the amount that could be used for photochemistry, excited chlorophylls in the LHCs and RCs may dissipate the excess energy as heat via interactions with other chlorophylls or carotenoids. During HL, the xanthophyll cycle is activated: violaxanthin is de-epoxidated by VDE to zeaxanthin (via antheraxanthin), which in turn acts to relax excited chlorophylls and prevent ROS formation. Remaining excess energy excites chlorophyll to a triplet state where it reacts with O2 to form ROS. The ROS then interacts with and damages the photosynthetic reaction centres and LHC subunits, leading to photoinhibition. It is suggested that SEPs/Lils and ELIPs serve as a temporary reservoir for the chlorophylls of the damaged proteins during their turnover (see also Figure 2).
Owing to their significant enhancing effect on the ability of PSI and PSII to harvest sufficient light for efficient photosynthesis, LHCs have enabled eukaryotic photosynthetic organisms to evolve and thrive in various light-limited habitats. However, under intensive high-light (HL) intensities or when photosynthetic electron flow is inhibited, the efficient antenna property of LHCs is problematic and could increase photoinhibition. When the PSs are saturated with absorbed energy, the chlorophylls embedded in the LHC are excited with more energy than can actually be exploited by the photochemical reaction centres, resulting in the formation of chlorophyll triplets. These chlorophyll triplets react with O2 to form harmful reactive oxygen species (ROS) (Figure 1). In a similar fashion, ROS is produced in the RC of the PSII and PSI, possibly causing photoinhibition (PI), a condition in which photosynthesis is inhibited since the PS, primarily PSII, is damaged faster than the rate of being repaired (Figure 1 and Figure 2) [1,2,3]. During PI, photosynthesis declines up to a point where the organism may not survive. Photosynthetic organisms employ various mechanisms to minimize the effects of PI, starting from reduction in antenna (LHC) size to limit harvesting of excess light [4,5], through energy dissipation via non-photochemical quenching (NPQ) processes [6,7], and up to a rapid repair of the damaged photosynthetic apparatus [8,9,10] (Figure 1 and Figure 2). While LHCs function mainly as antennas that harvest light energy via chlorophylls and funnel it to the RCs (Figure 1 and Figure 5) [11,12,13,14], they also participate in the quenching of excess energy. An important and well described example of its role in energy quenching is the activation of the highly protective xanthophyll cycle during exposure to HL intensities. Violaxanthin, a carotenoid-bound to LHCII, is de-epoxidized by violaxanthin de-epoxidase (VDE) in a two-step manner first to antheraxanthin and then to zeaxanthin. Zeaxanthin then quenches the excess energy of nearby excited chlorophyll and prevents the production of harmful ROS, thereby protecting the PSII (Figure 1).
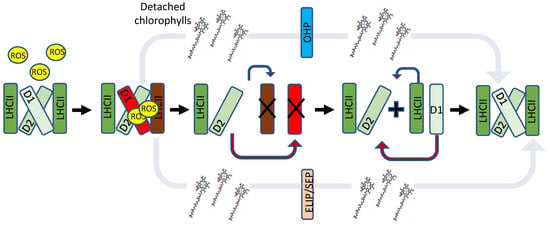
Figure 2.
Suggested roles of HLIP/OHP, ELIPs and SEP/Lil in the PSII repair cycle. During HL, generated ROS damages mainly the PSII reaction centre protein D1, followed by D2, and then the LHCII subunits. Chlorophylls from the damaged reaction centres and LHC are transferred to OHP1 or ELIP and SEP/Lil, respectively, while the damaged proteins are degraded. During this time, the energy of excited chlorophylls is quenched via carotenoids. Once newly synthesized subunits are incorporated and PSII reassembled, chlorophylls are transferred back to the reaction centre to harvest light for photosynthesis. It is important to note that ELIPs, SEPs and OHPs are most certainly involved in regulation of other mechanisms, including chlorophyll and carotenoid biosynthesis.
LHC proteins contain three transmembrane (TM) helices, of which two are homologous to HL-inducible protein (HLIP) (Figure 3a,b) and harbour a conserved amino acid sequence known as the LHC motif, as well conserved carotenoid locations and light-harvesting chlorophyll binding sites [15]. Intriguingly, it was discovered that LHC proteins evolved from proteins that do not participate in light harvesting (Figure 4), but instead serve to protect the PS RCs from excess light (Figure 1, Figure 2 and Figure 5) [16,17]. These LHC-like proteins share many structural similarities with the LHC proteins (Figure 3a,b), contain LHC motifs in their TM helices (Table 1), and are mostly able to bind pigments.
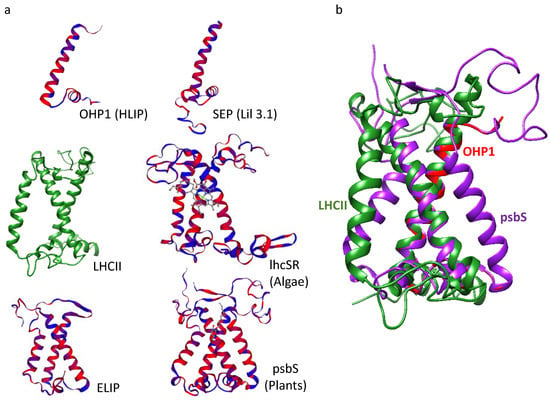
Figure 3.
LHCII and LHC-like proteins share vast structural similarities. (a) The known structure of LHCII (taken from pdb id: 6KAF) is shown in green. The structure models of LHC-like proteins as predicted by Swiss model. (b) OHP1 and psbS predicted models superimposed on the known LHCII structure. Note the overlapping of the two conserved TM helices in OHP, LHCI and psbS. All models (including OHP and SEP) were predicted with Arabidopsis thaliana sequences of the relevant proteins except lhcSR. The lhcSR model was predicted with C. reinhardtii sequence. The predicted structures are coloured by hydrophobicity: red indicates hydrophobic regions and blue indicates hydrophilic regions.
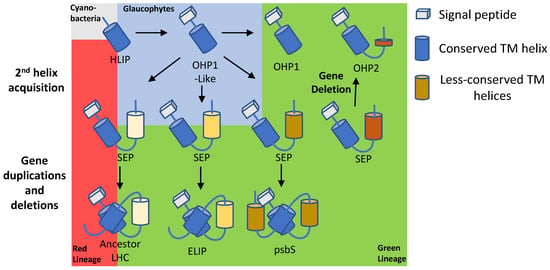
Figure 4.
Suggested evolution of LHC and LHC-like proteins. HLIP was first introduced to eukaryotes via primary endosymbiosis and acquired a signal peptide when its gene transferred to the nuclear genome. OHP1-like protein gave rise to a pool of different SEPs. OHP2 is likely a consequence of a gene-deletion event of SEP while a small portion remains and acts as a membrane anchor. Different SEPs evolved into LHCs, ELIPs and psbS.

Table 1.
Conserved LHC motif in selected photoprotective LHC-like proteins. The symbols * in the consensus sequence indicate less conserved residues. Adapted and modified from Rochaix and Bassi, 2019 [18].
2. LHC-like Proteins Do Not Serve as Antenna but Function in Protection against Photoinhibition
The first known ancestor of LHC and LHC-like proteins was discovered in the HL-stressed cyanobacteria Synechococcus and was named HL-inducible protein (HLIP) [19]. It is now thought that HLIP was involved in a series of gene fusions, duplications, and deletions, which gave rise to all other LHC-like and LHC proteins, with the latter being the most recently formed group [17,20] (Figure 4). The LHC-like proteins are classified into four groups based on their structural characteristics and the number of TM helices they carry (Table 2). Group 1 includes proteins with one TM helix. They include HLIPs in cyanobacteria and one-helix proteins (OHP) or small chlorophyll a/b binding (CAB) proteins in eukaryotes. Group 2 includes proteins with two TM helices, known as stress-enhanced proteins (SEP) or LHC-like proteins (Lil). Group 3 includes proteins with three TM helices, which contain both early light-induced proteins (ELIP) and stress-related LHCs (lhcSR). Group 4 contains a protein with four TM helices named PSII subunit S (psbS) [20]. LHC-like proteins are generally similar to LHCs in their amino acid sequence and also share structural similarities (Figure 3) [16,20,21]. However, as mentioned above, they do not participate in light harvesting to drive photosynthesis. On the contrary, there is evidence that at least some of these proteins have critical roles in the quenching of excess energy in the NPQ reactions (Figure 1, Figure 2, and Figure 5 and Table 2). Among these, the most studied are psbS and the stress-related LHCs (lhcSR), which are present in land plants and algae, respectively, while mosses carry both of them [6,22,23] (Figure 5). LHC-like proteins are generally not detectable in non-stressed environmental conditions, and are induced by various stress factors, including cold temperatures, limited nutrients, desiccation, and intense HL [24,25,26]. Consequently, it was suggested that LHC-like proteins take part in adaptation to growth-stress conditions. The LHC-like proteins, which are induced by HL, with emphasis on those reported to function in photoprotective mechanisms, are the focus of this review.
