
Influence of Butyrate on Impaired Gene Expression in Colon from Patients with High Blood Pressure

 , ,
, ,
Abstract
:1. Introduction
2. Results
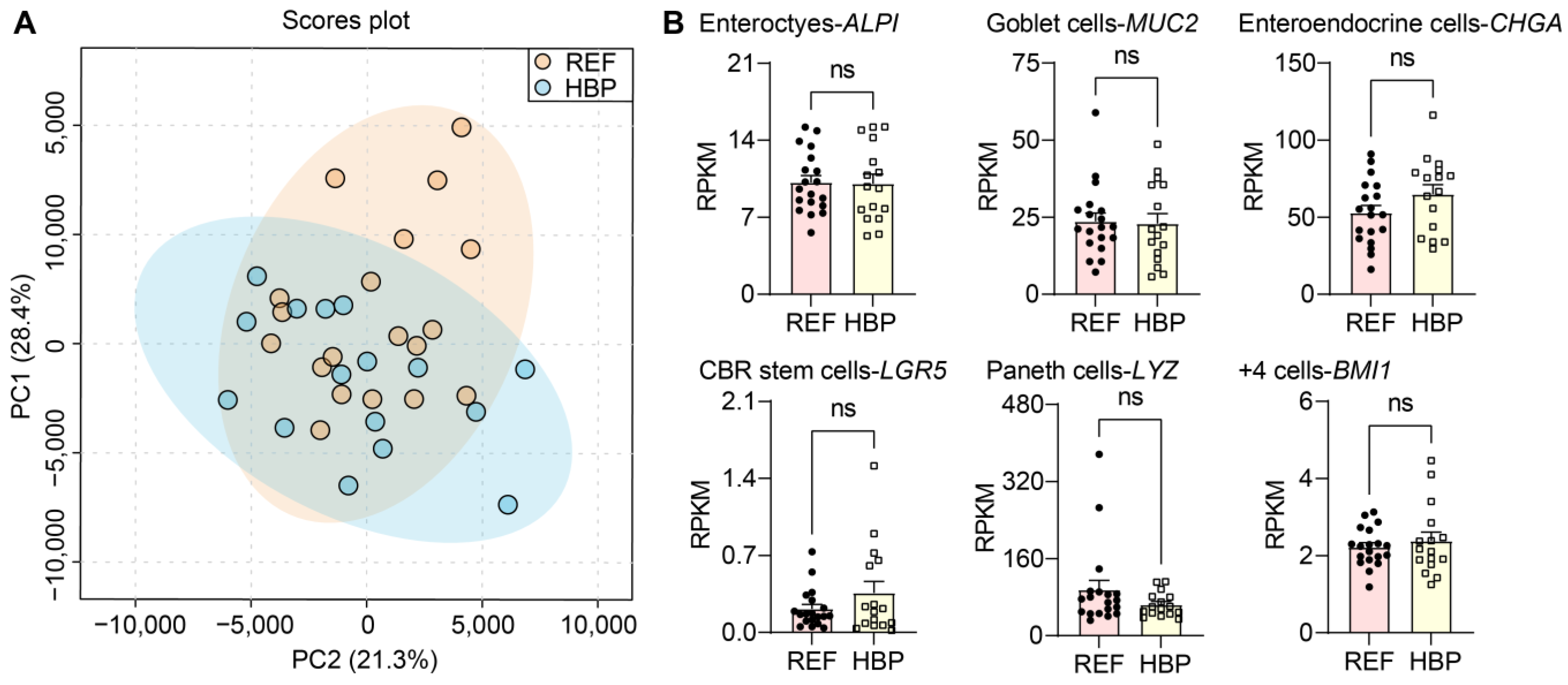
2.1. Characterization of Subjects and Colonic Tissues
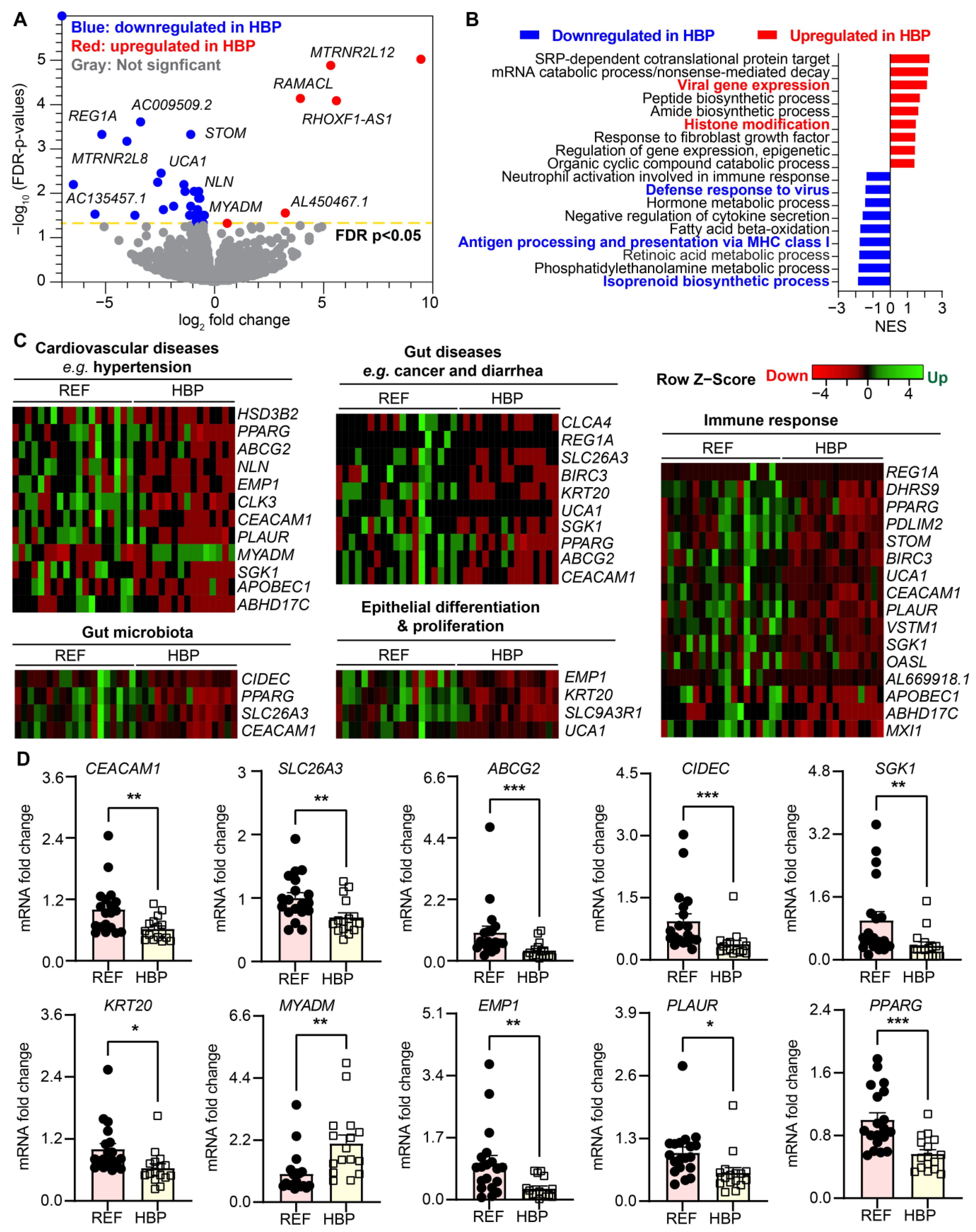
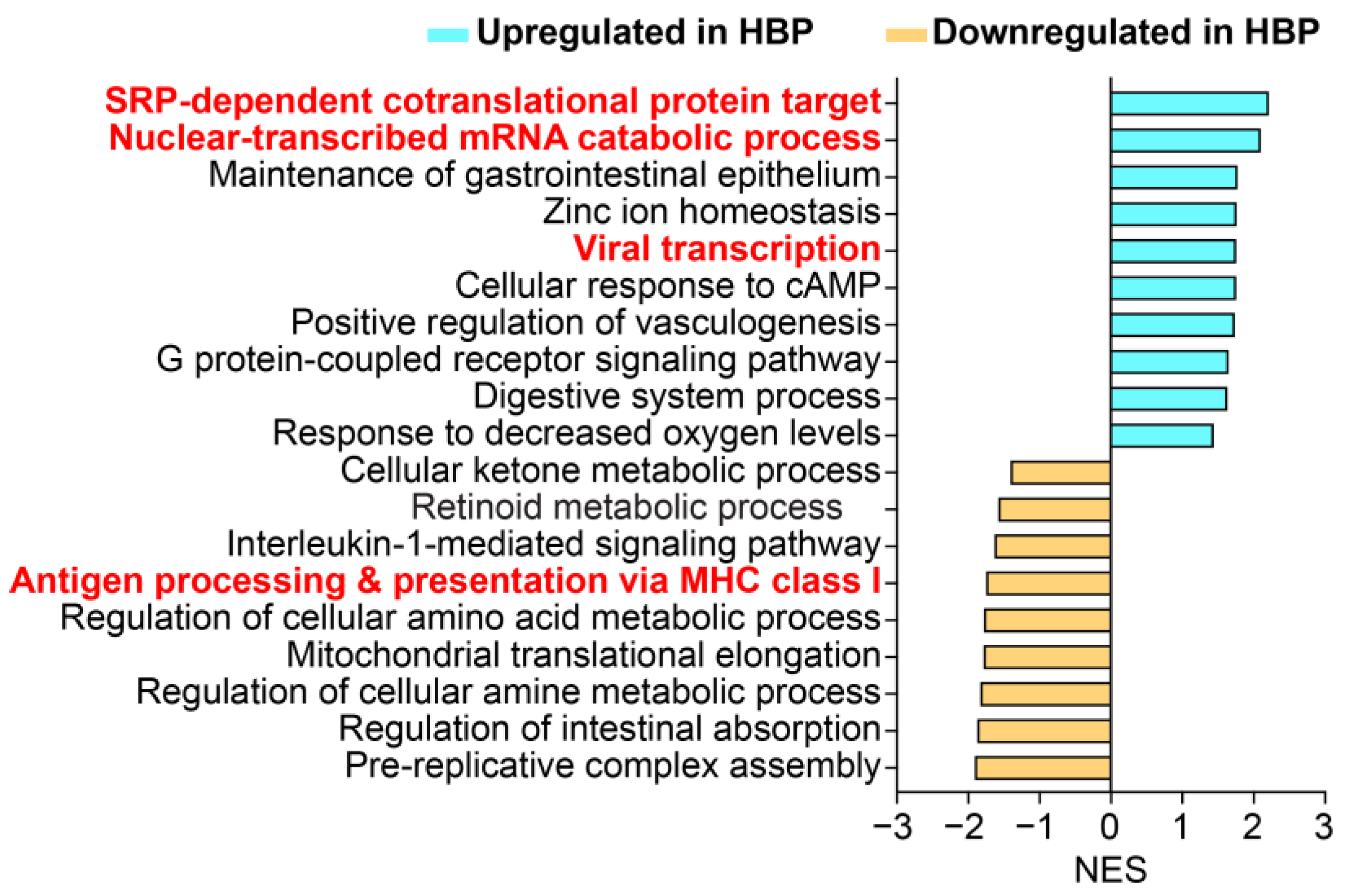
2.2. HBP Patients Have Distinct Colon Transcriptome Signatures
2.3. Identification of Potential Novel Drug Targets
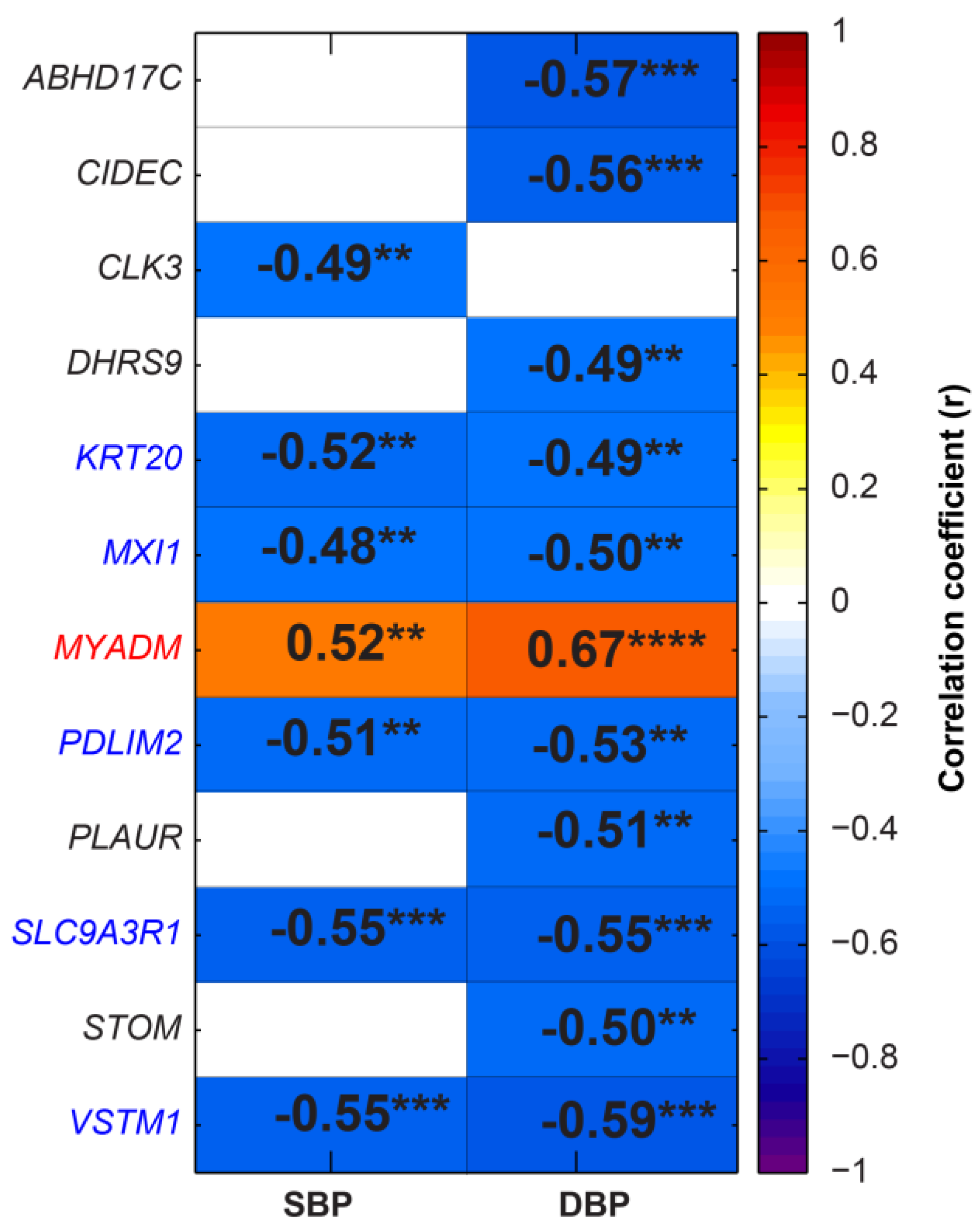
2.4. Specific Differentially Expressed Genes (DEGs) Correlated with SBP and DBP
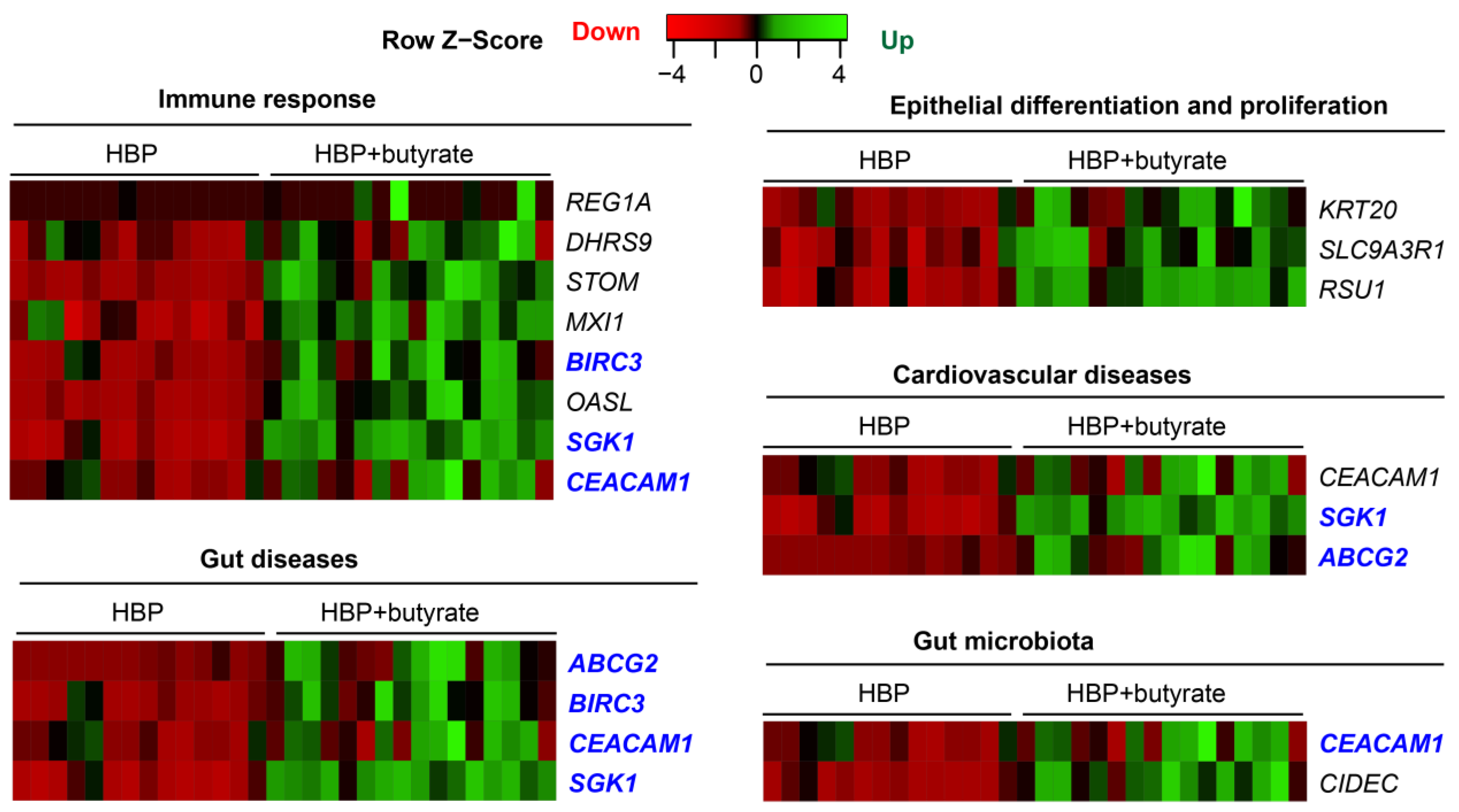
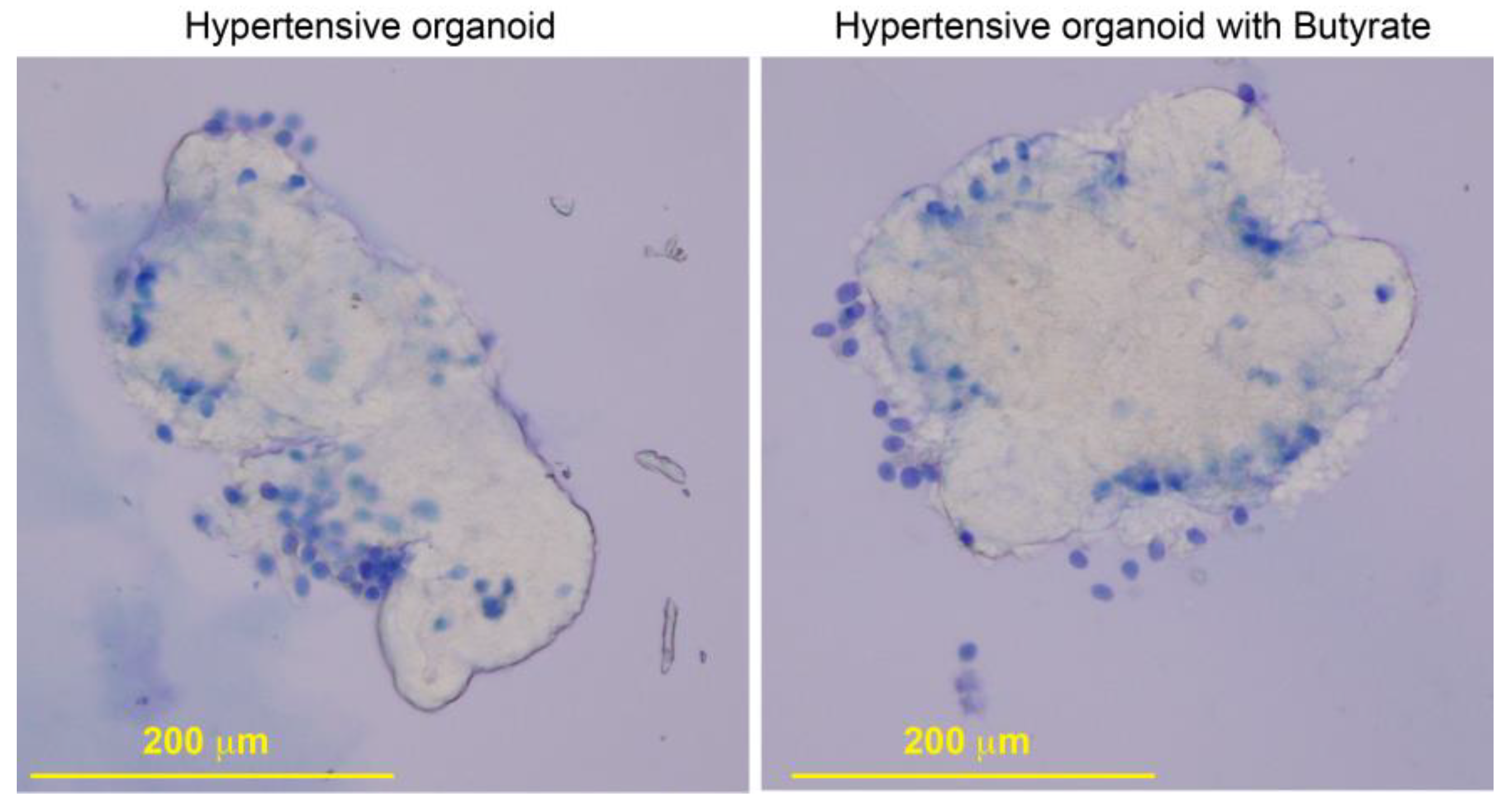
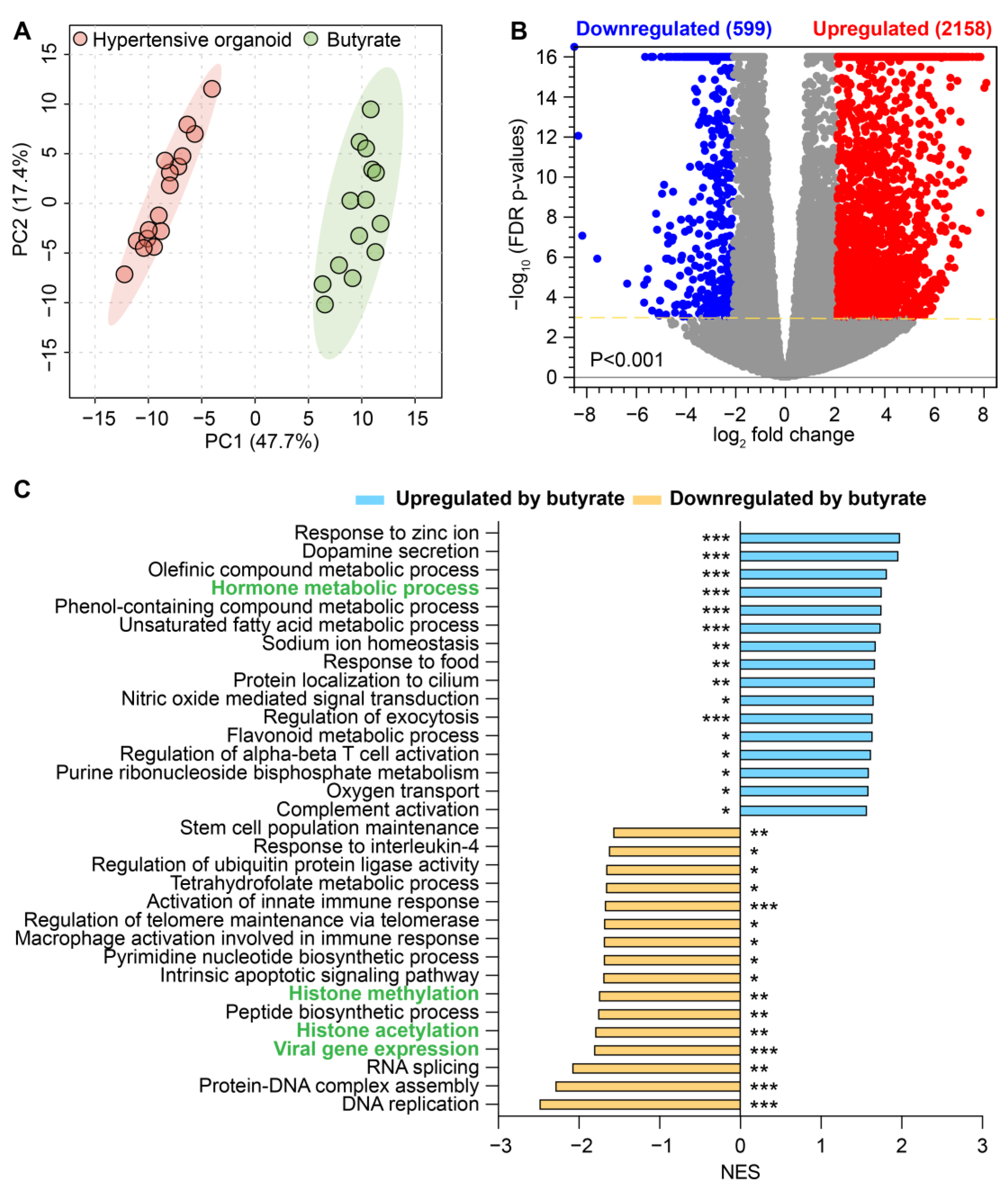
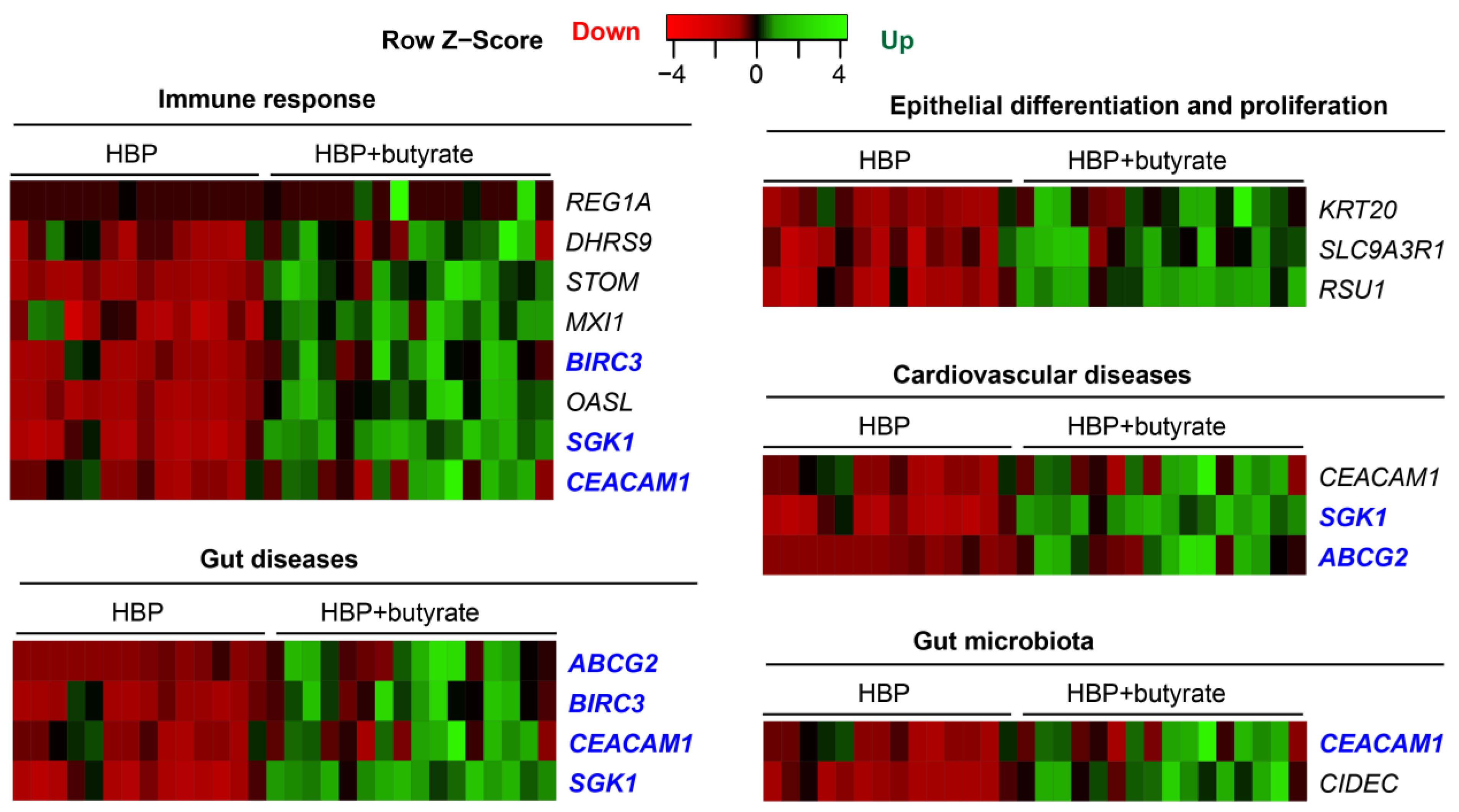
2.5. Butyrate Treatment Rescued Impaired Gene Expression in Organoids from HBP Subjects
3. Discussion
4. Materials and Methods
4.1. Study Cohort
4.2. RNA-seq
4.3. Quantitative PCR (qPCR)
4.4. Human Colonic 3D Organoid Culture
4.5. Correlation Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yang, T.; Santisteban, M.M.; Rodriguez, V.; Li, E.; Ahmari, N.; Carvajal, J.M.; Zadeh, M.; Gong, M.; Qi, Y.; Zubcevic, J.; et al. Gut dysbiosis is linked to hypertension. Hypertension 2015, 65, 1331–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santisteban, M.M.; Kim, S.; Pepine, C.J.; Raizada, M.K. Brain–gut–bone marrow axis: Implications for hypertension and related therapeutics. Circ. Res. 2016, 118, 1327–1336. [Google Scholar] [CrossRef] [PubMed]
- Santisteban, M.M.; Qi, Y.; Zubcevic, J.; Kim, S.; Yang, T.; Shenoy, V.; Cole-Jeffrey, C.T.; Lobaton, G.O.; Stewart, D.C.; Rubiano, A.; et al. Hypertension-linked pathophysiological alterations in the gut. Circ. Res. 2017, 120, 312–323. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.; Goel, R.; Kumar, A.; Qi, Y.; Lobaton, G.; Hosaka, K.; Mohammed, M.; Handberg, E.M.; Richards, E.M.; Pepine, C.J.; et al. Imbalance of gut microbiome and intestinal epithelial barrier dysfunction in patients with high blood pressure. Clin. Sci. 2018, 132, 701–718. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Raizada, M.K.; Richards, E.M. Gut–brain–bone marrow axis in hypertension. Curr. Opin. Nephrol. Hypertens. 2021, 30, 159–165. [Google Scholar] [CrossRef]
- Richards, E.M.; Li, J.; Stevens, B.R.; Pepine, C.J.; Raizada, M.K. Gut Microbiome and Neuroinflammation in Hypertension. Circ. Res. 2022, 130, 401–417. [Google Scholar] [CrossRef]
