
gp120 Envelope Glycoproteins of HIV-1 Group M Subtype A and Subtype B Differentially Affect Gene Expression in Human Vascular Endothelial Cells

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Sequence Analysis of gp120 Subtypes A and B
2.2. gp120 Subtypes A and B Differentially Affect Gene Expression
2.3. gp120 Subtypes A and B Differentially Downregulates Prostasin, MMP-2, and ErbB3
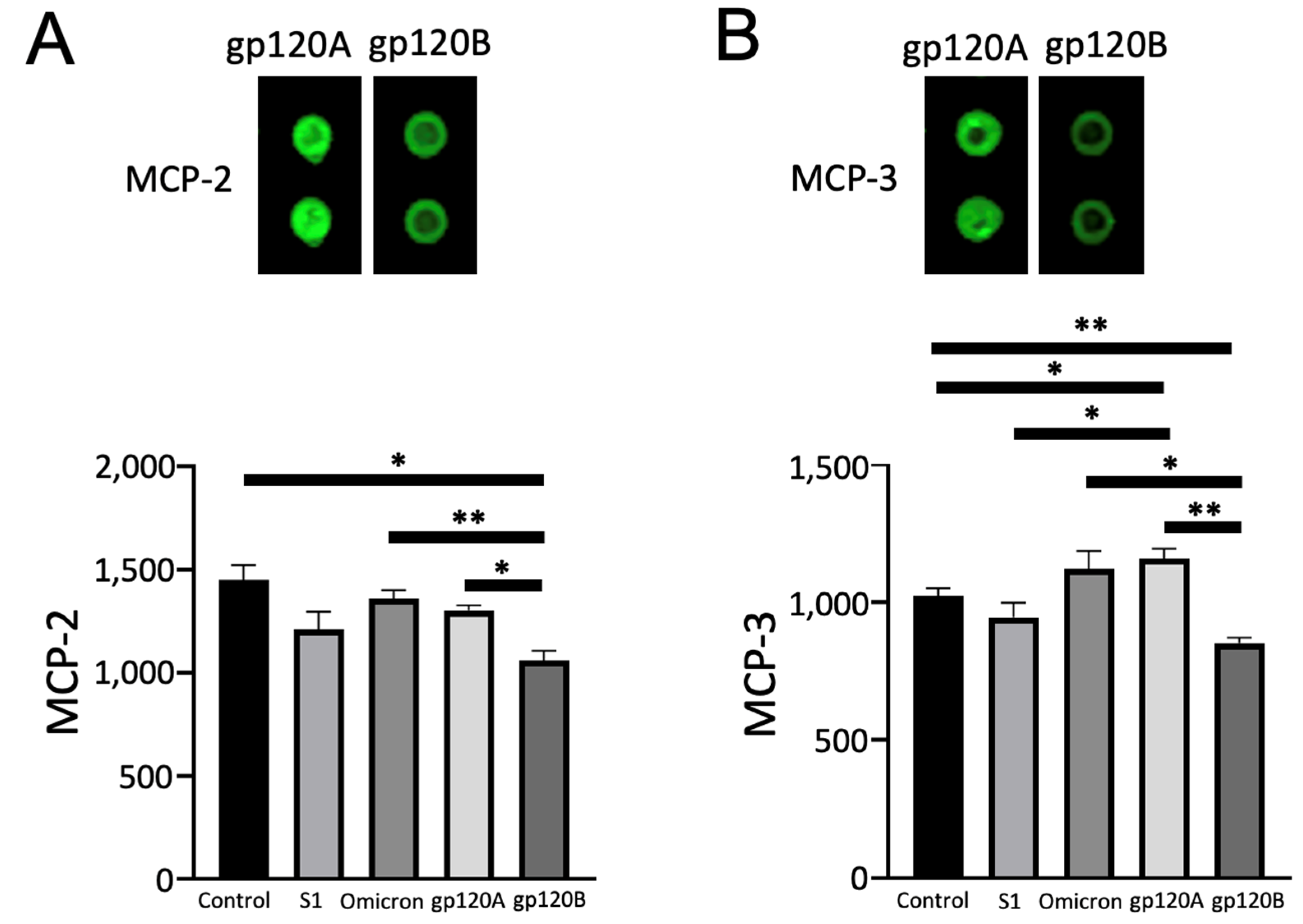
2.4. gp120 Subtypes A and B Differentially Affect MCP-2 and MCP-3
2.5. gp120 Subtypes A and B Differentially Affect TARC
3. Discussion
4. Materials and Methods
4.1. Protein Sequence Analysis
4.2. Cell Culture
4.3. Protein Array
4.4. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Basyal, B.; Jarrett, H.; Barnett, C.F. Pulmonary hypertension in HIV. Can. J. Cardiol. 2019, 35, 288–298. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; He, N. HIV and pulmonary hypertension: CD4 and viral load matter. Lancet Healthy Longev. 2021, 2, e389–e390. [Google Scholar] [CrossRef] [PubMed]
- Parikh, R.V.; Ma, Y.; Scherzer, R.; Heringer, A.S.; Macgregor, J.S.; Martin, J.N.; Deeks, S.G.; Ganz, P.; Hsue, P.Y. Endothelin-1 predicts hemodynamically assessed pulmonary arterial hypertension in HIV infection. PLoS ONE 2016, 11, e0146355. [Google Scholar] [CrossRef] [PubMed]
- Mehta, N.J.; Khan, I.A.; Mehta, R.N.; Sepkowitz, D.A. HIV-Related pulmonary hypertension: Analytic review of 131 cases. Chest 2000, 118, 1133–1141. [Google Scholar] [CrossRef] [PubMed]
- Sitbon, O.; Lascoux-Combe, C.; Delfraissy, J.F.; Yeni, P.G.; Raffi, F.; De Zuttere, D.; Gressin, V.; Clerson, P.; Sereni, D.; Simonneau, G. Prevalence of HIV-related pulmonary arterial hypertension in the current antiretroviral therapy era. Am. J. Respir. Crit. Care Med. 2008, 177, 108–113. [Google Scholar] [CrossRef]
- Isasti, G.; Moreno, T.; Pérez, I.; Cabrera, F.; Palacios, R.; Santos, J. High prevalence of pulmonary arterial hypertension in a cohort of asymptomatic HIV-infected patients. AIDS Res. Hum. Retrovir. 2013, 29, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Pellicelli, A.M.; Barbaro, G.; Palmieri, F.; Girardi, E.; D’Ambrosio, C.; Rianda, A.; Barbarini, G.; Frigiotti, D.; Borgia, M.C.; Petrosillo, N. Primary pulmonary hypertension in HIV patients: A systematic review. Angiology 2001, 52, 31–41. [Google Scholar] [CrossRef]
- Bigna, J.J.R.; Sime, P.S.D.; Koulla-Shiro, S. HIV related pulmonary arterial hypertension: Epidemiology in Africa, physiopathology, and role of antiretroviral treatment. AIDS Res. Ther. 2015, 12, 36. [Google Scholar] [CrossRef]
