1. Introduction
Because of their high nutritional content and low anti-nutritional components,
Moringa oleifera products have gained prominence and are used for livestock and human diets. Recent studies of the nutrients found in the plant’s leaves, seeds, and stems have revealed that they are high in protein, vital amino acids, minerals, and vitamins such as vitamin C, which prevents scurvy, as well as some B-Complexes and other bioactive chemicals [
1,
2]. Additionally, because they are low in calories, Moringa leaves can be included in an obese person’s diet. Fibrous pods are useful for treating digestive issues and preventing colon cancer [
3,
4]. According to the research, immature pods have a fiber level of about 46.78 percent and a protein content of about 20.66%. The immature pods and flowers showed similar quantities of palmitic, linolenic, linoleic, and oleic acids, whereas the leaves and flowers had higher amino acid contents than the pods (30%, 44%, and 31%). Among the several elements found in Moringa that are necessary for growth and development, calcium is regarded as one of the key minerals for human growth [
5]. The significant variance in the nutritional composition may be caused by elements including the growth environment, harvesting stage, type of soil, and processing technique. Significant levels of vital minerals, vitamins, amino acids, and fatty acids are present in the leaves and seeds [
6,
7].
M. oleifera is significant for its therapeutic effects [
8] as well as its high nutritional content. As cardiac and circulatory stimulants, this plant’s leaves, roots, seeds, bark, fruits, flowers, and immature pods also have antioxidant, diuretic, antihypertensive, cholesterol-lowering, antiulcer, and antispasmodic properties according to [
9,
10]; antipyretic, antiepileptic, anti-inflammatory, antifungal, and antibacterial properties according to [
10,
11]; and antitumor properties according to [
9,
12]. Many in vitro studies have demonstrated the inhibitory activity of the variant extracts from different parts of
M. oleifera on Gram-positive bacteria (
Enterococcus faecalis, methicillin-resistant
Staphylococcus aureus, and
Staphylococcus epidermidis) and Gram-negative bacteria (
Salmonella enterica,
Pseudomonas aeruginosa,
Klebsiella pneumoniae, and
Escherichia coli) isolated from hospital samples [
13,
14]. The antimicrobial activity of the crude extracts on
E. coli and
K. pneumoniae strains have been compared with that of the antibiotic streptomycin [
15,
16].
Understanding the molecular makeup of the target species that will be used as a source of biomolecules should be the first step in the logical planning of a green synthesis approach toward the creation of silver nanoparticles (Ag-NPs) [
17]. In fact, when choosing a suitable biological resource, the balance between the profiles of the molecular components should be one of the most important factors to take into account; otherwise, it will be an entirely empirical trial-and-error method. Unexpectedly, only a small number of research studies have assessed or at least published the bioresource substances used to synthesize Ag-NPs. Additionally, given the current “omics” era, this is an unexpected finding given that there are molecular profiles from multiple potential studies involving creatures from all Earth biomes [
18].
The current research aimed to investigate the chemical composition of Moringa leaves, the polyphenolic burden in variant successive extracts of Moringa leaves, the antimicrobial activity of the ethanolic and aqueous successive extracts, and to study the effect of green chemically synthesized characterized Ag-NPs as antimicrobial agents compared with leaf ethanolic and aqueous successive and crude aqueous extracts.
3. Discussion
The moisture percent was determined to be 7.94 ± 0.10%, DM (dry matter) was 92.06 ± 0.10%, and OM (organic matter) was 88.35 ± 0.54%. The value of organic matter obtained in this study was slightly lower than the value (93.7%) obtained by [
19]. The nitrogen-free extract (sugars and starches) was determined to be 14.05 ± 2.68% (dry weight), and this was much lower than the total carbohydrates (63.11 ± 0.09%) determined by [
19]. The total lipids were determined to be 13.55 ± 1.10%. The total protein value reported by [
20] was lower (27.44%) than the value obtained in this study (28.59 ± 0.11%) (dry weight), but Mutayoba et al. (2011) reported much higher (30.65%) crude protein in Moringa leaves. The ash was determined to be 11.65 ± 0.54%, and the crude fiber was 32.15 ± 1.87%. These findings were much higher than those reported by [
21].
