
A DNA Finite-State Machine Based on the Programmable Allosteric Strategy of DNAzyme

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Programmable Allosteric Strategy of DNAzyme
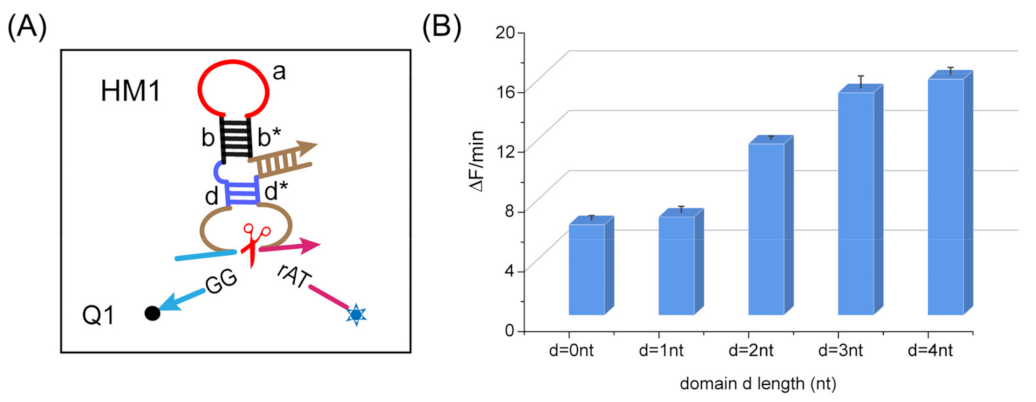
2.2. Parameter Optimization of the Strategy
2.3. Design of the State Machine Reversible Conversion
2.4. Implementation of a DNA Finite-State Machine with Five States
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Assembly Procedure
4.3. Nondenaturing Polyacrylamide Gel Electrophoresis
4.4. Fluorescence Experiments
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Purvis, J.E.; Lahav, G. Encoding and Decoding Cellular Information through Signaling Dynamics. Cell 2013, 152, 945–956. [Google Scholar] [CrossRef]
- Iturbide, A.; Torres-Padilla, M.E. Starting embryonic transcription for the first time. Nat. Genet. 2017, 49, 820–821. [Google Scholar] [CrossRef]
- Park, J.; Lim, J.M.; Jung, I.; Heo, S.J.; Park, J.; Chang, Y.; Kim, H.K.; Jung, D.; Yu, J.H.; Min, S.; et al. Recording of elapsed time and temporal information about biological events using Cas9. Cell 2021, 184, 1047–1063. [Google Scholar] [CrossRef]
- Zuniga, A.; Guiziou, S.; Mayonove, P.; Ben Meriem, Z.; Camacho, M.; Moreau, V.; Ciandrini, L.; Hersen, P.; Bonnet, J. Rational programming of history-dependent logic in cellular populations. Nat. Commun. 2020, 11, 14. [Google Scholar] [CrossRef]
- Bhan, N.; Callisto, A.; Strutz, J.; Glaser, J.; Kalhor, R.; Boyden, E.S.; Church, G.; Kording, K.; Tyo, K.E.J. Recording Temporal Signals with Minutes Resolution Using Enzymatic DNA Synthesis. J. Am. Chem. Soc. 2021, 143, 16630–16640. [Google Scholar] [CrossRef]
- Qin, Z.H.; Liu, Y.; Zhang, L.H.; Liu, J.J.; Su, X. Programming Dissipation Systems by DNA Timer for Temporally Regulating Enzyme Catalysis and Nanostructure Assembly. ACS Nano 2022, 16, 14274–14283. [Google Scholar] [CrossRef] [PubMed]
- Cao, S.T.; Wang, F.; Wang, L.H.; Fan, C.H.; Li, J. DNA nanotechnology-empowered finite state machines. Nanoscale Horiz. 2022, 7, 578–588. [Google Scholar] [CrossRef]
- Hai, X.; Li, Y.F.; Zhu, C.Z.; Song, W.L.; Cao, J.Y.; Bi, S. DNA-based label-free electrochemical biosensors: From principles to applications. TRAC-Trends Anal. Chem. 2020, 133, 20. [Google Scholar] [CrossRef]
- Chen, F.; Lu, Q.J.; Huang, L.N.; Liu, B.W.; Liu, M.L.; Zhang, Y.Y.; Liu, J.W. DNA Triplex and Quadruplex Assembled Nanosensors for Correlating K+ and pH in Lysosomes. Angew. Chem. Int. Ed. 2021, 60, 5453–5458. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.; Burciu, B.; Filipe, C.D.M.; Li, Y.F.; Dellinger, K.; Didar, T.F. DNAzyme-Based Biosensors: Immobilization Strategies, Applications, and Future Prospective. ACS Nano 2021, 15, 13943–13969. [Google Scholar] [CrossRef] [PubMed]
