
Differential Roles of Cystathionine Gamma-Lyase and Mercaptopyruvate Sulfurtransferase in Hapten-Induced Colitis and Contact Dermatitis in Mice



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
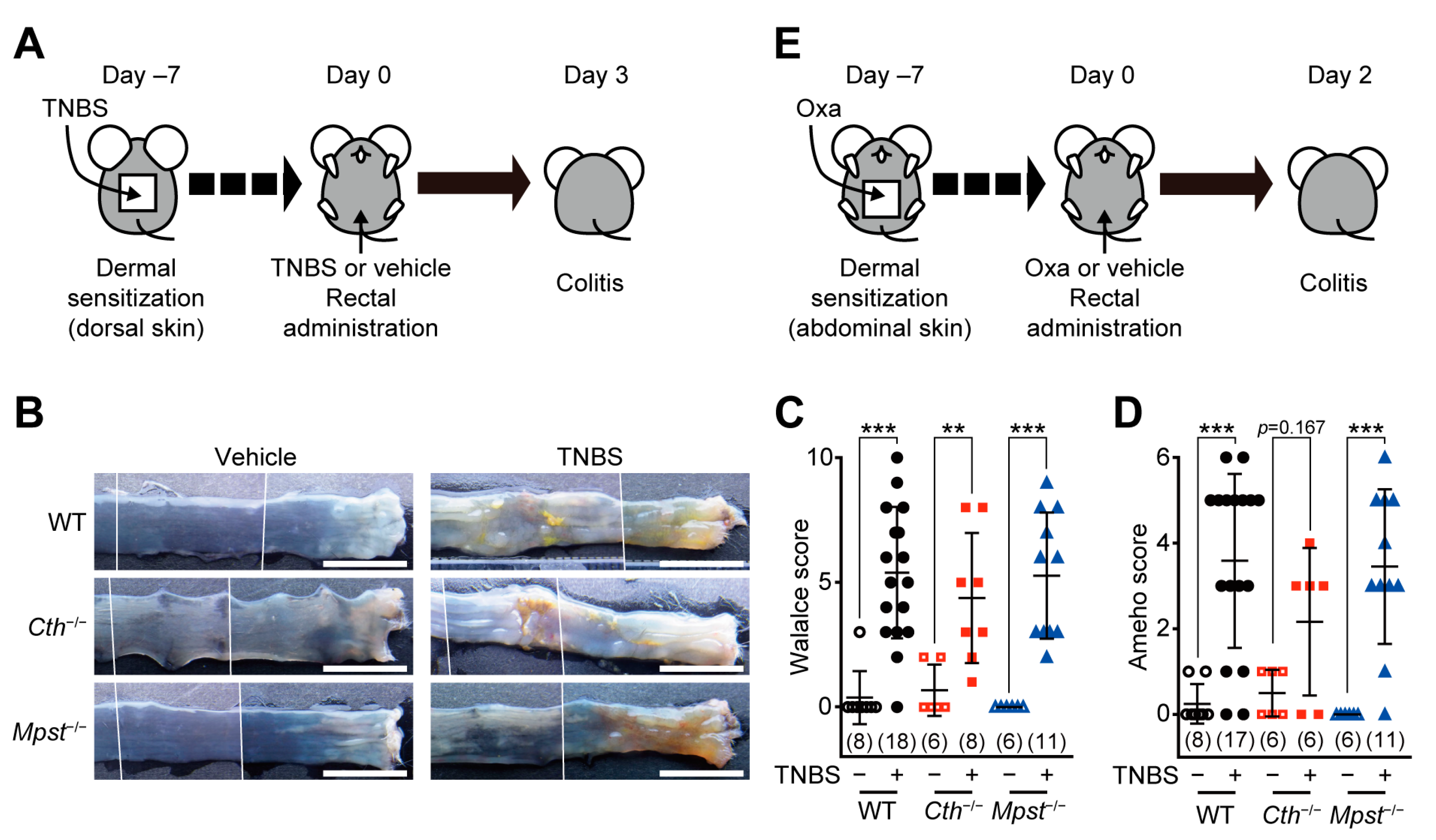
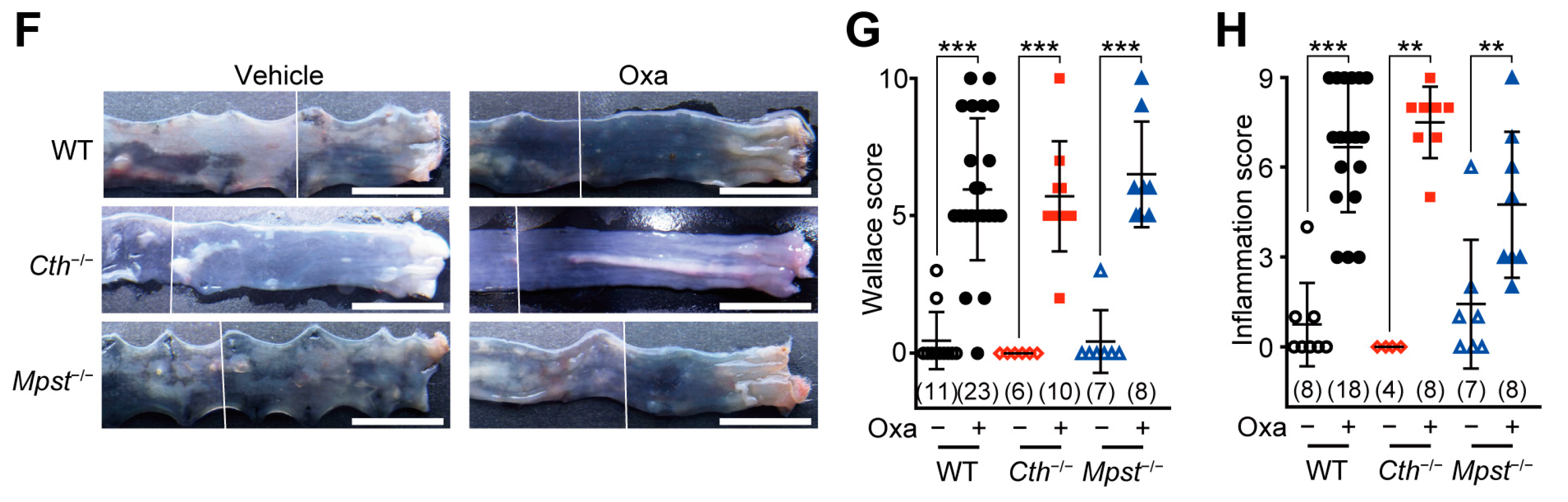
2.1. Normal Responses of Cth–/– and Mpst–/– Mice in TNBS or Oxazolone-Induced Colitis
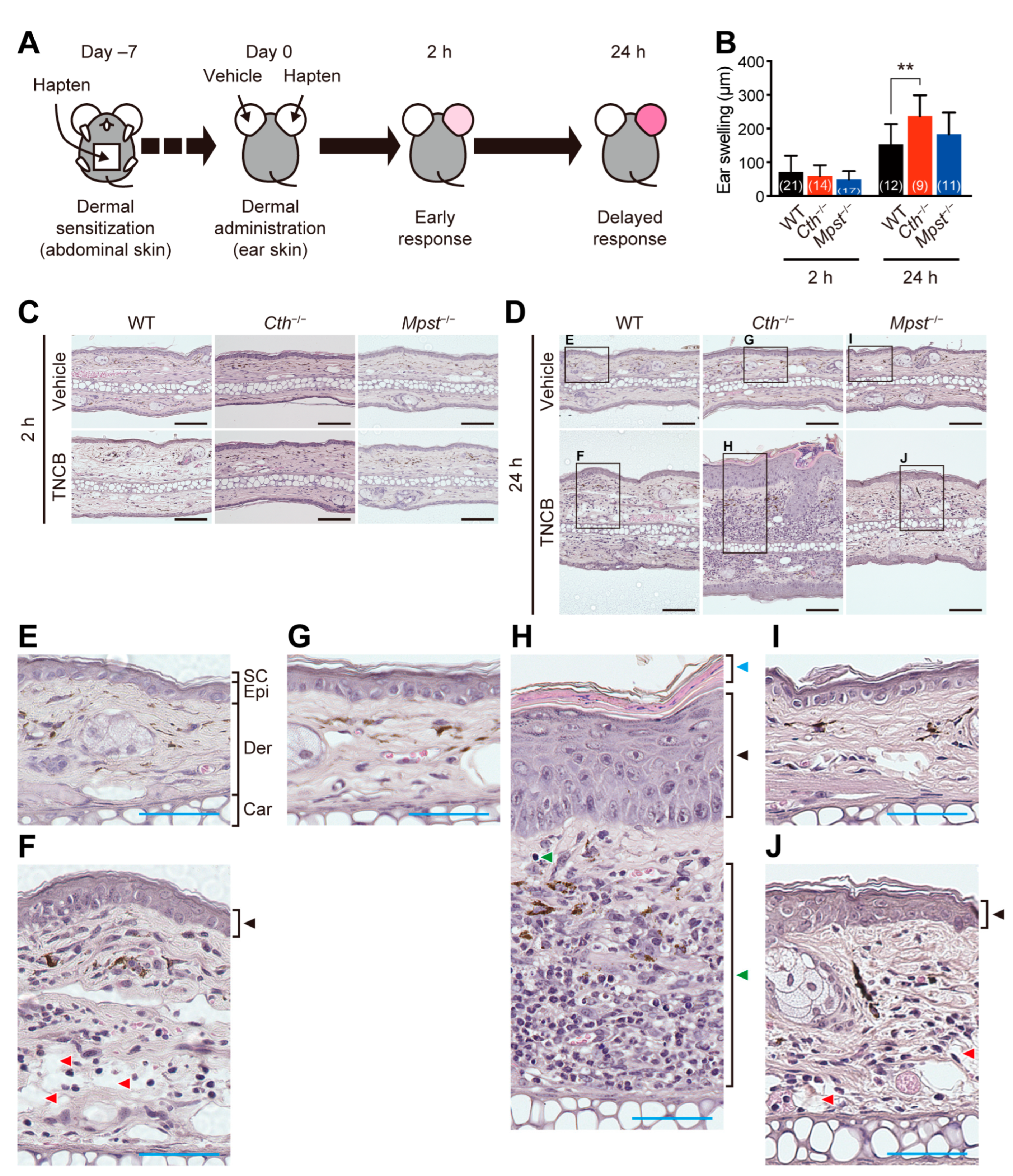
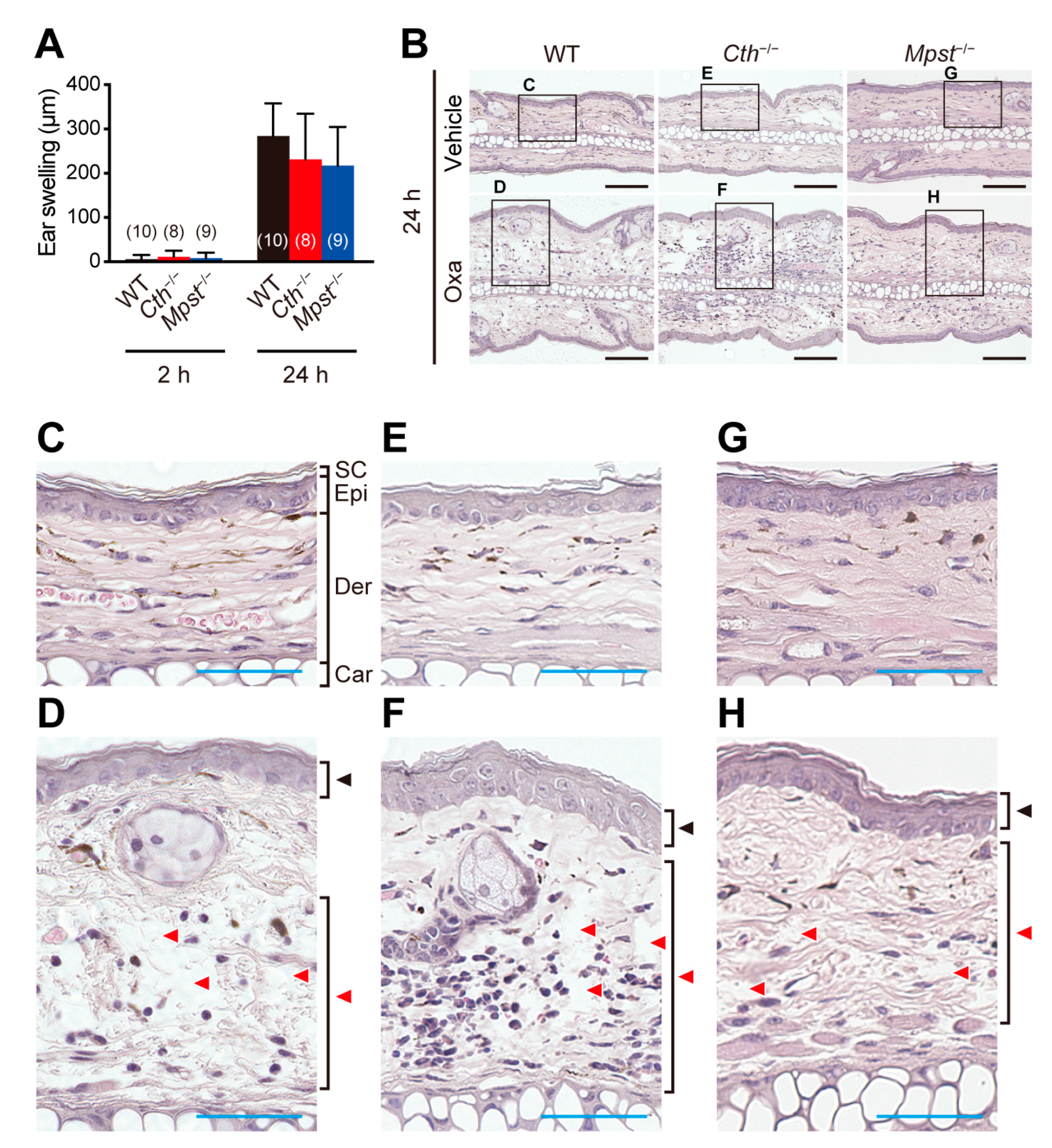
2.2. Exacerbated TNCB (not Oxazolone)-Induced Contact Dermatitis in Cth–/– Mice
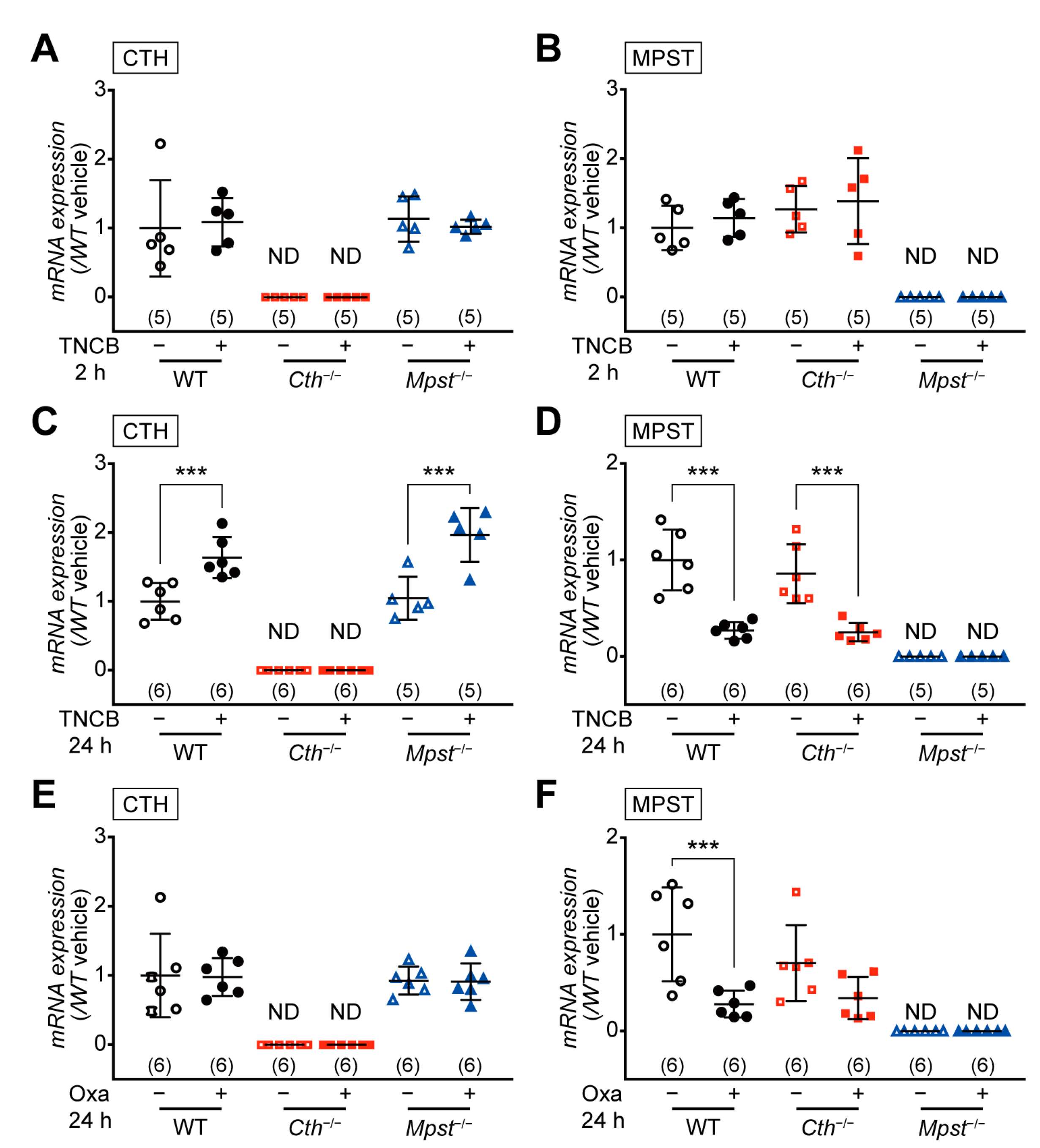
2.3. CTH mRNA Induction in the Delayed Phase of TNCB-Induced Contact Dermatitis
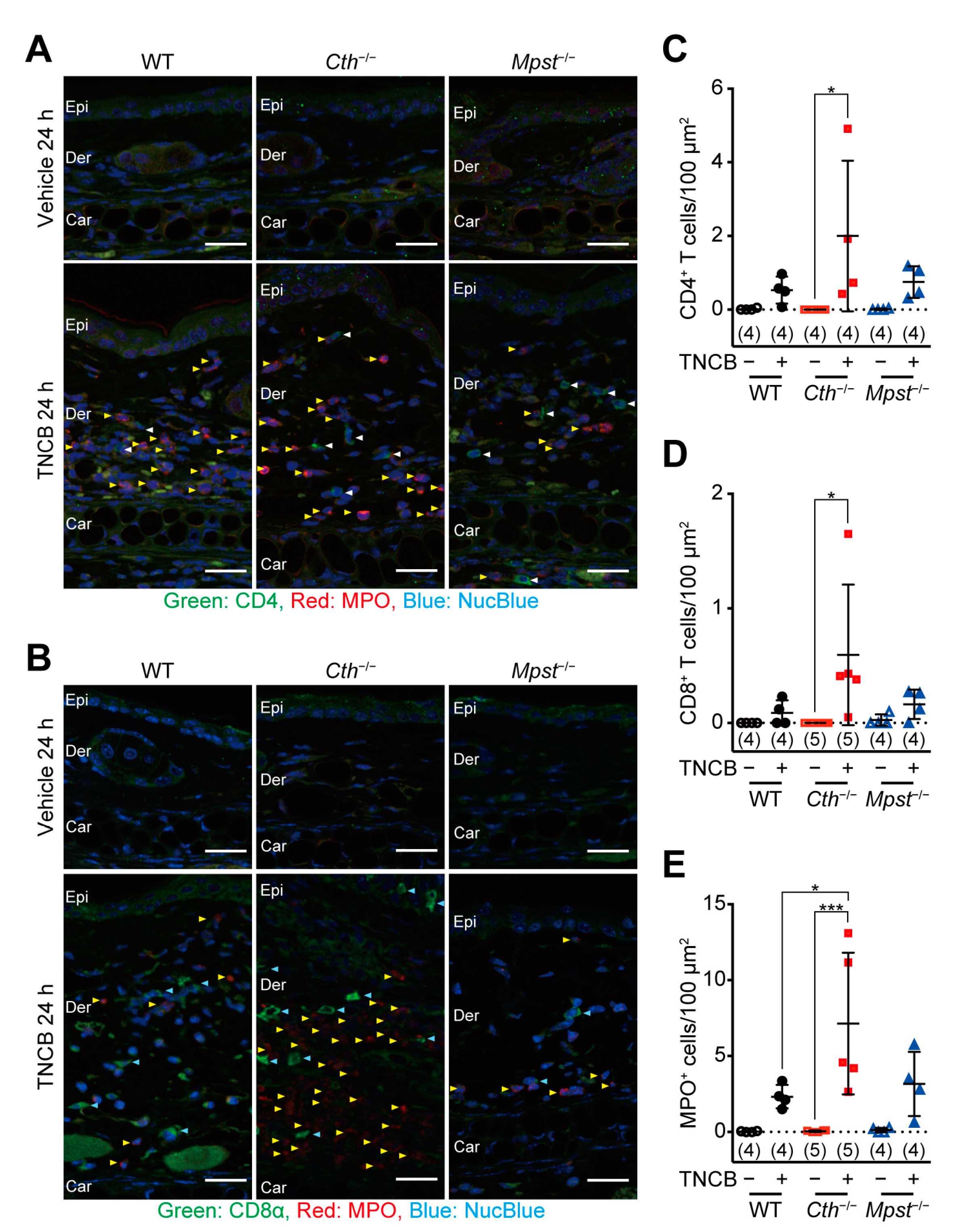
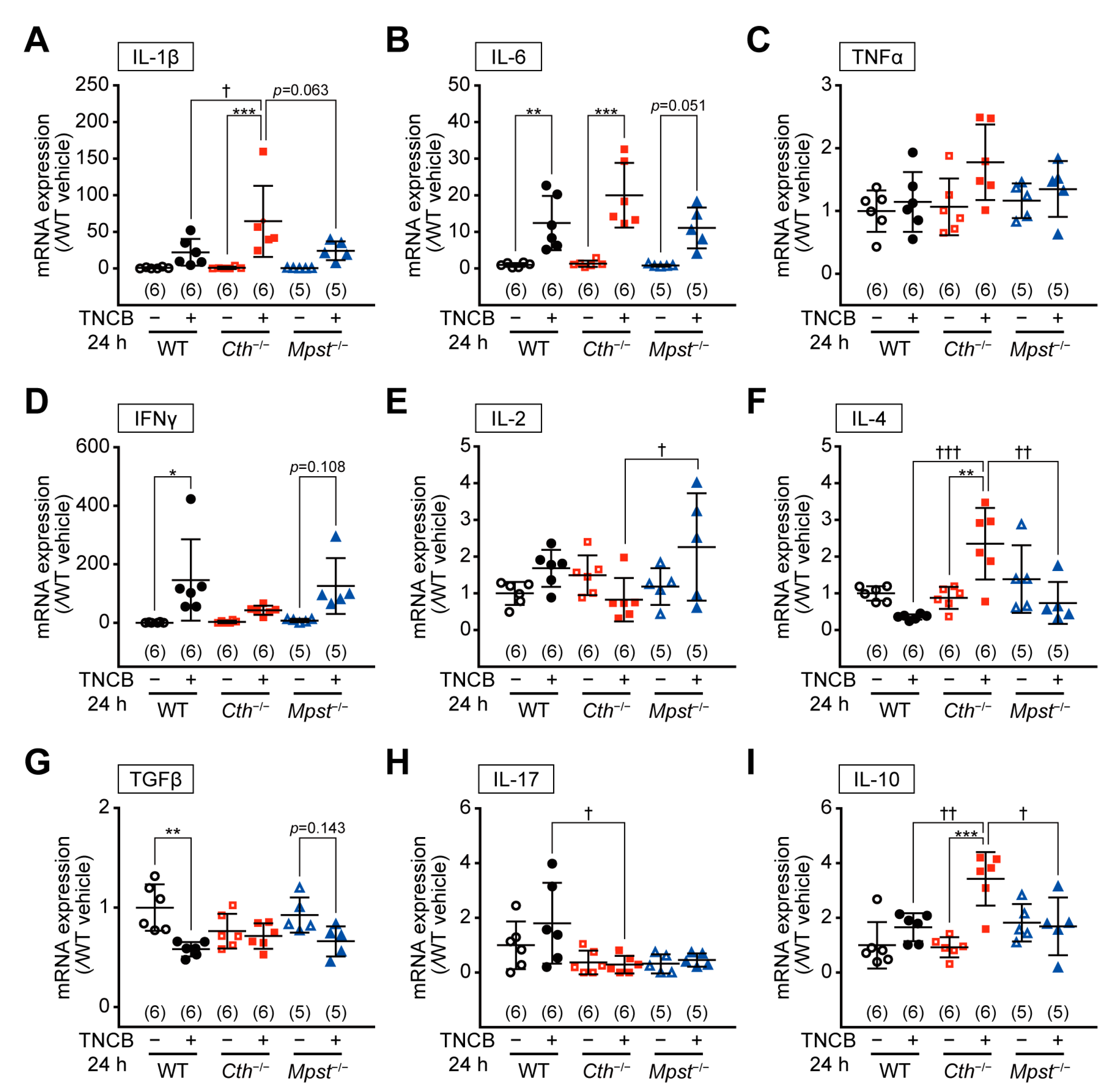
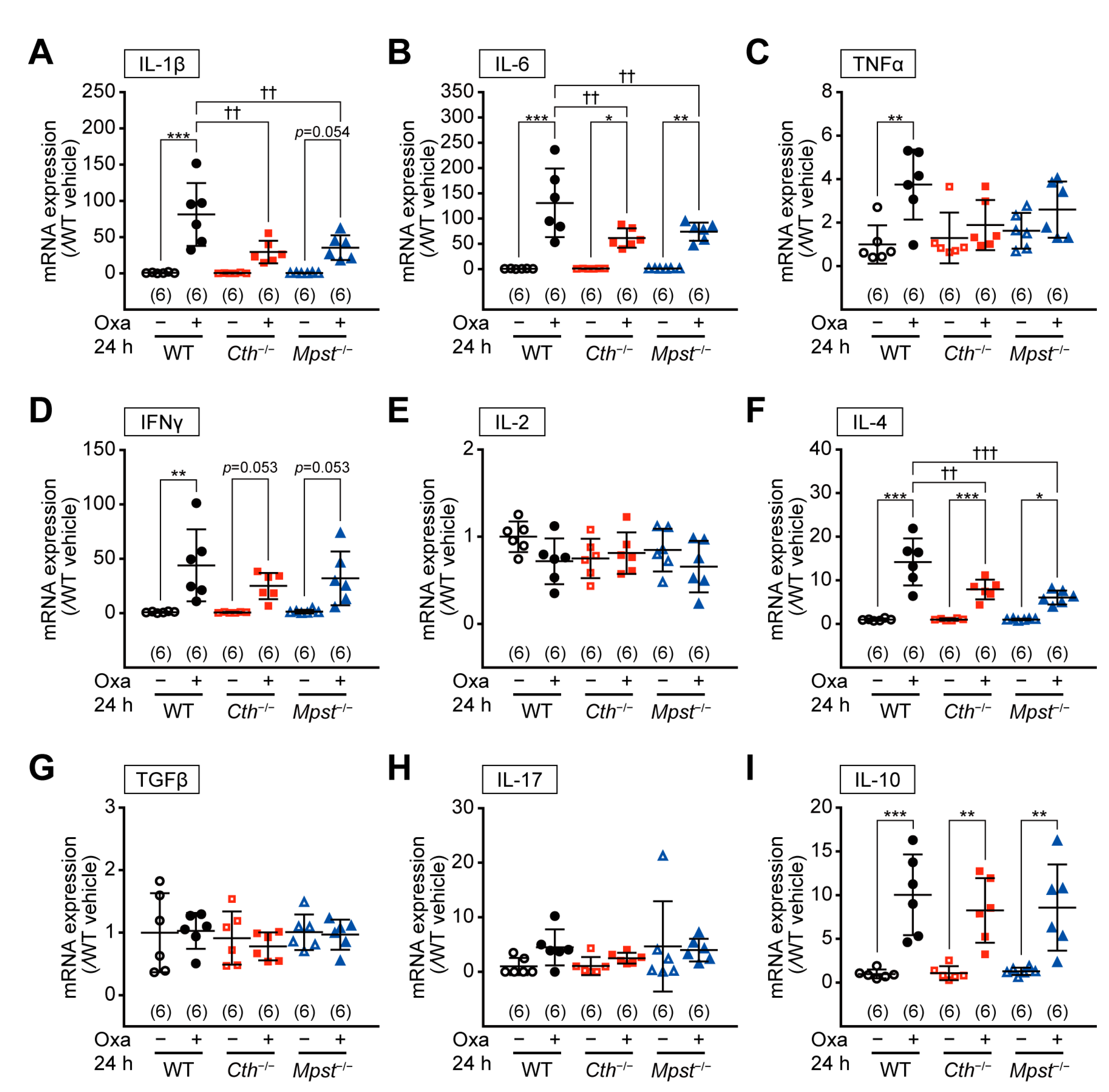
2.4. Marked Induction of Pro-inflammatory/Th2 Cytokine mRNA in the Delayed Phase of TNCB-Induced Contact Dermatitis in Cth–/– Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Hapten-Induced Colitis
4.3. Hapten-Induced Contact Dermatitis
4.4. Immunohistochemistry
4.5. Quantitative RT-PCR
4.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- M’Koma, A.E. Inflammatory Bowel Disease: Clinical Diagnosis and Surgical Treatment-Overview. Medicina (Kaunas) 2022, 58, 567. [Google Scholar] [CrossRef] [PubMed]
- Roda, G.; Chien, N.S.; Kotze, P.G.; Argollo, M.; Panaccione, R.; Spinelli, A.; Kaser, A.; Peyrin-Biroulet, L.; Danese, S. Crohn’s disease. Nat. Rev. Dis. Prim. 2020, 6, 22–40. [Google Scholar] [PubMed]
- Kobayashi, T.; Siegmund, B.; Le Berre, C.; Wei, S.C.; Ferrante, M.; Shen, B.; Bernstein, C.N.; Danese, S.; Peyrin-Biroulet, L.; Hibi, T. Ulcerative colitis. Nat. Rev. Dis. Prim. 2020, 6, 74–93. [Google Scholar] [PubMed]
- Kofla-Dłubacz, A.; Pytrus, T.; Akutko, K.; Sputa-Grzegrzółka, P.; Piotrowska, A.; Dzięgiel, P. Etiology of IBD-Is it still a mystery? Int. J. Mol. Sci. 2022, 23, 12445–12457. [Google Scholar] [PubMed]