Due to its high nutritional content and beneficial physiological characteristics, Moringa is regarded as a good food source [
22]. Water, protein, sugar, mineral salts, and fatty acids are the main components of MO leaves [
22]. L-arabinose, D-mannose, D-galactose, L-rhamnose, and D-xylose are a few of these sugar compounds that are pharmacologically active and have been demonstrated in numerous large-scale investigations to improve wound healing. Furthermore, numerous studies have shown that natural sugars with diverse antibacterial properties include D-mannose and D-glucose. Additionally, Moringa leaves contain a variety of fatty acids with molecular structures similar to 10-HDA, such as lauric acid, myristic acid, palmitic acid, arachidonic acid, and oleic acid [
23]. In numerous in-depth investigations, the bioactive substance 10-HDA, which is present in royal honey, has been demonstrated to improve wound healing. The transforming growth factor-1 (TGF-1) and vascular endothelial growth factor (VEGF) are two growth factors that these type of fatty acids causes fibroblasts to produce in wounds [
24].
One of the most significant subgroups of the secondary metabolites produced by plants is the phenolic chemicals. In order to scavenge free radicals, phenolics with at least one aromatic ring (C6) and one or more hydroxyl groups are ideal candidates [
25]. Generally, phenolic chemicals act as potential metal chelators and prevent lipid peroxidation by quenching free radicals and producing phenoxy radicals that are stabilized by resonance. The most important family of natural phenolics and flavonoids can directly scavenge re-oxygenic species and has the capacity to provide electrons or hydrogen atoms in a timely manner [
26]. This led to the measurement of the total phenol content (TPC) and total flavonoid content (TFC) of Moringa.
It was noticed that ethyl acetate, aqueous and ethanolic extracts were richer in total phenol content than other extracts, which suggested the presence of water-loving (polar) phenolic compounds. Additionally, total phenol content was relatively higher in hexane extract than in petroleum ether extract, which indicated that the highest amount of present non-polar phenolic compounds was extracted in hexane yield. These results were well-correlated with previous studies [
27]. They reported that the higher recovery of phenolic compounds was in polar solvents’ yields followed by non-polar ones. Adebayo et al. (2018) determined the phenolic and flavonoid contents in Moringa leaves to be varied from 13.61 to 20.42 mg gallic acid equivalence/g sample and 0.58 to 9.81 mg quercetin equivalence/g sample, respectively [
28]. Al-Owaisi et al. (2014) reported that
M. oleifera leaves exhibited the richest phenolic compounds and flavonoid contents among different species of Moringa [
29]. Ethanolic extract of
M. oleifera leaves showed higher phenolic compounds and flavonoid content than that reported by [
30]. In the previous study by Adebayo et al. (2018),
M. oleifera leaves showed higher phenolic content than what was reported by [
31], who used 80% ethanolic extract and obtained 8.21 µg/100 g, while in contrast, the results of this study were less than theirs in the flavonoid content with the same solvent (531.2 mg/g). That study on
M. oleifera leaves was supported by [
32] regarding the ethanolic extract in phenolic and flavonoid content determination and the moderate action of ethyl acetate extract, but also contradicted them for the hexane extract since they did not exhibit any phenolic compounds or flavonoids.
The aim of experimenting with different Moringa successive extracts of different and varying degrees of polarity to determine the phenolic content was to investigate the successive extracts which contain huge amounts of C6 compounds. In particular, the high phenolics and flavonoid content of ethanolic and aqueous successive extracts in Moringa was the critical dependent point of that research. After the determination of TPCs and TFCs in the different successive extracts from Moringa leaves, it was clear that the successive ethanolic and aqueous extracts are the most promising successive extracts to be tested as antimicrobials against some foodborne pathogenic bacteria and some of the other phytopathogens. It was expected that the positive effects of these polar extracts as antimicrobials will be significant against the largest number of microbial strains. Besides investigating Moringa successive extracts to detect which of them contains a huge amount of phenols, then to detect their effects as promising antimicrobials, there was a crucial purpose to reach the commercial and eco-friendly successive extract, then to use it in the creation of silver nanoparticles at the cheapest cost. It was clear that the extract that fits these criteria is the aqueous extract. Therefore, the aqueous extract was the template that was used to synthesize the silver nanoparticles in a newly developed technique.