- Abolhasan, R.; Mehdizadeh, A.; Rashidi, M.R.; Aghebati-Maleki, L.; Yousefi, M. Application of hairpin DNA-based biosensors with various signal amplification strategies in clinical diagnosis. Biosens. Bioelectron. 2019, 129, 164–174. [Google Scholar] [CrossRef]
- Du, H.; Yang, P.; Hou, X.; Zhou, R.X.; Hou, X.D.; Chen, J.B. Expanding DNA nanomachine functionality through binding-induced DNA output for application in clinical diagnosis. Chem. Commun. 2019, 55, 3610–3613. [Google Scholar] [CrossRef]
- Zhang, C.; Zhao, Y.M.; Xu, X.M.; Xu, R.; Li, H.W.; Teng, X.Y.; Du, Y.Z.; Miao, Y.Y.; Lin, H.C.; Han, D. Cancer diagnosis with DNA molecular computation. Nat. Nanotechnol. 2020, 15, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Su, H.M.; Xu, J.L.; Wang, Q.; Wang, F.; Zhou, X. High-efficiency and integrable DNA arithmetic and logic system based on strand displacement synthesis. Nat. Commun. 2019, 10, 8. [Google Scholar] [CrossRef]
- Sun, X.B.; Wei, B.; Guo, Y.J.; Xiao, S.Y.; Li, X.; Yao, D.B.; Yin, X.; Liu, S.Y.; Liang, H.J. A Scalable “Junction Substrate” to Engineer Robust DNA Circuits. J. Am. Chem. Soc. 2018, 140, 9979–9985. [Google Scholar] [CrossRef]
- Gong, X.; Wei, J.; Liu, J.; Li, R.M.; Liu, X.Q.; Wang, F. Programmable intracellular DNA biocomputing circuits for reliable cell recognitions. Chem. Sci. 2019, 10, 2989–2997. [Google Scholar] [CrossRef] [PubMed]
- Song, T.Q.; Eshra, A.; Shah, S.L.; Bui, H.; Fu, D.; Yang, M.; Mokhtar, R.; Reif, J. Fast and compact DNA logic circuits based on single-stranded gates using strand-displacing polymerase. Nat. Nanotechnol. 2019, 14, 1075–1081. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.J.; Cao, B.; Liu, Y.; Shi, P.J.; Zheng, Y.F.; Wang, B.; Zhang, Q. TripDesign: A DNA Triplex Design Approach Based on Interaction Forces. J. Phys. Chem. B 2022, 126, 8708–8719. [Google Scholar] [CrossRef]
- Yin, Q.; Zheng, Y.; Wang, B.; Zhang, Q. Design of Constraint Coding Sets for Archive DNA Storage. IEEE/ACM Trans. Comput. Biol. Bioinform. 2021, 19, 3384–3394. [Google Scholar] [CrossRef]
- Zhu, D.; Huang, Z.; Liao, S.; Zhou, C.; Yan, S.; Chen, G. Improved Bare Bones Particle Swarm Optimization for DNA Sequence Design. IEEE Trans. Nanobiosci. 2022. accepted. [Google Scholar] [CrossRef]
- Liu, L.Q.; Hu, Q.Y.; Zhang, W.K.; Li, W.H.; Zhang, W.; Ming, Z.H.; Li, L.J.; Chen, N.; Wang, H.B.; Xiao, X.J. Multifunctional Clip Strand for the Regulation of DNA Strand Displacement and Construction of Complex DNA Nanodevices. Acs Nano 2021, 15, 11573–11584. [Google Scholar] [CrossRef]