- Li, J.; Zhao, F.; Wang, Y.; Chen, J.; Tao, J.; Tian, G.; Wu, S.; Liu, W.; Cui, Q.; Geng, B. Gut microbiota dysbiosis contributes to the development of hypertension. Microbiome 2017, 5, 14. [Google Scholar] [CrossRef] [Green Version]
- Toral, M.; Robles-Vera, I.; De la Visitacion, N.; Romero, M.; Yang, T.; Sánchez, M.; Gómez-Guzmán, M.; Jiménez, R.; Raizada, M.K.; Duarte, J. Critical role of the interaction gut microbiota–sympathetic nervous system in the regulation of blood pressure. Front. Physiol. 2019, 10, 231. [Google Scholar] [CrossRef] [Green Version]
- Zheng, D.; Liwinski, T.; Elinav, E. Interaction between microbiota and immunity in health and disease. Cell Res. 2020, 30, 492–506. [Google Scholar] [CrossRef]
- Stevens, B.R.; Goel, R.; Seungbum, K.; Richards, E.M.; Holbert, R.C.; Pepine, C.J.; Raizada, M.K. Increased human intestinal barrier permeability plasma biomarkers zonulin and FABP2 correlated with plasma LPS and altered gut microbiome in anxiety or depression. Gut 2018, 67, 1555–1557. [Google Scholar] [CrossRef]
- Bartolomaeus, H.; Balogh, A.; Yakoub, M.; Homann, S.; Markó, L.; Höges, S.; Tsvetkov, D.; Krannich, A.; Wundersitz, S.; Avery, E.G.; et al. Short-chain fatty acid propionate protects from hypertensive cardiovascular damage. Circulation 2019, 139, 1407–1421. [Google Scholar] [CrossRef] [PubMed]
- Cai, Z.; Klein, T.; Tu, L.; Geenen, L.; Tian, S.; Van Der Ley, C.; Van Faassen, M.; Kema, I.; Van Den Bosch, A.; De Rijke, J.; et al. Different tryptophan-kynurenine metabolism profiles in human pulmonary arterial hypertension and animal models of pulmonary hypertension. Eur. Heart J. 2020, 41, ehaa946.3814. [Google Scholar] [CrossRef]
- Kaye, D.M.; Shihata, W.A.; Jama, H.A.; Tsyganov, K.; Ziemann, M.; Kiriazis, H.; Horlock, D.; Vijay, A.; Giam, B.; Vinh, A.; et al. Deficiency of prebiotic fiber and insufficient signaling through gut metabolite-sensing receptors leads to cardiovascular disease. Circulation 2020, 141, 1393–1403. [Google Scholar] [CrossRef]
- Kong, C.-Y.; Li, Z.-M.; Mao, Y.-Q.; Chen, H.-L.; Hu, W.; Han, B.; Wang, L.-S. Probiotic yogurt blunts the increase of blood pressure in spontaneously hypertensive rats via remodeling of the gut microbiota. Food Funct. 2021, 12, 9773–9783. [Google Scholar] [CrossRef] [PubMed]
- Robles-Vera, I.; Toral, M.; Romero, M.; Jiménez, R.; Sánchez, M.; Pérez-Vizcaíno, F.; Duarte, J. Antihypertensive effects of probiotics. Curr. Hypertens. Rep. 2017, 19, 26. [Google Scholar] [CrossRef] [PubMed]