In order to combat the many types of pathogenic bacteria that are resistant to or less sensitive to the present and traditionally known antibiotics, there is an alarming rise in the number of bacterial strains that are resistant to a variety of antimicrobials [
33,
34]. Scientific research into the antimicrobial properties of many plants has revealed that a wide variety of plant products can impede the growth of dangerous bacteria. Many of these drugs looked to have unique structures and modes of action from those of the antibiotics now in use [
35]. The leaves of
M. oleifera contain numerous phytochemicals with antimicrobial properties, including tannins, phenolic compounds, and flavonoids [
36,
37]. As a result, it would suggest that the antifungal activity observed in these research studies could be attributed to such compounds. Leaf extracts of
M. oleifera have been found to be effective in controlling the growth of fungi such as
Basidiobolus haptosporus and
Basidiobolus ranarums [
38]. Devendra et al. (2011) reported that chloroform (semi-miscible solvent) extract from Moringa leaves showed a growth inhibition against some pathogenic bacteria, such as
Escherichia coli (MTCC 443),
Pseudomonas aeruginosa,
Staphylococcus aureus (MTCC 3160), and
Streptococcus pyogenes (MTCC 442), and also with some of the phytopathogenic fungi, such as
Aspergillus niger (MTCC 1781) and
Candida albicans (MTCC 181), as compared with petroleum ether (completely immiscible “non-polar” solvent) extract, which was not able to inhibit bacterial and fungal strains [
39]. Aqueous and ethanolic extracts of
M. oleifera leaves have shown antibacterial activity by producing zones of inhibition using the disc diffusion method against
Escherichia coli,
Proteus vulgaris, and
Salmonella typhi, respectively, but acetone and chloroform extracts with non-polar natures did not possess any antibacterial activity [
40].
Broad-spectrum activity against all the investigated species of foodborne pathogenic bacteria has been reported for Moringa leaf aqueous extract. According to Patel (2001), Moringa leaves are frequently used in the treatment of bacterial infection, fungal infection, and diarrhea [
41]. This is because Moringa leaves contain chemical compounds such as kaempferol and rutin, which are said to have antibiotic and antioxidant properties that are linked to the inhibition of microorganisms. When compared with
Staph. sciuri,
P. aeruginosa, and
Staph. aureus, the ethanolic extract of Moringa leaves showed higher inhibitory characteristics against
E. coli,
S. enterica,
S. typhi, and
B. cereus. These results were in agreement with a prior study [
42].
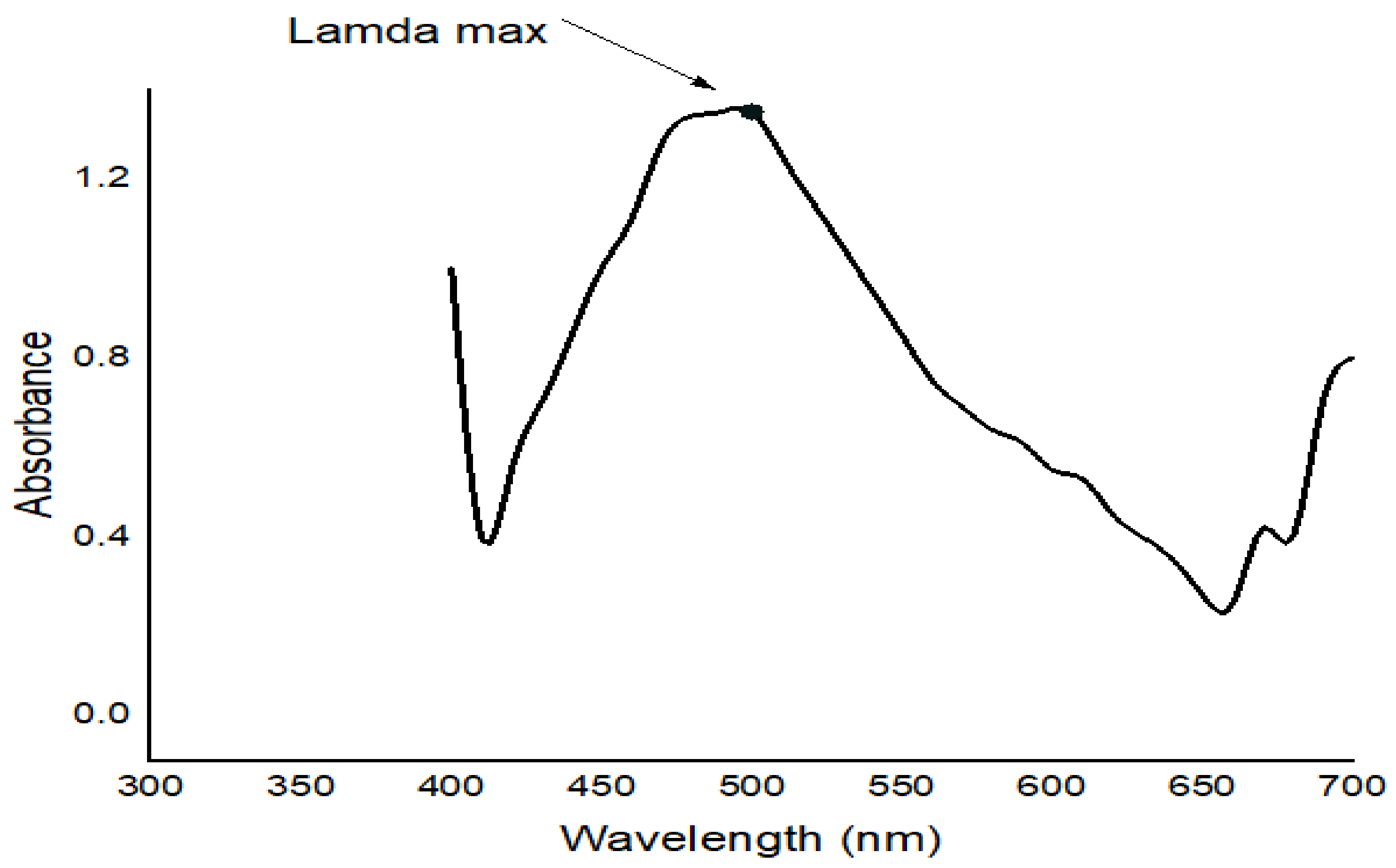