- Kakuta, Y.; Naito, T.; Kinouchi, Y.; Masamune, A. Current Status and Future Prospects of Inflammatory Bowel Disease Genetics. Digestion 2023, 104, 7–15. [Google Scholar] [CrossRef] [PubMed]
- Rowan, F.E.; Docherty, N.G.; Coffey, J.C.; O’Connell, P.R. Sulphate-reducing bacteria and hydrogen sulphide in the aetiology of ulcerative colitis. Br. J. Surg. 2009, 96, 151–158. [Google Scholar] [CrossRef] [PubMed]
- Stummer, N.; Weghuber, D.; Feichtinger, R.G.; Huber, S.; Mayr, J.A.; Kofler, B.; Neureiter, D.; Klieser, E.; Hochmann, S.; Lauth, W.; et al. Hydrogen Sulfide Metabolizing Enzymes in the Intestinal Mucosa in Pediatric and Adult Inflammatory Bowel Disease. Antioxid 2022, 11, 2235. [Google Scholar] [CrossRef]
- Murphy, B.; Bhattacharya, R.; Mukherjee, P. Hydrogen sulfide signaling in mitochondria and disease. FASEB J. 2019, 33, 13098–13125. [Google Scholar] [CrossRef]
- Sawa, T.; Motohashi, H.; Ihara, H.; Akaike, T. Enzymatic Regulation and Biological Functions of Reactive Cysteine Persulfides and Polysulfides. Biomolecules 2020, 10, 1245. [Google Scholar] [CrossRef]
- Dilek, N.; Papapetropoulos, A.; Toliver-Kinsky, T.; Szabo, C. Hydrogen sulfide: An endogenous regulator of the immune system. Pharmacol. Res. 2020, 161, 105119–105157. [Google Scholar] [CrossRef]
- Li, M.; Mao, J.; Zhu, Y. New Therapeutic Approaches Using Hydrogen Sulfide Donors in Inflammation and Immune Response. Antioxid. Redox Signal. 2021, 35, 341–356. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, M.; Gaddam, R.R. Hydrogen Sulfide in Inflammation: A Novel Mediator and Therapeutic Target. Antioxid. Redox Signal. 2021, 34, 1368–1377. [Google Scholar] [CrossRef] [PubMed]
- Nagaoka, M.; Radi, Z.A. Pharmacologic efficacy in inflammatory bowel disease models. Front. Biosci (Sch. Ed) 2012, 4, 1295–1314. [Google Scholar] [CrossRef]
- Ishii, I.; Akahoshi, N.; Yamada, H.; Nakano, S.; Izumi, T.; Suematsu, M. Cystathionine gamma-lyase-deficient mice require dietary cysteine to protect against acute lethal myopathy and oxidative injury. J. Biol. Chem. 2010, 285, 26358–26368. [Google Scholar] [CrossRef]