- Wang, X.; Sun, H.Z.; Liu, B.J.; Jiang, K.M.; Li, Z.H.; Meng, H.M. DNA Dendrimer-Based Directed 3D Walking Nanomachine for the Sensitive Detection and Intracellular Imaging of miRNA. Anal. Chem. 2022, 94, 17232–17239. [Google Scholar] [CrossRef]
- Wong, R.; Chang, W.L. Fast quantum algorithm for protein structure prediction in hydrophobic-hydrophilic model. J. Parallel Distrib. Comput. 2022, 164, 178–190. [Google Scholar] [CrossRef]
- Chang, W.L.; Chen, J.C.; Chung, W.Y.; Hsiao, C.Y.; Wong, R.; Vasilakos, A.V. Quantum Speedup and Mathematical Solutions of Implementing Bio-Molecular Solutions for the Independent Set Problem on IBM Quantum Computers. IEEE Trans Nanobiosci. 2021, 20, 354–376. [Google Scholar] [CrossRef] [PubMed]
- Roquet, N.; Soleimany, A.P.; Ferris, A.C.; Aaronson, S.; Lu, T.K. Synthetic recombinase-based state machines in living cells. Science 2016, 353, aad8559. [Google Scholar] [CrossRef]
- Santini, C.C.; Bath, J.; Tyrrell, A.M.; Turberfield, A.J. A clocked finite state machine built from DNA. Chem. Commun. 2013, 49, 237–239. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; Hong, F.; Liu, H.; Zhou, X.; Jiang, S.X.; Sulc, P.; Jiang, J.H.; Yan, H. A localized DNA finite-state machine with temporal resolution. Sci. Adv. 2022, 8, abm9530. [Google Scholar] [CrossRef]
- Orbach, R.; Willner, B.; Willner, I. Catalytic nucleic acids (DNAzymes) as functional units for logic gates and computing circuits: From basic principles to practical applications. Chem. Commun. 2015, 51, 4144–4160. [Google Scholar] [CrossRef]
- Cui, M.R.; Li, X.L.; Xu, J.J.; Chen, H.Y. Acid-Switchable DNAzyme Nanodevice for Imaging Multiple Metal Ions in Living Cells. ACS Appl. Mater. Interfaces 2020, 12, 13005–13012. [Google Scholar] [CrossRef]
- Chen, F.; Bai, M.; Cao, K.; Zhao, Y.; Cao, X.W.; Wei, J.; Wu, N.; Li, J.; Wang, L.H.; Fan, C.H.; et al. Programming Enzyme-Initiated Autonomous DNAzyme Nanodevices in Living Cells. ACS Nano 2017, 11, 11908–11914. [Google Scholar] [CrossRef]
- Xu, S.X.; Liu, Y.; Zhou, S.H.; Zhang, Q.; Kasabov, N.K. DNA Matrix Operation Based on the Mechanism of the DNAzyme Binding to Auxiliary Strands to Cleave the Substrate. Biomolecules 2021, 11, 1797. [Google Scholar] [CrossRef]
- Bone, S.M.; Lima, N.E.; Todd, A.V. DNAzyme switches for molecular computation and signal amplification. Biosens. Bioelectron. 2015, 70, 330–337. [Google Scholar] [CrossRef]
- Orbach, R.; Wang, F.; Lioubashevski, O.; Levine, R.D.; Remacle, F.; Willner, I. A full-adder based on reconfigurable DNA-hairpin inputs and DNAzyme computing modules. Chem. Sci. 2014, 5, 3381–3387. [Google Scholar] [CrossRef]