Table 2.
Summary of LHC-like proteins’ suggested pigments and functions.
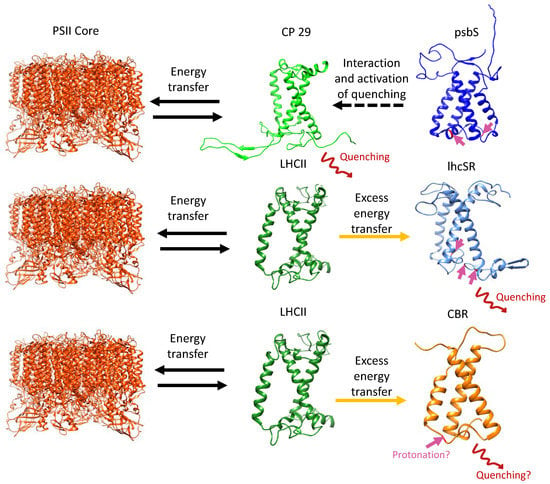
Figure 5.
Proposed quenching mechanisms for psbS, lhcSR, and C. ohadii CBR. HL conditions promote thylakoid lumen acidification. psbS (mainly in plants) is protonated at conserved Glu residues (indicated by pink arrows) and undergoes conformational shift before interacting with and activating energy quenching in the LHCII subunit CP29 (top). lhcSR is protonated at conserved Glu and Asp residues and also undergoes conformational shift and interacts with LHCII subunits. Excess energy can be transferred to lhcSR, where it is quenched via carotenoids (middle). The mechanism of CBR is unknown, yet evidence points to a conserved Glu residue with the same location of one of the proton-sensing Glutamates of psbS, suggesting a possibility in which CBR acts as a substitute for psbS and lhcSR in the highly light-tolerant C. ohadii (Levin et al., 2022) [27]. Figure adapted and modified from Pinnola., 2019 [23] LHCII and PSII core structures were taken from pdb id: 6KAF.
While the photoprotective roles of psbS and lhcSR are well defined (Figure 5) [23], little is known about the photoprotective mechanisms of other LHC-like proteins. The known and suggested photoprotective roles of LHC-like proteins include the sensing of thylakoid lumen acidification, activation of the photoprotective xanthophyll cycle, chlorophyll and carotenoid synthesis regulation, repair of PSI and PSII, and more [28,29,30,31]. Together, LHC-like proteins act as the guardians of photosynthesis in a variety of stress conditions. As such, it is important to explore the molecular mechanisms underlying their photoprotection capabilities. Doing so will enhance our understanding of PI resistance, and may identify candidates for improving photosynthesis in agriculture crops. For example, transgenic expression of Arabidopsis psbS alongside the key enzymes of the xanthophyll cycle VDE and zeaxanthin epoxidase (ZEP) in tobacco lead to faster NPQ relaxation kinetics during a shift from HL to LL conditions, which, in turn, accelerates CO2 assimilation recovery in the shade. Importantly, both field and greenhouse experiments resulted in a 15% increase in crop productivity, as measured by dry mass, and a 33% increase in seed yield [32,33,34]. It is of note that PsbS could not be detected in one of the most light-tolerant algae on the planet, Chlorella ohadii, and lhcSR is entirely absent from its genome [5,35], raising questions as to how this alga so efficiently ensures PI protection and why it eliminated these PI protection players in its evolution.
Herein, we review the suggested general characteristics and photoprotective roles of LHC-like proteins. We explore the potential of improving photosynthesis via the genetic engineering of photoprotective LHC-like proteins in agriculturally important crops, discuss the importance of a gaining a better understanding of the photoprotective functions of LHC-like proteins, and suggest future research directions.
3. lhcSR and psbS Are Stress-Induced Proteins That Protect Photosynthesis against Photoinhibition
Similar to LHC proteins, lhcSR contains three TM helices (Figure 3). It is found mainly in various algae lineages and mosses and binds chlorophylls and carotenoids [36,37]. In the model green alga Chlamydomonas reinhardtii, three genes encode lhcSR: lhcSR 1, and lhcSR’s 3.1 and 3.2, which the later share the same amino acid sequence. Excessive light conditions trigger lhcSR expression and their accumulation within several hours [38]. Protonatable conserved Glu and Asp residues located at the luminal side of lhcSR sense thylakoid lumen acidification [39], which occurs as a result of proton intake during the elevated activity of the photosynthetic electron flow under HL intensities (Figure 1 and Figure 5). This acidification activates qe, the pH-dependent component of NPQ [40]. lhcSR binds chlorophyll a, lutein and violaxanthin, which under HL is converted to zeaxanthin (via antheraxanthin) through consecutive de-epoxidation events, a process known as “the xanthophyll cycle” (Figure 1). While lhcSR associates with the LHCs, zeaxanthin quenches the excited chlorophylls and thereby prevents ROS formation in both PSI and PSII [41] (Figure 1 and Figure 5).
In land plants, psbS is the pH sensor and not lhcSR. However, while lhcSR has three TM helices, psbS contains four (Figure 3) and while it does carry LHC motifs, structural analysis of Spinacia oleracea psbS revealed a structure that is too tightly packed to bind pigments such as chlorophylls [42]. Therefore, psbS may be responsible for proton sensing and the activation of qe (the quenching of energy due to lowering the lumen pH) in higher plants, but it cannot be the quencher itself. Instead, it likely undergoes a conformational shift that facilitates its interaction with and activation of other LHC proteins for energy quenching, with recent studies indicating the minor antenna CP29 [43] (Figure 5). Interestingly, mosses contain both lhcSR and psbS, where both contribute to NPQ [44] and although initially psbS could not be detected in the model green algae C. reinhardtii under a variety of growth conditions [45], it was recently shown to be transiently induced under HL conditions. However, it was reported to accumulate to low levels, and to play a minor role in NPQ compared to lhcSR [46,47]. Surprisingly, psbS appears to be continuously expressed upon exposure to low light (LL) levels during a diurnal cycle [48,49]. Considering this evidence, psbS in C. reinhardtii was recently suggested to participate in the initial activation of lhcSR and NPQ, as well as in the reorganization of thylakoid membranes and LHC proteins [50]. Furthermore, the accumulation of lhcSR 1 and psbS in the thylakoid membrane of C. reinhardtii is induced by UV-B light using the nucleo-cytosolic UVR-8 photoreceptor. Accumulation of lhcSR 1 and psbS in UV-B light leads to a significantly better photoprotection under subsequent HL stress, which induces the accumulation of lhcSR 3 [51]. While most studies have focused on the protection of PSII via energy dissipation, under HL conditions the UV-B induced lhcSR 1 mediates energy transfer from LHCII to PSI instead of PSII, thus protecting it from excess energy [52].
4. One-Helix LHC-like Proteins (HLIPs/OHPs/Small-CABs) Protect PSII during Biogenesis and the Photoinhibition Repair Cycle
HLIPs were discovered in HL-stressed cyanobacteria [19] and are believed to be the earliest ancestor of all other LHC and LHC-like proteins [17,20]. HLIP homologs were later also found in eukaryotes [53] and were named OHPs or small chlorophyll a/b binding proteins (small-CABs). They possess a single trans-membrane helix, which harbours an LHC motif (Figure 3 and Table 1) and pigment-binding sites [54]. In both cyanobacteria and eukaryotic photosynthetic organisms, HLIPs/OHPs were shown to play central roles in the biogenesis and repair of PSII. In the cyanobacteria Synechocystis and Synechocococcus, there are four HLIP genes (HliA-D or ScpB-E) [55] which encode proteins that bind PSII and stabilize the trimeric structure of PSI, imperative for its activity under HL conditions [56,57,58]. HLIPs have been shown to retard PSII-associated chlorophyll degradation and have been suggested to temporarily bind the pigments of PSII during its repair, when its subunits are being substituted [56,59,60] (Figure 1 and Figure 2). In line with these reports, recent findings discovered HLIPs as part of chlorophyll-binding protein complexes that are important in the early assembly steps of PSII and chlorophyll biosynthesis [61,62]. A model of HLIP predicted a homo-dimeric structure and it was demonstrated that energy is dissipated via direct energy transfer between chlorophyll a and β-carotene [54]. The proteins may function by dissipating excess light energy that is absorbed by chlorophylls during the PSII repair cycle, when it cannot be utilized for photochemistry (Figure 1 and Figure 2).