- Akahoshi, N.; Minakawa, T.; Miyashita, M.; Sugiyama, U.; Saito, C.; Takemoto, R.; Honda, A.; Kamichatani, W.; Kamata, S.; Anan, Y.; et al. Increased Urinary 3-Mercaptolactate Excretion and Enhanced Passive Systemic Anaphylaxis in Mice Lacking Mercaptopyruvate Sulfurtransferase, a Model of Mercaptolactate-Cysteine Disulfiduria. Int. J. Mol. Sci. 2020, 21, 818. [Google Scholar] [CrossRef]
- Scheinman, P.L.; Vocanson, M.; Thyssen, J.P.; Johansen, J.D.; Nixon, R.L.; Dear, K.; Botto, N.C.; Morot, J.; Goldminz, A.M. Contact dermatitis. Nat. Rev. Dis. Prim. 2021, 7, 38–63. [Google Scholar] [PubMed]
- Johansen, J.D.; Bonefeld, C.M.; Schwensen, J.F.B.; Thyssen, J.P.; Uter, W. Novel insights into contact dermatitis. J. Allergy Clin. Immunol. 2022, 149, 1162–1171. [Google Scholar] [CrossRef]
- Xiao, Q.; Xiong, L.; Tang, J.; Li, L.; Li, L. Hydrogen Sulfide in Skin Diseases: A Novel Mediator and Therapeutic Target. Oxidative Med. Cell. Longev. 2021, 2021, 6652086–6652096. [Google Scholar] [CrossRef]
- Fernández-Vozmediano, J.M.; Hita, J.C.A.; Manrique-Plaza, A. Allergic contact dermatitis from diallyl disulfide. Contact Dermat. 2000, 42, 108–109. [Google Scholar]
- Shi, Y.; Liu, Z.; Cui, X.; Zhao, Q.; Liu, T. Intestinal vitamin D receptor knockout protects from oxazolone-induced colitis. Cell Death Dis. 2020, 11, 461–471. [Google Scholar] [CrossRef]
- Christensen, A.D.; Haase, C. Immunological mechanisms of contact hypersensitivity in mice. Apmis 2011, 120, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Kimura, H. Hydrogen sulfide (H2S) and polysulfide (H2Sn) signaling: The first 25 years. Biomolecules 2021, 11, 896. [Google Scholar] [CrossRef] [PubMed]
- Di Villa Bianca, R.; Coletta, C.; Mitidieri, E.; De Dominicis, G.; Rossi, A.; Sautebin, L.; Cirino, G.; Bucci, M.; Sorrentino, R. Hydrogen sulphide induces mouse paw oedema through activation of phospholipase A2. Br. J. Pharmacol. 2010, 161, 1835–1842. [Google Scholar] [CrossRef]