- Yang, T.; Li, H.; Oliveira, A.C.; Goel, R.; Richards, E.M.; Pepine, C.J.; Raizada, M.K. Transcriptomic signature of gut microbiome-contacting cells in colon of spontaneously hypertensive rats. Physiol. Genom. 2020, 52, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Richards, E.M.; Handberg, E.M.; Pepine, C.J.; Raizada, M.K. Distinct Gene Expression Profiles in Colonic Organoids from Normotensive and the Spontaneously Hypertensive Rats. Cells 2021, 10, 1523. [Google Scholar] [CrossRef]
- Engevik, M.A.; Aihara, E.; Montrose, M.H.; Shull, G.E.; Hassett, D.J.; Worrell, R.T. Loss of NHE3 alters gut microbiota composition and influences Bacteroides thetaiotaomicron growth. Am. J. Physiol. Gastrointest. Liver Physiol. 2013, 305, G697–G711. [Google Scholar] [CrossRef] [Green Version]
- Pluznick, J.L.; Protzko, R.J.; Gevorgyan, H.; Peterlin, Z.; Sipos, A.; Han, J.; Brunet, I.; Wan, L.-X.; Rey, F.; Wang, T.; et al. Olfactory receptor responding to gut microbiota-derived signals plays a role in renin secretion and blood pressure regulation. Proc. Natl. Acad. Sci. USA 2013, 110, 4410–4415. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Richards, E.M.; Handberg, E.M.; Pepine, C.J.; Raizada, M.K. Butyrate regulates COVID-19–relevant genes in gut epithelial organoids from normotensive rats. Hypertension 2021, 77, e13–e16. [Google Scholar] [CrossRef]
- Zhang, C.; Jin, D.-D.; Wang, X.-Y.; Lou, L.; Yang, J. Key Enzymes for the Mevalonate Pathway in the Cardiovascular System. J. Cardiovasc. Pharmacol. 2021, 77, 142–152. [Google Scholar] [CrossRef] [PubMed]
- Shimodaira, M.; Nakayama, T.; Sato, N.; Aoi, N.; Sato, M.; Izumi, Y.; Soma, M.; Matsumoto, K. Association of HSD3B1 and HSD3B2 gene polymorphisms with essential hypertension, aldosterone level, and left ventricular structure. Eur. J. Endocrinol. 2010, 163, 671–680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Liu, Y.; Jiang, C.-J.; Chen, Y.-M.; Li, H.; Liu, Q.-A. Calcium-activated chloride channel A4 (CLCA4) plays inhibitory roles in invasion and migration through suppressing epithelial-mesenchymal transition via PI3K/AKT signaling in colorectal cancer. Med. Sci. Monit. Int. Med. J. Exp. Clin. Res. 2019, 25, 4176. [Google Scholar] [CrossRef] [PubMed]
- Xiao, F.; Yu, Q.; Li, J.; Johansson, M.; Singh, A.; Xia, W.; Riederer, B.; Engelhardt, R.; Montrose, M.; Soleimani, M. Slc26a3 deficiency is associated with loss of colonic HCO 3− secretion, absence of a firm mucus layer and barrier impairment in mice. Acta Physiol. 2014, 211, 161–175. [Google Scholar] [CrossRef] [PubMed]