- Nikolopoulos, G.K.; Kostaki, E.G.; Paraskevis, D. Overview of HIV molecular epidemiology among people who inject drugs in Europe and Asia. Infect. Genet. Evol. 2016, 46, 256–268. [Google Scholar] [CrossRef]
- Saad, M.D.; Shcherbinskaya, A.M.; Nadai, Y.; Kruglov, Y.V.; Antonenko, S.V.; Lyullchuk, M.G.; Kravchenko, O.N.; Earhart, K.C.; Sanchez, J.L.; Birx, D.L.; et al. Molecular epidemiology of HIV Type 1 in Ukraine: Birthplace of an epidemic. AIDS Res. Hum. Retrovir. 2006, 22, 709–714. [Google Scholar] [CrossRef] [Green Version]
- Checkley, M.A.; Luttge, B.G.; Freed, E.O. HIV-1 envelope glycoprotein biosynthesis, trafficking, and incorporation. J. Mol. Biol. 2011, 410, 582–608. [Google Scholar] [CrossRef] [PubMed]
- Hallenberger, S.; Bosch, V.; Angliker, H.; Shaw, E.; Klenk, H.D.; Garten, W. Inhibition of furin-mediated cleavage activation of HIV-1 glycoprotein gp160. Nature 1992, 360, 358–361. [Google Scholar] [CrossRef]
- Schecter, A.D.; Berman, A.B.; Yi, L.; Mosoian, A.; McManus, C.M.; Berman, J.W.; Klotman, M.E.; Taubman, M.B. HIV envelope gp120 activates human arterial smooth muscle cells. Proc. Natl. Acad. Sci. USA 2001, 98, 10142–10147. [Google Scholar] [CrossRef] [PubMed]
- Amsellem, V.; Lipskaia, L.; Abid, S.; Poupel, L.; Houssaini, A.; Quarck, R.; Marcos, E.; Mouraret, N.; Parpaleix, A.; Bobe, R.; et al. CCR5 as a treatment target in pulmonary arterial hypertension. Circulation 2014, 130, 880–891. [Google Scholar] [CrossRef] [PubMed]
- Kanmogne, G.D.; Primeaux, C.; Grammas, P. Induction of apoptosis and endothelin-1 secretion in primary human lung endothelial cells by HIV-1 gp120 proteins. Biochem. Biophys. Res. Commun. 2005, 333, 1107–1115. [Google Scholar] [CrossRef] [PubMed]
- Pasquato, A.; Dettin, M.; Basak, A.; Gambaretto, R.; Tonin, L.; Seidah, N.G.; Di Bello, C. Heparin enhances the furin cleavage of HIV-1 gp160 peptides. FEBS Lett. 2007, 581, 5807–5813. [Google Scholar] [CrossRef]
- Suzuki, Y.J. The viral protein fragment theory of COVID-19 pathogenesis. Med. Hypotheses 2020, 144, 110267. [Google Scholar] [CrossRef]
- Suzuki, Y.J.; Suresh, S.J. SARS-CoV-2 spike protein and lung vascular cells. J. Respir. 2021, 1, 4. [Google Scholar]
- Green, L.A.; Yi, R.; Petrusca, D.; Wang, T.; Elghouche, A.; Gupta, S.K.; Petrache, I.; Clauss, M. HIV envelope protein gp120-induced apoptosis in lung microvascular endothelial cells by concerted upregulation of EMAP II and its receptor, CXCR3. Am. J. Physiol. Lung. Cell Mol. Physiol. 2014, 306, L372–L382. [Google Scholar] [CrossRef]