In eukaryotic organisms, OHP1 and OHP2 replace HLIPs and are similarly induced by HL [53,63]. Recent studies suggest that they are crucial for PSII assembly and chlorophyll biosynthesis [30,64,65,66,67]. Mutations of either OHP1 or OHP2 in Arabidopsis resulted in less pigmentation, a disrupted thylakoid architecture and severe growth deficits, [66] while a deletion of OHP1 affected the function of core PSII proteins and resulted in lower accumulation of PSI protein [30]. Further studies indicated that OHP1 and OHP2 form a temporary protein complex with high chlorophyll florescence 244 (HCF244) and core proteins of PSII during PSII biogenesis [64,67] and that OHP function is fully dependent on its pigment-binding capacity [67]. These findings indicate that similarly to HLIPs, OHPs may temporarily associate with PSs and bind pigments while damaged core proteins are being replaced (Figure 1 and Figure 2).
5. Two-Helix LHC-like Proteins (SEPs/Lils) Function in Chlorophyll Biosynthesis and Protection against Photoinhibition
SEPs/Lils likely evolved when an early eukaryotic OHP-like protein acquired a second TM helix, which did not contain the LHC motif (Figure 3 and Figure 4). Later, different SEPs gave rise to LHCs, ELIPs, and psbS through gene duplications and, in the case of LHC and ELIP, an eventual loss of the fourth helix [17,20] (Figure 4). Four SEPs were identified in Arabidopsis: the poorly studied HL-induced Lil4 and Lil5 (formerly known as SEP1 and SEP2, respectively) and Lil3.1 and Lil3.2, which are present at constant levels regardless of the light regime [15,68]. Although its expression is independent of light, Lil3 forms a homo-multimeric chlorophyll-binding protein complex in dark-grown barley seedlings that are de-etiolated (transferred to light), suggesting light-dependent regulation of the complex formation [69]. Lil3 has been implicated in the stabilization of geranylgeranyl reductase (GGR) and has been found associated with protochlorophyllide oxidoreductase (POR), key enzymes in the production of crucial metabolites used for chlorophyll a biosynthesis [70,71]. Its participation in chlorophyll biosynthesis was strengthened by additional studies in barley and Arabidopsis [72,73]. Finally, Lil3 binds pigments and is capable of NPQ via direct energy transfer from chlorophyll a to a xanthophyll (lutein or zeaxanthin), suggesting that similar to HLIPs, it can also transfer pigments to their final destination while quenching damaging light energy [74]. In contrast to HLIPs/OHPs, Lil3 associates with LHCII subcomplexes rather that the PS core, which may be an indication of its distinct targets for photoprotection during PI [72] (Figure 2).
6. Three-Helix LHC-like Proteins (ELIPs) Play Important Roles in Protection against Photoinhibition
Like SEPs, ELIPs co-localize with the LHC and, similar to lhcSR and LHC, contain three TM helices and accumulate during HL stress [20,75,76,77]. ELIPs bind chlorophyll a and numerous molecules of lutein and have therefore been suggested to scavenge excited chlorophylls that are unbound to LHC during LHC protein turnover and thereby prevent ROS formation [78,79] (Figure 1 and Figure 2). Two ELIPs were identified in Arabidopsis: ELIP1 and ELIP2 [68]. An Arabidopsis mutant (chaos) that cannot rapidly accumulate ELIPs, suffered from bleaching and photooxidative damage during exposure to HL, due to ROS formation by excited chlorophylls [80]. The constitutive expression of ELIP genes prior to HL stress was sufficient to prevent these affects [80]. Moreover, the constitutive expression of ELIP2 resulted in a decrease in the levels of chlorophyll, and of glutamyl tRNA reductase and magnesium chelatase, which are key enzymes in chlorophyll biosynthesis [31]. Together, these observations suggest that ELIPs bind the chlorophyll of damaged LHCs while suppressing chlorophyll synthesis in order to avoid the accumulation of excited free chlorophylls during HL stress. This scenario was later challenged, as it was shown that a double elip1/elip2 mutant of Arabidopsis was just as prone to PI and photooxidative stress as WT and did not accumulate free pigments [29]. It was noted that zeaxanthin was decreased in the elip1/elip2 mutant, perhaps indicating that ELIP regulates the protective xanthophyll cycle. In an additional study, the enhanced protection of PSII from PI was noted in an Arabidopsis mutant lacking ELIP2 but transgenically expressing the ELIP1/2 of the desiccation-tolerant moss Syntrichia caninervis [81].
Ten ELIP homologs were identified in Chlamydomonas, some of which are induced by HL stress [82,83]. ELIP3 is upregulated by a combination of HL and cold stress, resulting in improved survival of C. reinhardtii under photooxidative stress and higher PSII activity [26]. In accordance, an ELIP3 knockdown mutant showed significantly less PSII activity and did not survive under HL and cold stress conditions [26]. Moreover, the addition of a ROS quencher reduced ELIP3 expression, suggesting that ROS trigger ELIP3 expression [26]. In the green alga Dunaliella salina, an ELIP homolog is co-regulated with carotenoid biosynthesis and was therefore named carotenoid biosynthesis-related protein (CBR) [84]. CBR associates with LHCII, binds zeaxanthin [85,86], and was suggested to be involved in the organization and/or stabilization of PI-protective protein complexes with high lutein and zeaxanthin content [87]. Interestingly, CBR is the most upregulated protein in HL-adapted Chlorella ohadii cells, one of the most light-tolerant photosynthetic organisms [5,27]. Analysis of CBR accumulation in C. ohadii cells grown under HL found co-localization of CBR and LHCII, and ranked it to be the most abundant LHC-like protein with a light-dependent expression pattern [27]. It is important to note that C. ohadii does not contain genes encoding lhcSR [35] and psbS was found to be expressed below the proteomic detection levels under HL conditions [5,27]. These results implied that CBR plays a key role in the photoprotection of photosynthesis in an extremely light-tolerant organism, which lacks lhcSR and expresses limited or no quantities of psbS. Intriguingly, C. ohadii CBR has been found to be homologous to the abovementioned ELIP3 of Chlamydomonas, which was shown to be induced by ROS formation and crucial for its survival under HL and cold stress [26,27]. Additionally, amino acid sequence analysis and structural modelling of CBR implied that a conserved glutamate is located in the same position as one of the proton-sensing glutamates of psbS, suggesting, in the absence of psbs and lhcSR, a possible role in sensing low pH in the lumen during HL (Figure 5).
7. Soluble Pigment Binding Proteins also Contribute to Photoprotection
As described, LHC and LHC-like proteins share distinct structural similarities, are located in the thylakoid membrane, and contribute to photoprotection. These proteins, with the exception of psbS, bind both carotenoids and chlorophylls. In addition, soluble pigment-binding proteins also offer alternate routes for energy dissipation via NPQ and thus promote photoprotection. The structures of these proteins differ significantly from the membranal LHC and LHC-like proteins and they do not share the LHC and LHC-like ancestor protein. In cyanobacteria, one such protein is the orange carotenoid protein (OCP) [88]. OCP forms a homodimer complex where each monomer binds a single carotenoid [89]. Upon activation by sensing blue-green light, OCP undergoes a conformational change and interact with the cyanobacterial photosynthetic antenna (phycobilisome). Following this, via its bound carotenoid, it presents a route for the thermal dissipation of excess energy and acts as a singlet oxygen quencher [90,91,92,93].