- Freshour, S.L.; Kiwala, S.; Cotto, K.C.; Coffman, A.C.; McMichael, J.F.; Song, J.J.; Griffith, M.; Griffith, O.L.; Wagner, A.H. Integration of the Drug–Gene Interaction Database (DGIdb 40) with open crowdsource efforts. Nucleic Acids Res. 2021, 49, D1144–D1151. [Google Scholar] [CrossRef]
- Dubey, R.K.; Oparil, S.; Imthurn, B.; Jackson, E.K. Sex hormones and hypertension. Cardiovasc. Res. 2002, 53, 688–708. [Google Scholar] [CrossRef] [Green Version]
- Choi, U.Y.; Kang, J.-S.; Hwang, Y.S.; Kim, Y.-J. Oligoadenylate synthase-like (OASL) proteins: Dual functions and associations with diseases. Exp. Mol. Med. 2015, 47, e144. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Ingle, H.; Mishra, S.; Mahla, R.; Kumar, A.; Kawai, T.; Akira, S.; Takaoka, A.; Raut, A.; Kumar, H. IPS-1 differentially induces TRAIL, BCL2, BIRC3 and PRKCE in type I interferons-dependent and-independent anticancer activity. Cell Death Dis. 2015, 6, e1758. [Google Scholar] [CrossRef] [Green Version]
- Napoli, J.L. 17β-Hydroxysteroid dehydrogenase type 9 and other short-chain dehydrogenases/reductases that catalyze retinoid, 17β-and 3α-hydroxysteroid metabolism. Mol. Cell. Endocrinol. 2001, 171, 103–109. [Google Scholar] [CrossRef]
- Riquelme, P.; Hutchinson, J.A. Novel molecules mediate specialized functions of human regulatory macrophages. Curr. Opin. Organ Transplant. 2018, 23, 533–537. [Google Scholar] [CrossRef]
- Ahmadian, M.; Suh, J.M.; Hah, N.; Liddle, C.; Atkins, A.R.; Downes, M.; Evans, R.M. PPARγ signaling and metabolism: The good, the bad and the future. Nat. Med. 2013, 19, 557–566. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lefterova, M.I.; Haakonsson, A.K.; Lazar, M.A.; Mandrup, S. PPARγ and the global map of adipogenesis and beyond. Trends Endocrinol. Metab. 2014, 25, 293–302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, S.; Sigmund, C.D. PPARγ and RhoBTB1 in hypertension. Curr. Opin. Nephrol. Hypertens. 2020, 29, 161. [Google Scholar] [CrossRef] [PubMed]
- Gao, Z.; Yin, J.; Zhang, J.; Ward, R.E.; Martin, R.J.; Lefevre, M.; Cefalu, W.T.; Ye, J. Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 2009, 58, 1509–1517. [Google Scholar] [CrossRef] [Green Version]
- Den Besten, G.; Van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.; Wang, L.; Han, S.; Wang, Z.; Ni, Y.; Geng, Y.; Zhang, L. PPARG Silencing Improves Blood Pressure Control and Alleviates Renal Damage by Modulating RAS Circadian Rhythm in Hypertensive Rats. Ann. Clin. Lab. Sci. 2022, 52, 452–461. [Google Scholar]