- Ang, A.D.; Rivers-Auty, J.; Hegde, A.; Ishii, I.; Bhatia, M. The effect of CSE gene deletion in caerulein-induced acute pancreatitis in the mouse. Am. J. Physiol. Gastrointest Liver Physiol. 2013, 305, G712–G721. [Google Scholar] [CrossRef] [PubMed]
- Gaddam, R.R.; Fraser, R.; Badiei, A.; Chambers, S.; Cogger, V.C.; Le Couteur, D.G.; Ishii, I.; Bhatia, M. Cystathionine-Gamma-Lyase Gene Deletion Protects Mice against Inflammation and Liver Sieve Injury following Polymicrobial Sepsis. PLoS ONE 2016, 11, e0160521–e0160541. [Google Scholar] [CrossRef]
- Markó, L.; Szijártó, I.A.; Filipovic, M.R.; Kaßmann, M.; Balogh, A.; Park, J.-K.; Przybyl, L.; N’Diaye, G.; Krämer, S.; Anders, J.; et al. Role of Cystathionine Gamma-Lyase in Immediate Renal Impairment and Inflammatory Response in Acute Ischemic Kidney Injury. Sci. Rep. 2016, 6, 27517–27526. [Google Scholar] [CrossRef]
- Zhao, H.; Yan, R.; Zhou, X.; Ji, F.; Zhang, B. Hydrogen sulfide improves colonic barrier integrity in DSS-induced inflammation in Caco-2 cells and mice. Int. Immunopharmacol. 2016, 39, 121–127. [Google Scholar] [CrossRef]
- Qin, M.; Long, F.; Wu, W.; Yang, D.; Huang, M.; Xiao, C.; Chen, X.; Liu, X.; Zhu, Y.Z. Hydrogen sulfide protects against DSS-induced colitis by inhibiting NLRP3 inflammasome. Free. Radic. Biol. Med. 2019, 137, 99–109. [Google Scholar] [CrossRef]
- Liu, Y.; Liao, R.; Qiang, Z.; Yang, W.; Cao, J.; Zeng, H. Exogenous H2S Protects Colon Cells in Ulcerative Colitis by Inhibiting NLRP3 and Activating Autophagy. DNA Cell Biol. 2021, 40, 748–756. [Google Scholar] [CrossRef]
- Fiorucci, S.; Orlandi, S.; Mencarelli, A.; Caliendo, G.; Santagada, V.; Distrutti, E.; Santucci, L.; Cirino, G.; Wallace, J.L. Enhanced activity of a hydrogen sulphide-releasing derivative of mesalamine (ATB-429) in a mouse model of colitis. Br. J. Pharmacol. 2007, 150, 996–1002. [Google Scholar] [CrossRef]