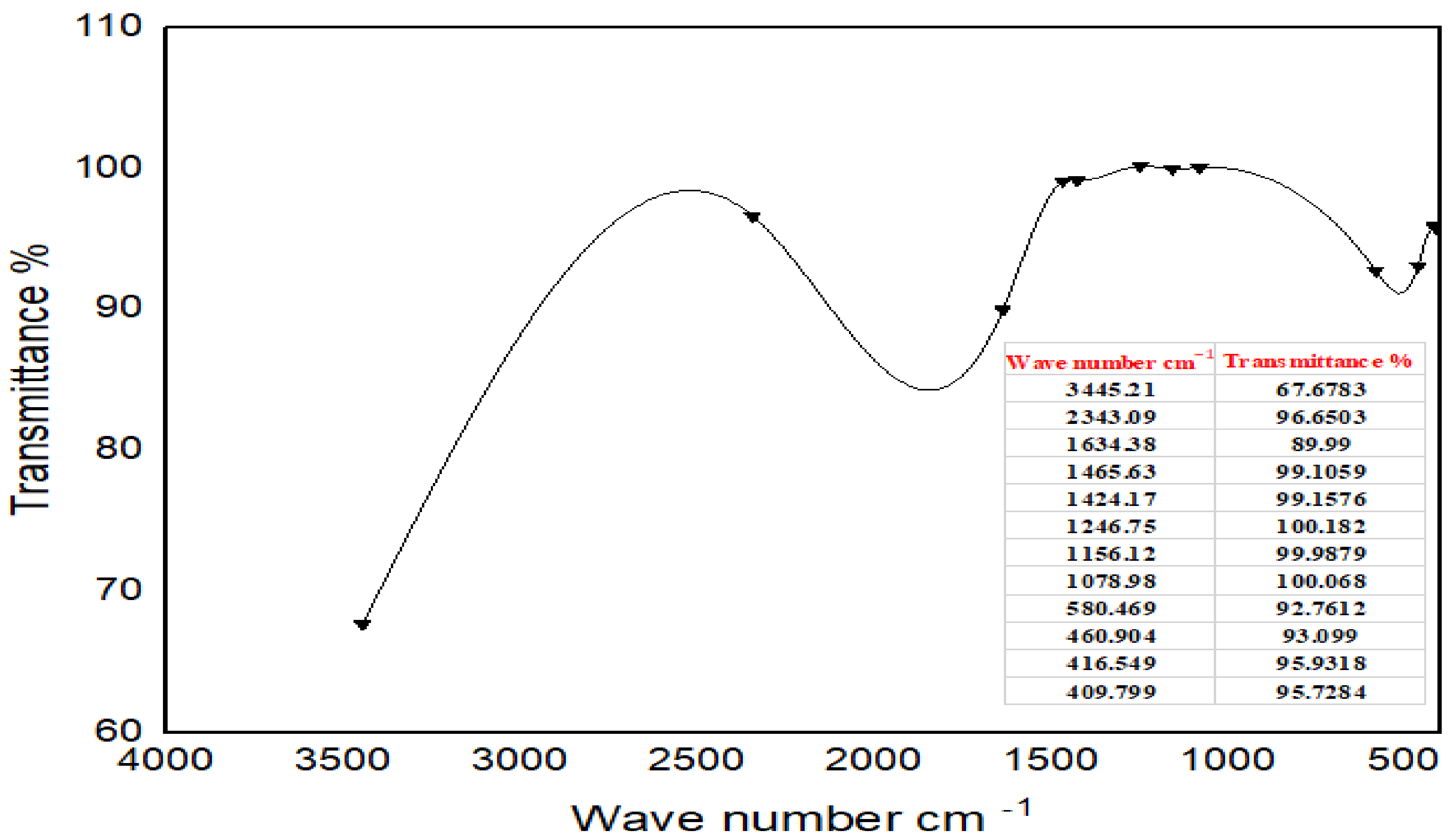
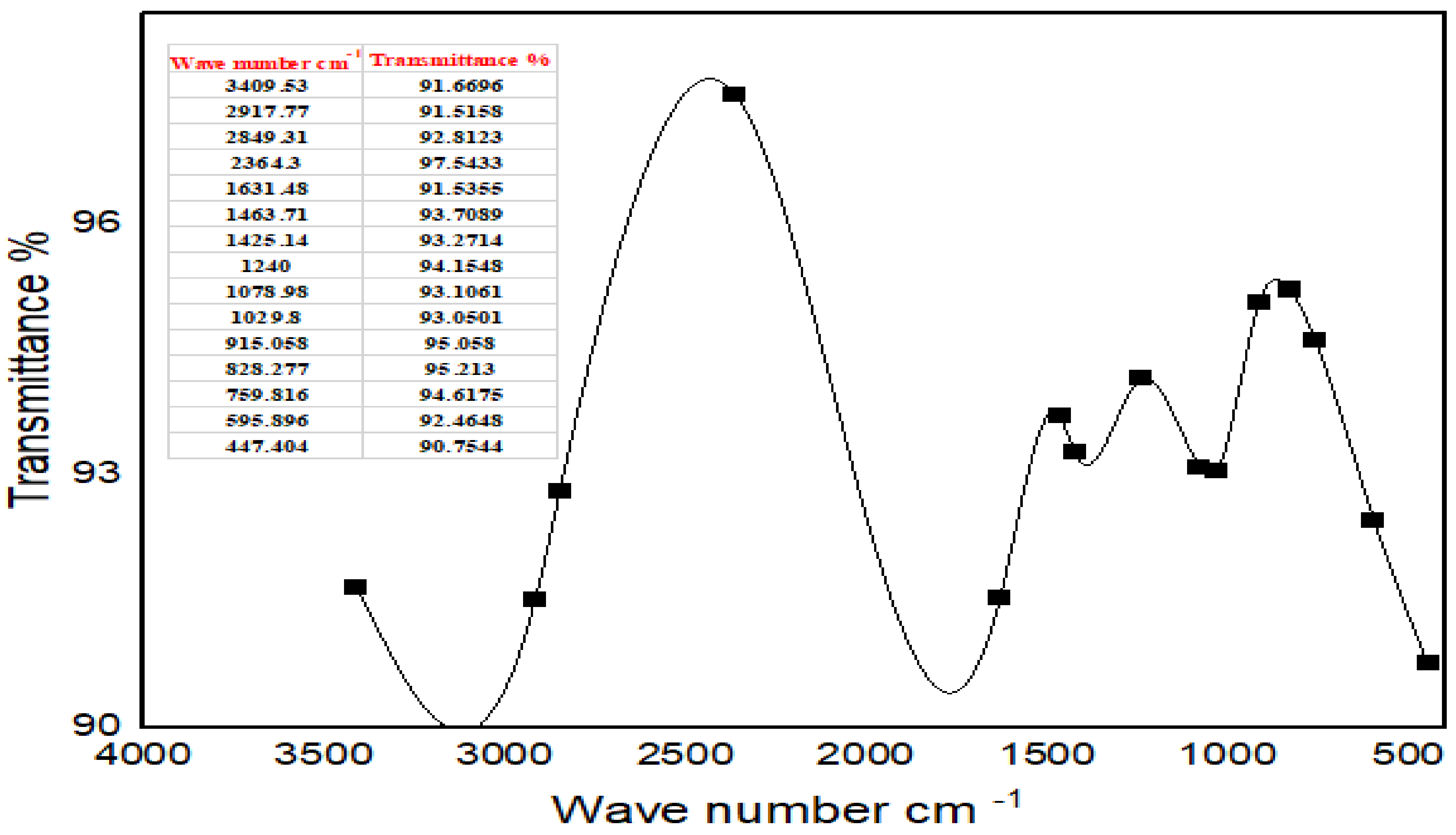
FTIR studies of Moringa aqueous extracts and Ag-NPs synthesized by Moringa crude aqueous extract and NaBH
4 were performed to characterize the chemical structure of the nanoparticles. The FTIR spectrum of Ag-NPs synthesized by Moringa leaves showed the presence of different peaks which were not detected in the FTIR spectra of Moringa crude aqueous extract or which have been shifted. A strong peak at 3409.53 cm
−1 corresponds to the combined peaks of OH group stretching vibration in carboxylic acids. The bands at 2917.77 cm
−1 and 2948.31 cm
−1 in the FTIR spectrum indicated C-H stretching of the alkane amide I band of proteins, and these results were matched with Ganapathy and Sivakumar (2015) [
43]. A band at 2364.3 cm
−1 indicated alkynes. The band at 1631.48 cm
−1 corresponded to amine groups of N–H bending vibrations of proteins and was characteristic of C=O carbonyl groups [
34]. The bands at 1463.71 cm
−1 and 1425.14 cm
−1 indicated nitro group N=O. The peaks at 1240 cm
−1, 1078.98 cm
−1, and 1029.8 cm
−1 corresponded to C-O stretching from alcohol, ether, ester, and carboxylic acid. The peaks at 915.058 cm
−1, 828.277 cm
−1, 828.277 cm
−1, 759.816 cm
−1, 595.896 cm
1, and 474.404 cm