- Lang, F.; Huang, D.Y.; Vallon, V. SGK, renal function and hypertension. J. Nephrol. 2010, 23, S124. [Google Scholar]
- Salvi, E.; Wang, Z.; Rizzi, F.; Gong, Y.; McDonough, C.W.; Padmanabhan, S.; Hiltunen, T.P.; Lanzani, C.; Zaninello, R.; Chittani, M.; et al. Genome-wide and gene-based meta-analyses identify novel loci influencing blood pressure response to hydrochlorothiazide. Hypertension 2017, 69, 51–59. [Google Scholar] [CrossRef] [Green Version]
- Domínguez, C.J.; Tocchetti, G.N.; Rigalli, J.P.; Mottino, A.D. Acute regulation of apical ABC transporters in the gut. Potential influence on drug bioavailability. Pharmacol. Res. 2021, 163, 105251. [Google Scholar] [CrossRef]
- Jiang, Y.-D.; Chuang, L.-M.; Wu, H.-P.; Shiau, S.-J.; Wang, C.-H.; Lee, Y.-J.; Juang, J.-H.; Lin, B.J.; Tai, T.-Y. Assessment of the function and effect of diabetes education programs in Taiwan. Diabetes Res. Clin. Pract. 1999, 46, 177–182. [Google Scholar] [CrossRef]
- Landin, K.; Tengborn, L.; Smith, U. Elevated fibrinogen and plasminogen activator inhibitor (PAI-1) in hypertension are related to metabolic risk factors for cardiovascular disease. J. Intern. Med. 1990, 227, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Eugen-Olsen, J.; Andersen, O.; Linneberg, A.; Ladelund, S.; Hansen, T.; Langkilde, A.; Petersen, J.; Pielak, T.; Møller, L.; Jeppesen, J.; et al. Circulating soluble urokinase plasminogen activator receptor predicts cancer, cardiovascular disease, diabetes and mortality in the general population. J. Intern. Med. 2010, 268, 296–308. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Q.; Cadrin, M.; Herrmann, H.; Chen, C.-H.; Chalkley, R.J.; Burlingame, A.L.; Omary, M.B. Keratin 20 serine 13 phosphorylation is a stress and intestinal goblet cell marker. J. Biol. Chem. 2006, 281, 16453–16461. [Google Scholar] [CrossRef] [Green Version]
- Aranda, J.F.; Reglero-Real, N.; Marcos-Ramiro, B.; Ruiz-Sáenz, A.; Fernández-Martín, L.; Bernabé-Rubio, M.; Kremer, L.; Ridley, A.J.; Correas, I.; Alonso, M.A.; et al. MYADM controls endothelial barrier function through ERM-dependent regulation of ICAM-1 expression. Mol. Biol. Cell 2013, 24, 483–494. [Google Scholar] [CrossRef] [Green Version]