- Wallace, J.L.; Vong, L.; McKnight, W.; Dicay, M.; Martin, G.R. Endogenous and Exogenous Hydrogen Sulfide Promotes Resolution of Colitis in Rats. Gastroenterology 2009, 137, 569–578. [Google Scholar] [CrossRef] [PubMed]
- Kupai, K.; Almási, N.; Kósa, M.; Nemcsók, J.; Murlasits, Z.; Török, S.; Al-Awar, A.; Baráth, Z.; Pósa, A.; Varga, C. H2S confers colonoprotection against TNBS-induced colitis by HO-1 upregulation in rats. Inflammopharmacology 2017, 26, 479–489. [Google Scholar] [CrossRef] [PubMed]
- Hirata, I.; Naito, Y.; Takagi, T.; Mizushima, K.; Suzuki, T.; Omatsu, T.; Handa, O.; Ichikawa, H.; Ueda, H.; Yoshikawa, T. Endogenous Hydrogen Sulfide Is an Anti-inflammatory Molecule in Dextran Sodium Sulfate-Induced Colitis in Mice. Dig. Dis. Sci. 2010, 56, 1379–1386. [Google Scholar] [CrossRef]
- Flannigan, K.L.; Ferraz, J.G.P.; Wang, R.; Wallace, J.L. Enhanced Synthesis and Diminished Degradation of Hydrogen Sulfide in Experimental Colitis: A Site-Specific, Pro-Resolution Mechanism. PLoS ONE 2013, 8, e71962–e71970. [Google Scholar] [CrossRef]
- Zhang, J.; Cen, L.; Zhang, X.; Tang, C.; Chen, Y.; Zhang, Y.; Yu, M.; Lu, C.; Li, M.; Li, S.; et al. MPST deficiency promotes intestinal epithelial cell apoptosis and aggravates inflammatory bowel disease via AKT. Redox Biol. 2022, 56, 102469–102484. [Google Scholar] [CrossRef] [PubMed]
- Motta, J.-P.; Flannigan, K.L.; Agbor, T.A.; Beatty, J.K.; Blackler, R.W.; Workentine, M.L.; Da Silva, G.J.; Wang, R.; Buret, A.G.; Wallace, J.L. Hydrogen Sulfide Protects from Colitis and Restores Intestinal Microbiota Biofilm and Mucus Production. Inflamm. Bowel Dis. 2015, 21, 1006–1017. [Google Scholar] [CrossRef]
- Li, M.; Xu, C.; Shi, J.; Ding, J.; Wan, X.; Chen, D.; Gao, J.; Li, C.; Zhang, J.; Lin, Y.; et al. Fatty acids promote fatty liver disease via the dysregulation of 3-mercaptopyruvate sulfurtransferase/hydrogen sulfide pathway. Gut 2018, 67, 2169–2180. [Google Scholar] [CrossRef]