−1 indicated a fingerprint region. They could also indicate C-N imines. The leaves of Moringa may contain high amounts of reductones (enediols with a carbonyl group adjacent to the enediol group), and the highly reducing action of Ag-NPs compared with the crude extracts could be credited to the presence of phenolic functional groups on the surface of synthesized Ag-NPs [
44], which enable them to be more effective as antimicrobials [
45] and antioxidants [
46].
Peixoto et al. (2011) found that aqueous ethanolic extracts of Moringa leaves showed effective antibacterial activity against
Staphylococcus aureus,
Vibrio parahaemolyticus,
Enterococcus faecalis, and
Aeromonas caviae, whereas no effects were seen against
Escherichia coli,
Pseudomonas aeruginosa, and
Salmonella enteritidis [
16]. Out of the 19 Gram-negative bacteria strains tested, the trials by [
47] demonstrated that the methanolic extract of Moringa leaves constrained 13 different bacterial strains [
47]. These strains included
Escherichia coli,
Klebsiella pneumonia, and
Pseudomonas aeruginosa. Moringa leaf extract with antibacterial properties contain alkaloids, polyphenols, flavonoids, anthraquinones, coumarins, tannins, triterpenes, sterols, saponins, and certain secondary metabolites, according to further proximate analysis. It was proposed that Moringa leaf extracts could be used alone or in combination with other medicines to treat a variety of infectious disorders.