- Huan, T.; Esko, T.; Peters, M.J.; Pilling, L.C.; Schramm, K.; Schurmann, C.; Chen, B.H.; Liu, C.; Joehanes, R.; Johnson, A.D. A meta-analysis of gene expression signatures of blood pressure and hypertension. PLoS Genet. 2015, 11, e1005035. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zeller, T.; Schurmann, C.; Schramm, K.; Müller, C.; Kwon, S.; Wild, P.S.; Teumer, A.; Herrington, D.; Schillert, A.; Iacoviello, L.; et al. Transcriptome-wide analysis identifies novel associations with blood pressure. Hypertension 2017, 70, 743–750. [Google Scholar] [CrossRef]
- Bai, Y.; Wang, J.; Chen, Y.; Lv, T.; Wang, X.; Liu, C.; Xue, H.; He, K.; Sun, L. The miR-182/Myadm axis regulates hypoxia-induced pulmonary hypertension by balancing the BMP-and TGF-β-signalling pathways in an SMC/EC-crosstalk-associated manner. Basic Res. Cardiol. 2021, 116, 53. [Google Scholar] [CrossRef]
- Gray-Owen, S.D.; Blumberg, R.S. CEACAM1: Contact-dependent control of immunity. Nat. Rev. Immunol. 2006, 6, 433–446. [Google Scholar] [CrossRef]
- Li, C.; Culver, S.A.; Quadri, S.; Ledford, K.L.; Al-Share, Q.Y.; Ghadieh, H.E.; Najjar, S.M.; Siragy, H.M. High-fat diet amplifies renal renin angiotensin system expression, blood pressure elevation, and renal dysfunction caused by Ceacam1 null deletion. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E802–E810. [Google Scholar] [CrossRef] [Green Version]
- De la Cuesta-Zuluaga, J.; Mueller, N.T.; Álvarez-Quintero, R.; Velásquez-Mejía, E.P.; Sierra, J.A.; Corrales-Agudelo, V.; Carmona, J.A.; Abad, J.M.; Escobar, J.S. Higher fecal short-chain fatty acid levels are associated with gut microbiome dysbiosis, obesity, hypertension and cardiometabolic disease risk factors. Nutrients 2018, 11, 51. [Google Scholar] [CrossRef] [Green Version]
- Huart, J.; Leenders, J.; Taminiau, B.; Descy, J.; Saint-Remy, A.; Daube, G.; Krzesinski, J.-M.; Melin, P.; De Tullio, P.; Jouret, F. Gut microbiota and fecal levels of short-chain fatty acids differ upon 24-hour blood pressure levels in men. Hypertension 2019, 74, 1005–1013. [Google Scholar] [CrossRef] [PubMed]
- Tilves, C.; Yeh, H.C.; Maruthur, N.; Juraschek, S.P.; Miller, E.; White, K.; Appel, L.J.; Mueller, N.T. Increases in circulating and fecal butyrate are associated with reduced blood pressure and hypertension: Results From the SPIRIT trial. J. Am. Heart Assoc. 2022, 11, e024763. [Google Scholar] [CrossRef] [PubMed]