- Alshorafa, A.K.; Guo, Q.; Zeng, F.; Chen, M.; Tan, G.; Tang, Z.; Yin, R. Psoriasis Is Associated with Low Serum Levels of Hydrogen Sulfide, a Potential Anti-inflammatory Molecule. Tohoku J. Exp. Med. 2012, 228, 325–332. [Google Scholar] [CrossRef]
- Coavoy-Sánchez, S.A.; Rodrigues, L.; Teixeira, S.A.; Soares, A.G.; Torregrossa, R.; Wood, M.E.; Whiteman, M.; Costa, S.K.P.; Muscará, M.N. Hydrogen sulfide donors alleviate itch secondary to the activation of type-2 protease activated receptors (PAR-2) in mice. Pharmacol. Res. 2016, 113, 686–694. [Google Scholar] [CrossRef]
- Xu, M.; Hua, Y.; Qi, Y.; Meng, G.; Yang, S. Exogenous hydrogen sulphide supplement accelerates skin wound healing via oxidative stress inhibition and vascular endothelial growth factor enhancement. Exp. Dermatol. 2019, 28, 776–785. [Google Scholar] [CrossRef]
- Yang, C.-T.; Chen, L.; Chen, W.-L.; Li, N.; Chen, M.-J.; Li, X.; Zheng, X.; Zhao, Y.-Z.; Wu, Y.-X.; Xian, M.; et al. Hydrogen sulfide primes diabetic wound to close through inhibition of NETosis. Mol. Cell. Endocrinol. 2019, 480, 74–82. [Google Scholar] [CrossRef]
- Sener, S.; Akbas, A.; Kilinc, F.; Erel, O.; Aktas, A. Thiol/disulfide homeostasis as a marker of oxidative stress in rosacea: A controlled spectrophotometric study. Cutan. Ocul. Toxicol. 2019, 38, 55–58. [Google Scholar] [CrossRef]
- Moniaga, C.S.; Kamata, Y.; Ogawa, H.; Suga, Y.; Tominaga, M.; Takamori, K. Hydrogen sulfide modulates the expression of axon-guidance molecules in human keratinocytes. J. Dermatol. Sci. 2020, 97, 232–235. [Google Scholar] [CrossRef] [PubMed]
- Kimber, I.; Basketter, D.A.; Gerberick, G.F.; Dearman, R.J. Allergic contact dermatitis. Int. Immunopharmacol 2002, 2, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Vocanson, M.; Hennino, A.; Rozières, A.; Poyet, G.; Nicolas, J.F. Effector and regulatory mechanisms in allergic contact dermatitis. Allergy 2009, 64, 1699–1714. [Google Scholar] [CrossRef]