In plants, an example is the water-soluble chlorophyll-binding protein (WSCP) [94]. WSCPs form a homo tetramer and, contrary to the cyanobacterial OPC, bind one chlorophyll per each monomer, but no carotenoids [95]. The exact function of WSCPs remain a mystery; however, evidence suggests a role in photoprotection, despite the lack of carotenoids. Chlorophyll bound to WSCP is much less sensitive to intense light compared to unbound chlorophylls due to a formation of a physical barrier by the phytyl moieties, shielding the oxidizable sites of the chlorophyll, and thus preventing damage by ROS to the chlorophylls themselves [95,96,97]. Moreover, the overexpression of WSCP in transgenic plants leads to a reduction in ROS generation, highlighting its role as a protective protein under stress [98].
8. LHC-like Proteins Function as Enhancers of Photosynthesis and Survivability during Abiotic Stress and Their Overexpression May Increase Crop Yield
Considering the growth of the Earth’s population and the anticipated food shortage, new approaches to increase food production should be explored and developed. Improving photosynthesis as a means of enhancing crop growth and yields has been a long-standing aim [33,34,99,100,101]. Other techniques implemented to achieve this important goal include the selection of light-tolerant strains and improving the efficiency of CO2 assimilation and light-to-biomass conversion, as well as the manipulation of metabolic pathways by genetic engineering. Alongside their crucial roles in the protection of photosynthesis from photoinhibition, studies have suggested the potential of LHC-like proteins as photosynthesis enhancers. The overexpression of psbS in field-grown tobacco led to a decrease in stomatal opening in response to light, which resulted in a significant 25% reduction in water loss per assimilated CO2 molecule [102]. As described above, the transgenic expression of Arabidopsis psbS alongside VDE and ZEP in tobacco accelerated NPQ relaxation kinetics during a shift from HL to LL conditions, which, in turn, shortened time to CO2 assimilation recovery in the shade. Importantly, both field and greenhouse experiments resulted in a 15% rise in crop productivity, as measured by dry mass [32]. Moreover, using the same strategy in the important crop soybean grown in fields, leads to an increase of up to 33% in seed yield [34]. In rice, another highly important crop, psbS overexpression resulted in a higher grain yield under fluctuating light conditions [103]. These studies provide proof of the ability to increase biomass and yield in important crops via the overexpression of the LHC-like protein psbS, exclusively or alongside other proteins. While psbS is the most studied LHC-like protein, other members of the family were also shown to increase biomass and to provide protection under abiotic stress. In C. reinhardtii mutants lacking all lhcSRs, expression of lhcSR under the control of a heat shock protein promoter allowed for the fine-tuning of NPQ, resulting in higher photosynthetic efficiency and biomass accumulation [104]. Transgenic tobacco overexpressing ELIP showed higher survivability after freezing, a higher net photosynthesis rate after chilling, a higher plant fresh weight after osmotic stress, and higher maximal photochemical efficiency after HL treatment [105]. As mentioned earlier, the overexpression of ELIP3 in Chlamydomonas improved the survival of the cells under HL and cold stress. In contrast, a knockdown mutant showed low photosynthetic efficiency and rapidly died under these conditions [26]. Similarly, the overexpression of Rhododendron ELIP in Arabidopsis resulted in a much stronger tolerance to freezing and higher PSII function after 3 days of recovery from freezing [106]. A recent study in the red alga Neopyropia yezoensis overexpressing OHPs failed to detect any obvious enhancement of photoprotection [107]. In contrast, Arabidopsis OHP knockout mutants exhibited higher susceptibility to HL [108], were unable to grow on soil [30], and demonstrated compromised growth [66]. Taken together, these studies highlight the potential of using psbS and other LHC-like proteins to protect plants and improve photosynthesis under various abiotic stresses.
9. Concluding Remarks and Future Perspectives
The LHC-like proteins form a large group of mostly HL-induced TM proteins that are capable of binding pigments and performing NPQ via energy transfer from chlorophylls to carotenoids. An exception to this rule is psbS, which is unable to bind pigments due to its tightly packed structure, containing four TM helices [42] (Figure 3a). Instead, psbS is protonated upon acidification of the thylakoid lumen and undergoes a conformational shift before interacting with PSII and activating NPQ in the LHCII minor antenna CP29 [43] (Figure 5). Similarly, under acidic conditions, lhcSR is protonated and associates with LHCII proteins [39], but unlike psbS, it binds carotenoids that offer a route for the thermal energy dissipation of excess energy in the LHCs of both PSs [41] (Figure 1 and Figure 5). Similarly, ELIPs and SEPs associate with LHCII subunits and bind pigments. Their suggested role involves the temporary binding of chlorophylls that were detached from damaged LHC subunits during their turnover under HL stress, while simultaneously regulating chlorophyll synthesis [72,74,78] (Figure 1 and Figure 2). In C. ohadii, one of the most light-tolerant photosynthetic organisms on the planet [35], an ELIP homolog named CBR accumulates massively under HL and co-localizes with LHCII [5,27]. The absence of lhcSR from the genome of C. ohadii and the failed detection of psbS in both LL- and HL-grown cells [5,27] raise questions about the significance and function of CBR in this particular organism. It was suggested that CBR is activated in a similar manner to psbS and lhcSR, as they all contain a conserved protonable Glu on their luminal side [27] (Figure 5). Unlike the abovementioned proteins, HLIPs in cyanobacteria and OHPs in eukaryotes seem to associate with the PS core proteins, rather than with the LHC, where they are also suggested to act as a temporary reservoir of detached pigments from damaged core proteins during their turnover in HL stress [56,60,64,67] (Figure 1 and Figure 2). The functions of the LHC-like proteins discussed in this review are summarized in Table 2.
Although the exact molecular mechanisms through which LHC-like proteins act to protect the PSs from PI are yet to be fully uncovered, it is clear that they play key roles in photoprotection, which can be exploited to increase the yield and survivability of agriculturally important crops. Recent experiments have shown this potential by overexpressing psbS, resulting in increased crop productivity and grain yield while reducing water loss due to stomatal opening in plants exposed to fluctuating light [32,34,102,103]. These promising results highlight the biotechnological potential and importance of understanding and utilizing LHC-like proteins to improve photoprotection capacities. Notably, psbS is by far the most studied LHC-like protein. However, as described above, other LHC-like proteins have shown to increase biomass and protect photosynthesis and should definitely be further tested for their ability to also increase crop productivity. In the future, psbS and the other LHC-like proteins can be introduced to important crops via genetic engineering, both to increase their yield and expand their environmental niche by making them more resilient to light and other stresses.
Author Contributions
G.L. and G.S. wrote this paper. Both authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Israel Science Foundation grant, 2199/22.
Institutional Review Board Statement
Not applicable.
Conflicts of Interest
The authors declare no competing financial interests.