- Ge, S.X.; Son, E.W.; Yao, R. iDEP: An integrated web application for differential expression and pathway analysis of RNA-Seq data. BMC Bioinform. 2018, 19, 534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Babicki, S.; Arndt, D.; Marcu, A.; Liang, Y.; Grant, J.R.; Maciejewski, A.; Wishart, D.S. Heatmapper: Web-enabled heat mapping for all. Nucleic Acids Res. 2016, 44, W147–W153. [Google Scholar] [CrossRef]
- Li, J.; Stevens, B.R.; Richards, E.M.; Raizada, M.K. SARS-CoV-2 receptor ACE2 (Angiotensin-Converting Enzyme 2) is upregulated in colonic organoids from hypertensive rats. Hypertension 2020, 76, e26–e28. [Google Scholar] [CrossRef] [PubMed]
- Peng, L.; Li, Z.-R.; Green, R.S.; Holzman, I.R.; Lin, J. Butyrate enhances the intestinal barrier by facilitating tight junction assembly via activation of AMP-activated protein kinase in Caco-2 cell monolayers. J. Nutr. 2009, 139, 1619–1625. [Google Scholar] [CrossRef] [Green Version]
- Ruemmele, F.; Schwartz, S.; Seidman, E.; Dionne, S.; Levy, E.; Lentze, M. Butyrate induced Caco-2 cell apoptosis is mediated via the mitochondrial pathway. Gut 2003, 52, 94–100. [Google Scholar] [CrossRef] [Green Version]
- Ranganna, K.; Yatsu, F.M.; Hayes, B.E.; Milton, S.G.; Jayakumar, A. Butyrate inhibits proliferation-induced proliferating cell nuclear antigen expression (PCNA) in rat vascular smooth muscle cells. Mol. Cell. Biochem. 2000, 205, 149–161. [Google Scholar] [CrossRef]
- Nagpal, R.; Wang, S.; Solberg Woods, L.C.; Seshie, O.; Chung, S.T.; Shively, C.A.; Register, T.C.; Craft, S.; McClain, D.A.; Yadav, H. Comparative microbiome signatures and short-chain fatty acids in mouse, rat, non-human primate, and human feces. Front. Microbiol. 2018, 9, 2897. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | RBP-Mean (95% CI) | HBP-Mean (95% CI) | p Value |
| Age (years) | 56 (52–60) | 61 (57–65) | 0.07 |
| Men | 6 (32) | 8 (50) | 0.32 |
| Systolic blood pressure (mmHg) | 111 (104–118) | 156 (146–165) | <0.0001 |
| Diastolic blood pressure (mmHg) | 68 (62–73) | 88 (84–91) | <0.0001 |
| Variable | RBP-Number (%) | HBP-Number (%) | p Value |
| White Race | 15 (79) | 10 (62.5) | 0.45 |
| Body mass index (kg/m2) | 29.0 (27–31) | 30.1 (28–33) | 0.32 |
| Smoker | 4 (21) | 4 (25) | >0.99 |
| Diabetes | 3 (16) | 4 (25) | 0.68 |
| Hyperlipidemia | 5 (26) | 1 (6) | 0.19 |
| Coronary artery disease | 0 | 0 | >0.99 |
| Peripheral Artery disease | 0 | 0 | >0.99 |
| Cerebrovascular disease | 1 (5) | 0 | >0.99 |
| Chronic kidney disease | 2 (11) | 0 | 0.49 |
| Number of Anti-Hypertensive Drug | RBP-Number (%) | HBP-Number (%) | p Value |
| 0 | 12 (63) | 8 (50) | 0.30 |