- Honda, T.; Egawa, G.; Grabbe, S.; Kabashima, K. Update of Immune Events in the Murine Contact Hypersensitivity Model: Toward the Understanding of Allergic Contact Dermatitis. J. Investig. Dermatol. 2013, 133, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Du, C.; Jin, M.; Hong, Y.; Li, Q.; Wang, X.H.; Xu, J.M.; Wang, F.; Zhang, Y.; Jia, J.; Liu, C.F.; et al. Downregulation of cystathionine beta-synthase/hydrogen sulfide contributes to rotenone-induced microglia polarization toward M1 type. Biochem. Biophys. Res. Commun 2014, 451, 239–245. [Google Scholar] [CrossRef]
- Yang, R.; Qu, C.; Zhou, Y.; Konkel, J.E.; Shi, S.; Liu, Y.; Chen, C.; Liu, S.; Liu, D.; Chen, Y.; et al. Hydrogen Sulfide Promotes Tet1- and Tet2-Mediated Foxp3 Demethylation to Drive Regulatory T Cell Differentiation and Maintain Immune Homeostasis. Immunity 2015, 43, 251–263. [Google Scholar] [CrossRef]
- Oussalah, A.; Guéant, J.-L.; Peyrin-Biroulet, L. Meta-analysis: Hyperhomocysteinaemia in inflammatory bowel diseases. Aliment. Pharmacol. Ther. 2011, 34, 1173–1184. [Google Scholar] [CrossRef]
- Zhu, S.; Li, J.; Bing, Y.; Yan, W.; Zhu, Y.; Xia, B.; Chen, M. Diet-Induced Hyperhomocysteinaemia Increases Intestinal Inflammation in an Animal Model of Colitis. J. Crohn’s Colitis 2015, 9, 708–719. [Google Scholar] [CrossRef]
- Gao, X.; Li, J.; Chen, M. Effect of Homocysteine on the Differentiation of CD4(+) T Cells into Th17 Cells. Dig. Dis. Sci. 2018, 63, 3339–3347. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Meng, X.; Song, Z. Homocysteine and psoriasis. Biosci. Rep. 2019, 39, BSR20190867–BSR20190881. [Google Scholar] [CrossRef] [PubMed]