References
- Schuster, G.; Timberg, R.; Ohad, I. Turnover of Thylakoid Photosystem II Proteins during Photoinhibition of Chlamydomonas reinhardtii. Eur. J. Biochem. 1988, 177, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Nishiyama, Y.; Allakhverdiev, S.I.; Murata, N. A New Paradigm for the Action of Reactive Oxygen Species in the Photoinhibition of Photosystem II. Biochim. Biophys. Acta Bioenerg. 2006, 1757, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Pospíšil, P. Production of Reactive Oxygen Species by Photosystem II. Biochim. Biophys. Acta Bioenerg. 2009, 1787, 1151–1160. [Google Scholar] [CrossRef]
- Erickson, E.; Wakao, S.; Niyogi, K.K. Light Stress and Photoprotection in Chlamydomonas reinhardtii. Plant J. 2015, 82, 449–465. [Google Scholar] [CrossRef] [PubMed]
- Levin, G.; Kulikovsky, S.; Liveanu, V.; Eichenbaum, B.; Meir, A.; Isaacson, T.; Tadmor, Y.; Adir, N.; Schuster, G. The Desert Green Algae Chlorella Ohadii Thrives at Excessively High Light Intensities by Exceptionally Enhancing the Mechanisms That Protect Photosynthesis from Photoinhibition. Plant J. 2021, 106, 1260–1277. [Google Scholar] [CrossRef] [PubMed]
- Niyogi, K.K.; Truong, T.B. Evolution of Flexible Non-Photochemical Quenching Mechanisms That Regulate Light Harvesting in Oxygenic Photosynthesis. Curr. Opin. Plant Biol. 2013, 16, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Croce, R.; van Amerongen, H. Natural Strategies for Photosynthetic Light Harvesting. Nat. Chem. Biol. 2014, 10, 492–501. [Google Scholar] [CrossRef]
- Melis, A. Photosystem-II Damage and Repair Cycle in Chloroplasts: What Modulates the Rate of Photodamage in Vivo? Trends Plant Sci. 1999, 4, 130–135. [Google Scholar] [CrossRef]
- Spetea, C.; Nishiyama, Y.; Gollan, P.J.; Liu, J.; Cn, L.; Hua, W.; Lu, Y.; Last, R.L. A New Light on Photosystem II Maintenance in Oxygenic Photosynthesis. Front. Plant Sci. 2019, 10, 975. [Google Scholar] [CrossRef]
- Nixon, P.J.; Michoux, F.; Yu, J.; Boehm, M.; Komenda, J. Recent Advances in Understanding the Assembly and Repair of Photosystem II. Ann. Bot. 2010, 106, 1–16. [Google Scholar] [CrossRef]
- Drop, B.; Webber-Birungi, M.; Yadav, S.K.N.; Filipowicz-Szymanska, A.; Fusetti, F.; Boekema, E.J.; Croce, R. Light-Harvesting Complex II (LHCII) and Its Supramolecular Organization in Chlamydomonas reinhardtii. Biochim. Biophys. Acta Bioenerg. 2014, 1837, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Caspy, I.; Neumann, E.; Fadeeva, M.; Liveanu, V.; Savitsky, A.; Frank, A.; Levi Kalisman, Y.; Shkolnisky, Y.; Murik, O.; Treves, H.; et al. Cryo-EM Photosystem I Structure Reveals Adaptation Mechanisms to Extreme High Light in Chlorella ohadii. Nat. Plants 2021, 7, 1314–1322. [Google Scholar] [CrossRef] [PubMed]
- Shen, L.; Huang, Z.; Chang, S.; Wang, W.; Wang, J.; Kuang, T.; Han, G.; Shen, J.R.; Zhang, X. Structure of a C2S2M2N2-Type PSII–LHCII Supercomplex from the Green Alga Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2019, 116, 21246–21255. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Yan, H.; Wang, K.; Kuang, T.; Zhang, J.; Gui, L.; An, X.; Chang, W. Crystal Structure of Spinach Major Light-Harvesting Complex at 2.72 A˚ Resolution. Nature 2004, 428, 287–292. [Google Scholar] [CrossRef] [PubMed]
- Jansson, S. A Guide to the Lhc Genes and Their Relatives in Arabidopsis. Trends Plant Sci. 1999, 4, 236–240. [Google Scholar] [CrossRef]
- Montané, M.-H.; Kloppstech, K. The Family of Light-Harvesting-Related Proteins (LHCs, ELIPs, HLIPs): Was the Harvesting of Light Their Primary Function? Gene 2000, 258, 1–8. [Google Scholar] [CrossRef]
- Engelken, J.; Brinkmann, H.; Adamska, I. Taxonomic Distribution and Origins of the Extended LHC (Light-Harvesting Complex) Antenna Protein Superfamily. BMC Evol. Biol. 2010, 10, 233. [Google Scholar] [CrossRef]
- Rochaix, J.D.; Bassi, R. LHC-like Proteins Involved in Stress Responses and Biogenesis/Repair of the Photosynthetic Apparatus. Biochem. J. 2019, 476, 581–593. [Google Scholar] [CrossRef]
- Dolganov, N.A.M.; Bhayat, D.; Grossman, A.R. Cyanobacterial Protein with Similarity to the Chlorophyll a/b Binding Proteins of Higher Plants: Evolution and Regulation. Proc. Natl. Acad. Sci. USA 1995, 92, 636–640. [Google Scholar] [CrossRef]
- Heddad, M.; Adamska, I. The Evolution of Light Stress Proteins in Photosynthetic Organisms. Comp. Funct. Genom. 2002, 3, 504–510. [Google Scholar] [CrossRef]
- Bassi, R.; Dall’osto, L. Dissipation of Light Energy Absorbed in Excess: The Molecular Mechanisms. Annu. Rev. Plant Biol. 2021, 72, 47–76. [Google Scholar] [CrossRef] [PubMed]
- Li, X.P.; Gilmore, A.M.; Caffarri, S.; Bassi, R.; Golan, T.; Kramer, D.; Niyogi, K.K. Regulation of Photosynthetic Light Harvesting Involves Intrathylakoid Lumen PH Sensing by the PsbS Protein. J. Biol. Chem. 2004, 279, 22866–22874. [Google Scholar] [CrossRef] [PubMed]
- Pinnola, A.; Griffiths, H. The Rise and Fall of Light-Harvesting Complex Stress-Related Proteins as Photoprotection Agents during Evolution. J. Exp. Bot. 2019, 70, 5527–5535. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Q.; Chen, X.; Wood, A.J. Two Early Light-Inducible Protein (ELIP) CDNAs from the Resurrection Plant Tortula Ruralis Are Differentially Expressed in Response to Desiccation, Rehydration, Salinity, and High Light. J. Exp. Bot. 2002, 53, 1197–1205. [Google Scholar] [CrossRef]
- Guo, J.; Wilken, S.; Jimenez, V.; Choi, C.J.; Ansong, C.; Dannebaum, R.; Sudek, L.; Milner, D.S.; Bachy, C.; Nahas Reistetter, E.; et al. Specialized Proteomic Responses and an Ancient Photoprotection Mechanism Sustain Marine Green Algal Growth during Phosphate Limitation. Nat. Microbiol. 2018, 3, 781–790. [Google Scholar] [CrossRef]
- Lee, J.W.; Lee, S.H.; Han, J.W.; Kim, G.H. Early Light-Inducible Protein (ELIP) Can Enhance Resistance to Cold-Induced Photooxidative Stress in Chlamydomonas reinhardtii. Front. Physiol. 2020, 11, 1083. [Google Scholar] [CrossRef]
- Levin, G.; Yasmin, M.; Simanowitz, M.C.; Meir, A.; Tadmor, Y.; Hirschberg, J.; Adir, N.; Schuster, G. A Desert Green Alga That Thrives at Extreme High-Light Intensities Using a Unique Photoinhibition Protection Mechanism. bioRxiv 2022. [Google Scholar] [CrossRef]
- Zhao, L.; Cheng, D.; Huang, X.; Chen, M.; Dall’osto, L.; Xing, J.; Gao, L.; Li, L.; Wang, Y.; Bassi, R.; et al. A Light Harvesting Complex-Like Protein in Maintenance of Photosynthetic Components in Chlamydomonas. Plant Physiol. 2017, 174, 2419–2433. [Google Scholar] [CrossRef]
- Rossini, S.; Casazza, A.P.; Engelmann, E.C.M.; Havaux, M.; Jennings, R.C.; Soave, C. Suppression of Both ELIP1 and ELIP2 in Arabidopsis Does Not Affect Tolerance to Photoinhibition and Photooxidative Stress. Plant Physiol. 2006, 141, 1264–1273. [Google Scholar] [CrossRef]