| 1 | 6 (32) | 5 (31) | >0.99 |
| 2 | 1 (5) | 2 (12) | 0.58 |
| 3 | 0 | 1 (6) | 0.46 |
| ≥4 | 0 | 0 | >0.99 |
| Type of Anti-Hypertensive Drug | RBP-Number (%) | HBP-Number (%) | p Value |
| RAS blockers (ACEIs or ARBs) | 2 (11) | 6 (37) | 0.11 |
| Aldosterone antagonists | 0 | 0 | >0.99 |
| Beta blockers | 1 (5) | 2 (12) | 0.58 |
| Calcium channel blockers | 3 (16) | 3 (19) | >0.99 |
| Diuretics | 2 (11) | 1 (6) | >0.99 |
| Alpha blockers | 0 | 0 | >0.99 |
| Arteriole vasodilators | 0 | 0 | >0.99 |
| Drug or Antibody | Gene | Fold Change | FDR P | Mechanism of Interaction |
|---|---|---|---|---|
| Mbx-2044 | PPARG | –1.43 | 0.0486 | Peroxisome proliferator-activated |
| Treprostinil | receptor gamma modulator | |||
| Aldosterone | SGK1 | –2.65 | 0.0067 | Activate serum/glucocorticoid |
| regulated kinase 1 | ||||
| AT-406 | BIRC3 | –1.85 | 0.0094 | cIAP1/cIAP2 inhibitor |
| Trilostane | HSD3B2 | –3.55 | 0.0199 | 3-beta-hydroxysteroid dehydrogenase/ |
| isomerase type II inhibitor | ||||
| Arcitumomab | CEACAM1 | –1.57 | 0.0327 | Antigen monoclonal antibody |
| Urokinase | PLAUR | –1.66 | 0.0319 | Plasminogen activator |
| Chembl488910 | ABCG2 | –1.78 | 0.0431 | Resistance protein inhibitors |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Richards, E.M.; Handberg, E.M.; Pepine, C.J.; Alakrad, E.; Forsmark, C.E.; Raizada, M.K. Influence of Butyrate on Impaired Gene Expression in Colon from Patients with High Blood Pressure. Int. J. Mol. Sci. 2023, 24, 2650. https://doi.org/10.3390/ijms24032650
Li J, Richards EM, Handberg EM, Pepine CJ, Alakrad E, Forsmark CE, Raizada MK. Influence of Butyrate on Impaired Gene Expression in Colon from Patients with High Blood Pressure. International Journal of Molecular Sciences. 2023; 24(3):2650. https://doi.org/10.3390/ijms24032650
Chicago/Turabian StyleLi, Jing, Elaine M. Richards, Eileen M. Handberg, Carl J. Pepine, Eyad Alakrad, Chris E. Forsmark, and Mohan K. Raizada. 2023. "Influence of Butyrate on Impaired Gene Expression in Colon from Patients with High Blood Pressure" International Journal of Molecular Sciences 24, no. 3: 2650. https://doi.org/10.3390/ijms24032650
APA StyleLi, J., Richards, E. M., Handberg, E. M., Pepine, C. J., Alakrad, E., Forsmark, C. E., & Raizada, M. K. (2023). Influence of Butyrate on Impaired Gene Expression in Colon from Patients with High Blood Pressure. International Journal of Molecular Sciences, 24(3), 2650. https://doi.org/10.3390/ijms24032650