- Del Cornò, M.; Donninelli, G.; Varano, B.; Da Sacco, L.; Masotti, A.; Gessani, S. HIV-1 gp120 activates the STAT3/interleukin-6 axis in primary human monocyte-derived dendritic cells. J. Virol. 2014, 88, 11045–11055. [Google Scholar] [CrossRef]
- Yang, B.; Akhter, S.; Chaudhuri, A.; Kanmogne, G.D. HIV-1 gp120 induces cytokine expression, leukocyte adhesion, and transmigration across the blood-brain barrier: Modulatory effects of STAT1 signaling. Microvasc. Res. 2009, 77, 212–219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hioe, C.E.; Tuen, M.; Vasiliver-Shamis, G.; Alvarez, Y.; Prins, K.C.; Banerjee, S.; Nádas, A.; Cho, M.W.; Dustin, M.L.; Kachlany, S.C. HIV envelope gp120 activates LFA-1 on CD4 T-lymphocytes and increases cell susceptibility to LFA-1-targeting leukotoxin (LtxA). PLoS ONE 2011, 6, e23202. [Google Scholar] [CrossRef] [PubMed]
- Madeira, F.; Pearce, M.; Tivey, A.R.N.; Basutkar, P.; Lee, J.; Edbali, O.; Madhusoodanan, N.; Kolesnikov, A.; Lopez, R. Search and sequence analysis tools services from EMBL-EBI in 2022. Nucleic Acids Res. 2022, 50, W276–W279. [Google Scholar] [CrossRef] [PubMed]
- Shults, N.V.; Almansour, F.S.; Rybka, V.; Suzuki, D.I.; Suzuki, Y.J. Ligand-mediated dephosphorylation signaling for MAP kinase. Cell. Signal. 2018, 52, 147–154. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein Name: HIV-1 (group M, subtype A, isolate 92RW020) Envelope glycoprotein gp160 Protein (gp120 subunit). |
| Catalog Number: Sino Biological 40403-V08H. |
| Protein Construction: A DNA sequence encoding the HIV-1 envelope glycoprotein gp160 extracellular domain (Glu31-Arg494), termed gp120, was expressed with a polyhistidine tag at the C-terminus. |
| Accession Number: AAT67478.1. |
| Expression Host: HEK293 Cells. |
| Molecular Mass: 465 amino acids and predicts a molecular mass of 52.2 kDa. |
| Formulation: Lyophilized from sterile PBS, pH 7.4. |
| Protein Name: HIV-1 (group M, subtype B, isolate BAL) gp120 Protein. |
| Catalog Number: Sino Biological 40404-V08H. |
| Protein Construction: A DNA sequence encoding the HIV1 (group M, subtype B, isolate BAL) Envelope glycoprotein gp160 Protein (gp120 subunit) (Glu30-Arg509) was expressed with a polyhistidine tag at the C-terminus. |
| Accession Number: AAA44191.1. |
| Expression Host: HEK293 Cells. |