- Wallace, J.L.; MacNaughton, W.K.; Morris, G.P.; Beck, P.L. Inhibition of leukotriene synthesis markedly accelerates healing in a rat model of inflammatory bowel disease. Gastroenterology 1989, 96, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Ameho, C.K.; Adjei, A.A.; Harrison, E.K.; Takeshita, K.; Morioka, T.; Arakaki, Y.; Ito, E.; Suzuki, I.; Kulkarni, A.D.; Kawajiri, A.; et al. Prophylactic effect of dietary glutamine supplementation on interleukin 8 and tumour necrosis factor α production in trinitrobenzene sulphonic acid induced colitis. Gut 1997, 41, 487–493. [Google Scholar] [CrossRef]
- Bendtsen, K.M.; Tougaard, P.; Hansen, A.K. An Early Life Mucosal Insult Temporarily Decreases Acute Oxazolone-Induced Inflammation in Mice. Inflammation 2018, 41, 1437–1447. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Akahoshi, N.; Hasegawa, R.; Yamamoto, S.; Takemoto, R.; Yoshizawa, T.; Kamichatani, W.; Ishii, I. Differential Roles of Cystathionine Gamma-Lyase and Mercaptopyruvate Sulfurtransferase in Hapten-Induced Colitis and Contact Dermatitis in Mice. Int. J. Mol. Sci. 2023, 24, 2659. https://doi.org/10.3390/ijms24032659
Akahoshi N, Hasegawa R, Yamamoto S, Takemoto R, Yoshizawa T, Kamichatani W, Ishii I. Differential Roles of Cystathionine Gamma-Lyase and Mercaptopyruvate Sulfurtransferase in Hapten-Induced Colitis and Contact Dermatitis in Mice. International Journal of Molecular Sciences. 2023; 24(3):2659. https://doi.org/10.3390/ijms24032659
Chicago/Turabian StyleAkahoshi, Noriyuki, Ryoka Hasegawa, Shingo Yamamoto, Rintaro Takemoto, Toshiki Yoshizawa, Waka Kamichatani, and Isao Ishii. 2023. "Differential Roles of Cystathionine Gamma-Lyase and Mercaptopyruvate Sulfurtransferase in Hapten-Induced Colitis and Contact Dermatitis in Mice" International Journal of Molecular Sciences 24, no. 3: 2659. https://doi.org/10.3390/ijms24032659
APA StyleAkahoshi, N., Hasegawa, R., Yamamoto, S., Takemoto, R., Yoshizawa, T., Kamichatani, W., & Ishii, I. (2023). Differential Roles of Cystathionine Gamma-Lyase and Mercaptopyruvate Sulfurtransferase in Hapten-Induced Colitis and Contact Dermatitis in Mice. International Journal of Molecular Sciences, 24(3), 2659. https://doi.org/10.3390/ijms24032659