- Myouga, F.; Takahashi, K.; Tanaka, R.; Nagata, N.; Kiss, A.Z.; Funk, C.; Nomura, Y.; Nakagami, H.; Jansson, S.; Shinozaki, K. Stable Accumulation of Photosystem II Requires ONE-HELIX PROTEIN1 (OHP1) of the Light Harvesting-like Family. Plant Physiol. 2018, 176, 2277–2291. [Google Scholar] [CrossRef]
- Tzvetkova-Chevolleau, T.; Franck, F.; Alawady, A.E.; Dall’Osto, L.; Carrière, F.; Bassi, R.; Grimm, B.; Nussaume, L.; Havaux, M. The Light Stress-Induced Protein ELIP2 Is a Regulator of Chlorophyll Synthesis in Arabidopsis thaliana. Plant J. 2007, 50, 795–809. [Google Scholar] [CrossRef] [PubMed]
- Kromdijk, J.; Głowacka, K.; Leonelli, L.; Gabilly, S.T.; Iwai, M.; Niyogi, K.K.; Long, S.P. Improving Photosynthesis and crop Productivity by Accelerating from Photoprotection. Science (1979) 2016, 354, 857–861. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.G.; Long, S.P.; Ort, D.R. Improving Photosynthetic Efficiency for Greater Yield. Annu. Rev. Plant Biol. 2010, 61, 235–261. [Google Scholar] [CrossRef] [PubMed]
- de Souza, A.P.; Burgess, S.J.; Doran, L.; Hansen, J.; Manukyan, L.; Maryn, N.; Gotarkar, D.; Leonelli, L.; Niyogi, K.K.; Long, S.P. Soybean Photosynthesis and Crop Yield Are Improved by Accelerating Recovery from Photoprotection. Science (1979) 2022, 377, 851–854. [Google Scholar] [CrossRef] [PubMed]
- Treves, H.; Raanan, H.; Kedem, I.; Murik, O.; Keren, N.; Zer, H.; Berkowicz, S.M.; Giordano, M.; Norici, A.; Shotland, Y.; et al. The Mechanisms Whereby the Green Alga Chlorella Ohadii, Isolated from Desert Soil Crust, Exhibits Unparalleled Photodamage Resistance. New Phytol. 2016, 210, 1229–1243. [Google Scholar] [CrossRef]
- Peers, G.; Truong, T.B.; Ostendorf, E.; Busch, A.; Elrad, D.; Grossman, A.R.; Hippler, M.; Niyogi, K.K. An Ancient Light-Harvesting Protein Is Critical for the Regulation of Algal Photosynthesis. Nature 2009, 462, 518–521. [Google Scholar] [CrossRef]
- Prandi, I.G.; Sláma, V.; Pecorilla, C.; Cupellini, L.; Mennucci, B. Structure of the Stress-Related LHCSR1 Complex Determined by an Integrated Computational Strategy. Commun. Biol. 2022, 5, 145. [Google Scholar] [CrossRef]
- Mou, S.; Zhang, X.; Ye, N.; Dong, M.; Liang, C.; Liang, Q.; Miao, J.; Xu, D.; Zheng, Z. Cloning and Expression Analysis of Two Different LhcSR Genes Involved in Stress Adaptation in an Antarctic Microalga, Chlamydomonas sp. ICE-L. Extremophiles 2012, 16, 193–203. [Google Scholar] [CrossRef]
- Ballottari, M.; Truong, T.B.; Re De, E.; Erickson, E.; Stella, G.R.; Fleming, G.R.; Bassi, R.; Niyogi, K.K. Identification of Ph-Sensing Sites in the Light Harvesting Complex Stress-Related 3 Protein Essential for Triggering Non-Photochemical Quenching in Chlamydomonas reinhardtii. J. Biol. Chem. 2016, 291, 7334–7346. [Google Scholar] [CrossRef] [PubMed]
- Liguori, N.; Roy, L.M.; Opacic, M.; Durand, G.; Croce, R. Regulation of Light Harvesting in the Green Alga Chlamydomonas reinhardtii: The C-Terminus of LHCSR Is the Knob of a Dimmer Switch. J. Am. Chem. Soc. 2013, 135, 18339–18342. [Google Scholar] [CrossRef]
- Girolomoni, L.; Cazzaniga, S.; Pinnola, A.; Perozeni, F.; Ballottari, M.; Bassi, R. LHCSR3 Is a Nonphotochemical Quencher of Both Photosystems in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2019, 116, 4212–4217. [Google Scholar] [CrossRef] [PubMed]
- Fan, M.; Li, M.; Liu, Z.; Cao, P.; Pan, X.; Zhang, H.; Zhao, X.; Zhang, J.; Chang, W. Crystal Structures of the PsbS Protein Essential for Photoprotection in Plants. Nat. Struct. Mol. Biol. 2015, 22, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Guardini, Z.; Bressan, M.; Caferri, R.; Bassi, R.; Dall’Osto, L. Identification of a Pigment Cluster Catalysing Fast Photoprotective Quenching Response in CP29. Nat. Plants 2020, 6, 303–313. [Google Scholar] [CrossRef]
- Alboresia, A.; Gerottob, C.; Giacomettib, G.M.; Bassia, R.; Morosinotto, T. Physcomitrella Patens Mutants Affected on Heat Dissipation Clarify the Evolution of Photoprotection Mechanisms upon Land Colonization. Proc. Natl. Acad. Sci USA 2010, 107, 11128–11133. [Google Scholar] [CrossRef]
- Bonente, G.; Passarini, F.; Cazzaniga, S.; Mancone, C.; Buia, M.C.; Tripodi, M.; Bassi, R.; Caffarri, S. The Occurrence of the PsbS Gene Product in Chlamydomonas reinhardtii and in Other Photosynthetic Organisms and Its Correlation with Energy Quenching. Photochem. Photobiol. 2008, 84, 1359–1370. [Google Scholar] [CrossRef] [PubMed]
- Correa-Galvis, V.; Redekop, P.; Guan, K.; Griess, A.; Truong, T.B.; Wakao, S.; Niyogi, K.K.; Jahns, P. Photosystem II Subunit PsbS Is Involved in the Induction of LHCSR Protein-Dependent Energy Dissipation in Chlamydomonas Reinhardtii. J. Biol. Chem. 2016, 291, 17478–17487. [Google Scholar] [CrossRef] [PubMed]
- Tibiletti, T.; Auroy, P.; Peltier, G.; Caffarri, S. Chlamydomonas reinhardtii PsbS Protein Is Functional and Accumulates Rapidly and Transiently under High Light. Plant Physiol. 2016, 171, 2717–2730. [Google Scholar] [CrossRef]
- Redekop, P.; Sanz-Luque, E.; Yuan, Y.; Villain, G.; Petroutsos, D.; Grossman, A.R. Transcriptional Regulation of Photoprotection in Dark-to-Light Transition-More than Just a Matter of Excess Light Energy. Sci. Adv. 2022, 8, 1832. [Google Scholar] [CrossRef]
- Nawrocki, W.J.; Liu, X.; Croce, R. Chlamydomonas reinhardtii Exhibits De Facto Constitutive NPQ Capacity in Physiologically Relevant Conditions. Plant Physiol. 2020, 182, 472–479. [Google Scholar] [CrossRef]
- Redekop, P.; Rothhausen, N.; Rothhausen, N.; Melzer, M.; Mosebach, L.; Dülger, E.; Bovdilova, A.; Caffarri, S.; Hippler, M.; Jahns, P. PsbS Contributes to Photoprotection in Chlamydomonas reinhardtii Independently of Energy Dissipation. Biochim. Biophys. Acta Bioenerg. 2020, 1861, 148183. [Google Scholar] [CrossRef]
- Allorent, G.; Lefebvre-Legendre, L.; Chappuis, R.; Kuntz, M.; Truong, T.B.; Niyogi, K.K.; Ulm, R.; Goldschmidt-Clermont, M. UV-B Photoreceptor-Mediated Protection of the Photosynthetic Machinery in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2016, 113, 14864–14869. [Google Scholar] [CrossRef] [PubMed]
- Kosuge, K.; Tokutsu, R.; Kim, E.; Akimoto, S.; Yokono, M.; Ueno, Y.; Minagawa, J. LHCSR1-Dependent Fluorescence Quenching Is Mediated by Excitation Energy Transfer from LHCII to Photosystem I in Chlamydomonas reinhardtii. Proc. Natl. Acad. Sci. USA 2018, 115, 3722–3727. [Google Scholar] [CrossRef] [PubMed]