These previous results support the results of the current study, as the polar extracts from Moringa usually contain the most effective compounds that are mainly responsible for the inhibition and death of different types of pathogenic bacteria. These bactericidal behaviors may be related to some of the internal compounds such as 4-(α-L-rhamnopyranosyloxy) benzyl isothiocyanate, methyl
N-4-(α-L-rhamnopyranosyloxy) benzyl carbamate, and 4-(β-D-glucopyranosyl-1→4-α-L-rhamnopyranosyloxy)-benzyl thiocarboxamide that were identified from the extract of Moringa [
47].
Along with the different chemical components that have antibacterial activity, Moringa also contains a number of bioactive peptides that have been isolated and studied. As an insecticidal compound against
Aedes ageypti, a lectin from Moringa was first obtained from the seed aqueous extract using chitin column chromatography [
48].
Staphylococcus aureus growth in contaminated waterways was reduced by the water-soluble lectin that was extracted from Moringa, demonstrating the effectiveness of the lectin’s antibacterial properties [
49]. According to Moura et al. (2015), the water-soluble lectin inhibited the growth, survival, and cell permeability of
Bacillus cereus,
Bacillus pumillus,
Bacillus megaterium,
Micrococcus sp.,
Pseudomonas sp.,
Pseudomonas fluorescens,
Pseudomonas stutzeri, and Serratia. Besides, it was noticed that after lectin treatment, some of the bacteria exhibited a tendency for biocorrosion [
50].
Numerous research claimed that Moringa has the potential to control fungal infestation in plants because phytopathogenic fungi can spread disease and endanger the growth of economically important crops. From the Moringa, a thermostable protein known as Moringa chitin-binding protein (Mo-CBP3) was identified using affinity chromatography and ion exchange chromatography. The in vitro antifungal activity showed by Moringa extracts against different fungal strains in the current study may be attributed to Mo-CBP3; as earlier, it showed promising activity against the phytopathogenic fungi
Fusarium solani,
Fusarium oxysporum, and
Colletotrichum gloesporioides at a very low concentration (0.05 mg/mL) with 62% inhibition within 49 h [
51]. Additional research showed that
Fusarium solani’s mycelia development and conidial vitality were inhibited by Mo-CBP3. It was proposed that Mo-CBP3 triggered cell death and reactive oxygen substances (ROS) formation by interacting with the cell membrane in the fungi [
51]. The creation of genetically engineered crops is now considered Mo-CBP3 as a potential antifungal medication because of its physiochemical properties, broad antifungal coverage, and poor toxicity [
52].
Silver nanoparticles were demonstrated to be involved in antibacterial action in addition to phytochemical compounds and bacteriocins (antimicrobial peptides). The extract of Moringa leaves, as demonstrated [
53], produced silver nanoparticles extracellularly by means of the quick reduction of silver ions to form the crystals of nano silvers.
K. pneumoniae,
P. aeruginosa, and
Staph. aureus bacteria’s development was suppressed by solutions of Ag-NPs from Moringa leaf preparations at a concentration of 25 g/mL [
54]. Additionally, they stated that the antifungal effectiveness of the silver nanoparticles made by Moringa was tested using three Candida reference strains. All fungal strains had their growth stopped at a concentration of 6.25 g/mL Ag-NPs, but not with the crude leaf extracts. According to Ankanna et al. (2010), compared with Gram-negative bacteria, Gram-positive bacteria were less resistant to the antibacterial effects of synthesized Ag-NPs from Moringa [
55]. This was explained by the peptidoglycan layer’s negative charge, which prevented Ag
+ ions from freely entering the cell wall and was consistent with the results of this investigation.
Examining the outcomes of antibacterial and antifungal properties for synthesized Ag-NPs and crude aqueous extracts from Moringa, it is believed that the antibacterial activity of crude aqueous extract from Moringa against various Gram-positive and -negative foodborne pathogenic bacteria was caused by affecting the growth and cell permeability of these bacteria. This is because the activity shown by the crude extract may be somewhat related to differences found in the composition of the different types of bacteria. On the other hand, mycelia growth inhibition may be what causes the crude extract’s antifungal efficacy. Some reactive oxygen compounds were thought to originate as a result of interactions with the cell membrane [
51]. Additionally, Moringa Ag-NPs caused the bacterial cells to produce silver ions, which increased their antimicrobial properties. Ag-NP activity may be related to Ag-NP preferential attack on the electron transport chain, cell proliferation, and ultimately cell death.


 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}