| Molecular Mass: 491 amino acids and predicts a molecular mass of 55.4 kDa. |
| Formulation: Lyophilized from sterile PBS, pH 7.4. |
| Protein Name: SARS-CoV-2 (2019-nCoV) Spike S1-His Recombinant Protein. |
| Catalog Number: Sino Biological 40591-V08H. |
| Protein Construction: A DNA sequence encoding the SARS-CoV-2 (2019-nCoV) spike protein S1 Subunit (Val16-Arg685) was expressed with a polyhistidine tag at the C-terminus. |
| Accession Number: YP_009724390.1. |
| Expression Host: HEK293 Cells. |
| Molecular Mass: 681 amino acids and predicts a molecular mass of 76.5 kDa. |
| Formulation: Lyophilized from sterile PBS, pH 7.4. |
| Protein Name: SARS-CoV-2 B.1.1.529 (Omicron) Spike S1 Protein. |
| Catalog Number: Sino Biological 40591-V08H41. |
| Protein Construction: A DNA sequence encoding the SARS-CoV-2 Spike S1 (with mutations A67V, HV69-70 deletion, T95I, G142D, VYY143-145 deletion, N211 deletion, L212I, ins214EPE, G339D, S371L, S373P, S375F, K417N, N440K, G446S, S477N, T478K, E484A, Q493R, G496S, Q498R, N501Y, Y505H, T547K, D614G, H655Y, N679K, P681H) (Met1-Arg685) was expressed with a polyhistidine tag at the C-terminus. The mutations were identified in the SARS-CoV-2 variant (known as variant B.1.1.529), which emerged in South Africa. |
| Accession Number: YP_009724390.1. |
| Expression Host: HEK293 Cells. |
| Molecular Mass: 678 amino acids and predicts a molecular mass of 76.48 kDa. |
| Formulation: Lyophilized from sterile PBS, pH 7.4. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Suh, A.J.; Suzuki, D.I.; Gychka, S.G.; Brelidze, T.I.; Suzuki, Y.J. gp120 Envelope Glycoproteins of HIV-1 Group M Subtype A and Subtype B Differentially Affect Gene Expression in Human Vascular Endothelial Cells. Int. J. Mol. Sci. 2023, 24, 3536. https://doi.org/10.3390/ijms24043536
Suh AJ, Suzuki DI, Gychka SG, Brelidze TI, Suzuki YJ. gp120 Envelope Glycoproteins of HIV-1 Group M Subtype A and Subtype B Differentially Affect Gene Expression in Human Vascular Endothelial Cells. International Journal of Molecular Sciences. 2023; 24(4):3536. https://doi.org/10.3390/ijms24043536
Chicago/Turabian StyleSuh, Andrew J., Dante I. Suzuki, Sergiy G. Gychka, Tinatin I. Brelidze, and Yuichiro J. Suzuki. 2023. "gp120 Envelope Glycoproteins of HIV-1 Group M Subtype A and Subtype B Differentially Affect Gene Expression in Human Vascular Endothelial Cells" International Journal of Molecular Sciences 24, no. 4: 3536. https://doi.org/10.3390/ijms24043536