- Andersson, U.; Heddad, M.; Adamska, I. Light Stress-Induced One-Helix Protein of the Chlorophyll a/b-Binding Family Associated with Photosystem I. Plant Physiol. 2003, 132, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Staleva, H.; Komenda, J.; Shukla, M.K.; Ka, R.; Sobotka, R. Mechanism of Photoprotection in the Cyanobacterial Ancestor of Plant Antenna Proteins. Nat. Chem. Biol. 2015, 11, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Funk, C.; Vermaas, W. A Cyanobacterial Gene Family Coding for Single-Helix Proteins Resembling Part of the Light-Harvesting Proteins from Higher Plants. Biochemistry 1999, 38, 9397–9404. [Google Scholar] [CrossRef]
- Yao, D.; Kieselbach, T.; Komenda, J.; Promnares, K.; Hernández Prieto, M.A.; Tichy, M.; Vermaas, W.; Funk, C. Localization of the Small CAB-like Proteins in Photosystem II. J. Biol. Chem. 2007, 282, 267–276. [Google Scholar] [CrossRef]
- Komenda, J.; Sobotka, R. Cyanobacterial High-Light-Inducible Proteins - Protectors of Chlorophyll-Protein Synthesis and Assembly. Biochim. Biophys. Acta Bioenerg. 2016, 1857, 288–295. [Google Scholar] [CrossRef]
- Wang, Q.; Jantaro, S.; Lu, B.; Majeed, W.; Bailey, M.; He, Q. The High Light-Inducible Polypeptides Stabilize Trimeric Photosystem I Complex under High Light Conditions in Synechocystis PCC 6803. Plant Physiol. 2008, 147, 1239–1250. [Google Scholar] [CrossRef]
- Promnares, K.; Komenda, J.; Bumba, L.; Nebesarova, J.; Vacha, F.; Tichy, M. Cyanobacterial Small Chlorophyll-Binding Protein ScpD (HliB) Is Located on the Periphery of Photosystem II in the Vicinity of PsbH and CP47 Subunits. J. Biol. Chem. 2006, 281, 32705–32713. [Google Scholar] [CrossRef]
- Vavilin, D.; Yao, D.; Vermaas, W. Small Cab-like Proteins Retard Degradation of Photosystem II-Associated Chlorophyll in Synechocystis Sp. PCC 6803: Kinetic Analysis of Pigment Labeling with 15N and 13C. J. Biol. Chem. 2007, 282, 37660–37668. [Google Scholar] [CrossRef]
- Knoppová, J.; Sobotka, R.; Tichý, M.; Yu, J.; Konik, P.; Halada, P.; Nixon, P.J.; Komenda, J. Discovery of a Chlorophyll Binding Protein Complex Involved in the Early Steps of Photosystem II Assembly in Synechocystis. Plant Cell 2014, 26, 1200–1212. [Google Scholar] [CrossRef] [PubMed]
- Chidgey, J.W.; Linhartová, M.; Komenda, J.; Jackson, P.J.; Dickman, M.J.; Canniffe, D.P.; Koník, P.; Pilný, J.; Hunter, C.N.; Sobotka, R. A Cyanobacterial Chlorophyll Synthase-HliD Complex Associates with the Ycf39 Protein and the YidC/Alb3 Insertase. Plant Cell 2014, 26, 1267–1279. [Google Scholar] [CrossRef] [PubMed]
- Jansson, S.; Andersson, J.; Kim, S.J.; Jackowski, G. An Arabidopsis Thaliana Protein Homologous to Cyanobacterial High-Light-Inducible Proteins. Plant Mol. Biol. 2000, 42, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Liu, B.; Zhang, J.; Kong, F.; Zhang, L.; Meng, H.; Li, W.; Rochaix, J.D.; Li, D.; Peng, L. OHP1, OHP2, and HCF244 Form a Transient Functional Complex with the Photosystem II Reaction Center. Plant Physiol. 2019, 179, 195–208. [Google Scholar] [CrossRef] [PubMed]
- Hey, D.; Grimm, B. ONE-HELIX PROTEIN2 (OHP2) Is Required for the Stability of OHP1 and Assembly Factor HCF244 and Is Functionally Linked to PSII Biogenesis. Plant Physiol. 2018, 177, 1453–1472. [Google Scholar] [CrossRef] [PubMed]
- Jahns, P.; Sakamoto, W.; Picorel, R.; Funck, D.; Beck, J.; Lohscheider, J.N.; Albert, S.; Andersson, U.; Mendgen, K.W.; Rojas-Stütz, M.C.; et al. Small One-Helix Proteins Are Essential for Photosynthesis in Arabidopsis. Front. Plant Sci. 2017, 8, 7. [Google Scholar] [CrossRef]
- Hey, D.; Grimm, B. One-Helix Protein 1 and 2 Form Heterodimers to Bind Chlorophyll in Photosystem II Biogenesis. Plant Physiol. 2020, 183, 179–193. [Google Scholar] [CrossRef]
- Heddad, M.; Adamska, I. Light Stress-Regulated Two-Helix Proteins in Arabidopsis Thaliana Related to the Chlorophyll a/b-Binding Gene Family. Proc. Natl. Acad. Sci. USA 2000, 97, 3741–3746. [Google Scholar] [CrossRef]
- Reisinger, V.; Plöscher, M.; Eichacker, L.A. Lil3 Assembles as Chlorophyll-Binding Protein Complex during Deetiolation. FEBS Lett. 2008, 582, 1547–1551. [Google Scholar] [CrossRef]
- Tanaka, R.; Rothbart, M.; Oka, S.; Takabayashi, A.; Takahashi, K.; Shibata, M.; Myouga, F.; Motohashi, R.; Shinozaki, K.; Grimm, B.; et al. LIL3, a Light-Harvesting-like Protein, Plays an Essential Role in Chlorophyll and Tocopherol Biosynthesis. Proc. Natl. Acad. Sci. USA 2010, 107, 16721–16725. [Google Scholar] [CrossRef]
- Hey, D.; Rothbart, M.; Herbst, J.; Wang, P.; Müller, J.; Wittmann, D.; Gruhl, K.; Grimm, B. LIL3, a Light-Harvesting Complex Protein, Links Terpenoid and Tetrapyrrole Biosynthesis in Arabidopsis Thaliana. Plant Physiol. 2017, 174, 1037–1050. [Google Scholar] [CrossRef] [PubMed]
- Lohscheider, J.N.; Rojas-Stütz, M.C.; Rothbart, M.; Andersson, U.; Funck, D.; Mendgen, K.; Grimm, B.; Adamska, I. Altered Levels of LIL3 Isoforms in Arabidopsis Lead to Disturbed Pigment-Protein Assembly and Chlorophyll Synthesis, Chlorotic Phenotype and Impaired Photosynthetic Performance. Plant Cell Environ. 2015, 38, 2115–2127. [Google Scholar] [CrossRef] [PubMed]
- Mork-Jansson, A.; Bue, A.K.; Gargano, D.; Furnes, C.; Reisinger, V.; Arnold, J.; Kmiec, K.; Eichacker, L.A. Lil3 Assembles with Proteins Regulating Chlorophyll Synthesis in Barley. PLoS ONE 2015, 10, e0133145. [Google Scholar] [CrossRef]
- Skotnicová, P.; Staleva, M.H.; Kuznetsova, V.; Bína, D.; Konert, M.M.; Lu, S.; Polívka, T.; Sobotka, R. Plant LHC-like Proteins Show Robust Folding and Static Non-Photochemical Quenching. Nat. Commun. 2021, 12, 6890. [Google Scholar] [CrossRef] [PubMed]
- Green, B.R.; Ktihlbrandt, W. Sequence Conservation of Light-Harvesting and Stress-Response Proteins in Relation to the Three-Dimensional Molecular Structure of LHCII. Photosynth. Res. 1995, 44, 139–148. [Google Scholar] [CrossRef] [PubMed]
- Adamska, I.; Ohad, I.; Kloppstech, K. Synthesis of the Early Light-Inducible Protein Is Controlled by Blue Light and Related to Light Stress. Proc. Natl. Acad. Sci. USA 1992, 89, 2610–2613. [Google Scholar] [CrossRef]
- Heddad, M.; Norén, H.; Reiser, V.; Dunaeva, M.; Andersson, B.; Adamska, I. Differential Expression and Localization of Early Light-Induced Proteins in Arabidopsis. Plant Physiol. 2006, 142, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Adamska, I.; Roobol-Bóza, M.; Lindahl, M.; Andersson, B. Isolation of Pigment-Binding Early Light-Inducible Proteins from Pea. Eur. J. Biochem. 1999, 260, 453–460. [Google Scholar] [CrossRef]
- Adamska, I. ELIPs–Light-Induced Stress Proteins. Physiol. Plant. 1997, 100, 794–805. [Google Scholar] [CrossRef]
- Hutin, C.; Nussaume, L.; Moise, N.; Moya, I.; Kloppstech, K.; Havaux, M. Early Light-Induced Proteins Protect Arabidopsis from Photooxidative Stress. Proc. Natl. Acad. Sci. USA 2003, 100, 4921–4926. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Yang, H.; Liang, Y.; Li, X.; Oliver, M.J.; Zhang, D. Functional Aspects of Early Light-induced Protein (Elip) Genes from the Desiccation-tolerant Moss Syntrichia Caninervis. Int. J. Mol. Sci. 2020, 21, 1411. [Google Scholar] [CrossRef] [PubMed]
- Elrad, D.; Grossman, A.R. A Genome’s Eye View of the Light-Harvesting Polypeptides of Chlamydomonas reinhardtii. Curr. Genet. 2004, 45, 61–75. [Google Scholar] [CrossRef] [PubMed]
- Zones, J.M.; Blaby, I.K.; Merchant, S.S.; Umen, J.G. High-Resolution Profiling of a Synchronized Diurnal Transcriptome from Chlamydomonas reinhardtii Reveals Continuous Cell and Metabolic Differentiation. Plant Cell 2015, 27, 2743–2769. [Google Scholar] [CrossRef]
- Lerss, A.; Levy, H.; Zamirq, A. Co-Regulation of a Gene Homologous to Early Light-Induced Genes in Higher Plants and Carotene Biosynthesis in the Alga Dunaiella bardawi. J. Biol. Chem. 1991, 266, 13698–13705. [Google Scholar] [CrossRef]
- Levy, H.; Gokhman, I.; Zamir, A. Regulation and Light-Harvesting Complex Association of a Dunaliella Protein Homologous to Early Light-Induced Proteins in Higher Plants. J. Biol. Chem. 1992, 267, 18831–18836. [Google Scholar] [CrossRef] [PubMed]
- Levy, H.; Tal, T.; Shaish, A.; Zamir, A. Cbr, an Algal Homolog of Plant Early Light-Induced Proteins, Is a Putative Zeaxanthin Binding Protein. J. Biol. Chem. 1993, 268, 20892–20896. [Google Scholar] [CrossRef] [PubMed]
- Banet, G.; Pick, U.; Zamir, A. Light-Harvesting Complex II Pigments and Proteins in association with Cbr, a Homolog of Higher-Plant Early Light-Inducible proteins in the Unicellular Green Alga Dunaliella. Planta 2000, 210, 947–955. [Google Scholar] [CrossRef] [PubMed]
- Holt, T.K.; Krogmann, D.W. A Carotenoid-Protein From Cyanobacteria. Biochim. Biophys. Acta Bioenerg. 1981, 637, 408–414. [Google Scholar] [CrossRef]
- Kerfeld, C.A.; Sawaya, M.R.; Brahmandam, V.; Cascio, D.; Ho, K.K.; Trevithick-Sutton, C.C.; Krogmann, D.W.; Yeates, T.O. The Crystal Structure of a Cyanobacterial Water-Soluble Carotenoid Binding Protein. Structure 2003, 11, 55–65. [Google Scholar] [CrossRef]
- Wilson, A.; Ajlani, G.; Verbavatz, J.M.; Vass, I.; Kerfeld, C.A.; Kirilovsky, D. A Soluble Carotenoid Protein Involved in Phycobilisome-Related Energy Dissipation in Cyanobacteria. Plant Cell 2006, 18, 992–1007. [Google Scholar] [CrossRef]
- Sedoud, A.; López-Igual, R.; Rehman, A.U.; Wilson, A.; Perreau, F.; Boulay, C.; Vass, I.; Krieger-Liszkay, A.; Kirilovsky, D. The Cyanobacterial Photoactive Orange Carotenoid Protein Is an Excellent Singlet Oxygen Quencher. Plant Cell 2014, 26, 1781–1791. [Google Scholar] [CrossRef] [PubMed]
- Kirilovsky, D.; Kerfeld, C.A. Cyanobacterial Photoprotection by the Orange Carotenoid Protein. Nat. Plants 2016, 2, 16180. [Google Scholar] [CrossRef] [PubMed]
- Lé Wilson, A.; Punginelli, C.; Gall, A.; Bonetti, C.; Alexandre, M.; Routaboul, J.-M.; Kerfeld, C.A.; van Grondelle, R.; Robert, B.; Kennis, J.T.M.; et al. A Photoactive Carotenoid Protein Acting as Light Intensity Sensor. Proc. Natl. Acad. Sci. USA 2008, 105, 12075–12080. [Google Scholar] [CrossRef] [PubMed]
- Yakushiji, E.; Uchino, K.; Sugimura, Y.; Shiratori, I.; Takamiya, F. Isolation of Water-Soluble Chlorophyll Protein from the Leaves of Chenopodium album. Biochim. Biophys. Acta 1963, 75, 293–298. [Google Scholar] [CrossRef]
- Horigome, D.; Satoh, H.; Itoh, N.; Mitsunaga, K.; Oonishi, I.; Nakagawa, A.; Uchida, A. Structural Mechanism and Photoprotective Function of Water-Soluble Chlorophyll-Binding Protein. J. Biol. Chem. 2007, 282, 6525–6531. [Google Scholar] [CrossRef]
- Agostini, A.; Palm, D.M.; Schmitt, F.J.; Albertini, M.; di Valentin, M.; Paulsen, H.; Carbonera, D. An Unusual Role for the Phytyl Chains in the Photoprotection of the Chlorophylls Bound to Water-Soluble Chlorophyll-Binding Proteins. Sci. Rep. 2017, 7, 7504. [Google Scholar] [CrossRef] [PubMed]
- Palm, D.M.; Agostini, A.; Pohland, A.C.; Werwie, M.; Jaenicke, E.; Paulsen, H. Stability of Water-Soluble Chlorophyll Protein (WSCP) Depends on Phytyl Conformation. ACS Omega 2019, 4, 7971–7979. [Google Scholar] [CrossRef] [PubMed]
- Damaraju, S.; Schlede, S.; Eckhardt, U.; Lokstein, H.; Grimm, B. Functions of the Water Soluble Chlorophyll-Binding Protein in Plants. J. Plant Physiol. 2011, 168, 1444–1451. [Google Scholar] [CrossRef]
- Evans, J.R. Improving Photosynthesis. Plant Physiol. 2013, 162, 1780–1793. [Google Scholar] [CrossRef]
- Nowicka, B.; Ciura, J.; Szymańska, R.; Kruk, J. Improving Photosynthesis, Plant Productivity and Abiotic Stress Tolerance – Current Trends and Future Perspectives. J. Plant Physiol. 2018, 231, 415–433. [Google Scholar] [CrossRef]
- Vecchi, V.; Barera, S.; Bassi, R.; Dall’osto, L. Potential and Challenges of Improving Photosynthesis in Algae. Plants 2020, 9, 67. [Google Scholar] [CrossRef] [PubMed]
- Głowacka, K.; Kromdijk, J.; Kucera, K.; Xie, J.; Cavanagh, A.P.; Leonelli, L.; Leakey, A.D.B.; Ort, D.R.; Niyogi, K.K.; Long, S.P. Photosystem II Subunit S Overexpression Increases the Efficiency of Water Use in a Field-Grown Crop. Nat. Commun. 2018, 9. [Google Scholar] [CrossRef] [PubMed]
- Hubbart, S.; Smillie, I.R.A.; Heatley, M.; Swarup, R.; Foo, C.C.; Zhao, L.; Murchie, E.H. Enhanced Thylakoid Photoprotection Can Increase Yield and Canopy Radiation Use Efficiency in Rice. Commun. Biol. 2018, 1, 22. [Google Scholar] [CrossRef] [PubMed]
- Perozeni, F.; Stella, G.R.; Ballottari, M. LHCSR Expression under HSP70/RBCS2 Promoter as a Strategy to Increase Productivity in Microalgae. Int. J. Mol. Sci. 2018, 19, 155. [Google Scholar] [CrossRef] [PubMed]
- Zhuo, C.; Cai, J.; Guo, Z. Overexpression of Early Light-Induced Protein (ELIP) Gene from Medicago Sativa Ssp. Falcata Increases Tolerance to Abiotic Stresses. J. Agron. 2013, 105, 1433–1440. [Google Scholar] [CrossRef]
- Liu, B.; Zhao, F.; Zhou, H.; Xia, Y.; Wang, X. Photoprotection Conferring Plant Tolerance to Freezing Stress through Rescuing Photosystem in Evergreen Rhododendron. Plant Cell Environ. 2022, 45, 2093–2108. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Z.; He, B.; Guo, M.L.; Xie, X.; Huan, L.; Zhang, B.; Shao, Z.; Wang, G. Overexpression of OHPs in Neopyropia Yezoensis (Rhodophyta) Reveals Their Possible Physiological Roles. Algal Res. 2022, 64, 102714. [Google Scholar] [CrossRef]
- Hey, D.; Grimm, B. Requirement of ONE-HELIX PROTEIN 1 (OHP1) in Early Arabidopsis Seedling Development and under High Light Intensity. Plant Signal. Behav. 2018, 13, e1550317. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).