- Orbach, R.; Remacle, F.; Levine, R.D.; Willner, I. DNAzyme-based 2:1 and 4:1 multiplexers and 1:2 demultiplexer. Chem. Sci. 2014, 5, 1074–1081. [Google Scholar] [CrossRef]
- Elbaz, J.; Lioubashevski, O.; Wang, F.A.; Remacle, F.; Levine, R.D.; Willner, I. DNA computing circuits using libraries of DNAzyme subunits. Nat. Nanotechnol. 2010, 5, 417–422. [Google Scholar] [CrossRef]
- Wang, S.; Yue, L.; Willner, I. Enzyme-Guided Selection and Cascaded Emergence of Nanostructured Constitutional Dynamic Networks. Nano Lett. 2020, 20, 5451–5457. [Google Scholar] [CrossRef]
- Zhou, Z.X.; Zhang, P.; Yue, L.; Willner, I. Triggered Interconversion of Dynamic Networks Composed of DNA-Tetrahedra Nanostructures. Nano Lett. 2019, 19, 7540–7547. [Google Scholar] [CrossRef] [PubMed]
- Lilienthal, S.; Luo, G.F.; Wang, S.; Yue, L.; Fischer, A.; Ehrlich, A.; Nahmias, Y.; Willner, I. Constitutional Dynamic Networks-Guided Synthesis of Programmed “Genes”, Transcription of mRNAs, and Translation of Proteins. J. Am. Chem. Soc. 2020, 142, 21460–21468. [Google Scholar] [CrossRef]
- Wang, S.; Yue, L.; Shpilt, Z.; Cecconello, A.; Kahn, J.S.; Lehn, J.M.; Willner, I. Controlling the Catalytic Functions of DNAzymes within Constitutional Dynamic Networks of DNA Nanostructures. J. Am. Chem. Soc. 2017, 139, 9662–9671. [Google Scholar] [CrossRef] [PubMed]
- Yue, L.; Wang, S.; Willner, I. Triggered reversible substitution of adaptive constitutional dynamic networks dictates programmed catalytic functions. Sci. Adv. 2019, 5, aav5564. [Google Scholar] [CrossRef] [Green Version]
- Yue, L.; Wang, S.; Cecconello, A.; Lehn, J.M.; Willner, I. Orthogonal Operation of Constitutional Dynamic Networks Consisting of DNA-Tweezer Machines. ACS Nano 2017, 11, 12027–12036. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Yang, J.; Jiang, S.X.; Liu, Y.; Yan, H. DNAzyme-Based Logic Gate-Mediated DNA Self-Assembly. Nano Lett. 2016, 16, 736–741. [Google Scholar] [CrossRef] [PubMed]
- Dong, J.T.; Ouyang, Y.; Wang, J.B.; O’Hagan, M.P.; Willner, I. Assembly of Dynamic Gated and Cascaded Transient DNAzyme Networks. ACS Nano 2022, 16, 6153–6164. [Google Scholar] [CrossRef]
- Lu, S.S.; Wang, S.; Zhao, J.H.; Sun, J.; Yang, X.R. A pH-regulated stimuli-responsive strategy for RNA-cleaving DNAzyme. Sci. China-Chem. 2020, 63, 404–410. [Google Scholar] [CrossRef]
- Elbaz, J.; Wang, F.A.; Remacle, F.; Willner, I. pH-Programmable DNA Logic Arrays Powered by Modular DNAzyme Libraries. Nano Lett. 2012, 12, 6049–6054. [Google Scholar] [CrossRef]
- Nakama, T.; Takezawa, Y.; Sasaki, D.; Shionoya, M. Allosteric Regulation of DNAzyme Activities through Intrastrand Transformation Induced by Cu(II)-Mediated Artificial Base Pairing. J. Am. Chem. Soc. 2020, 142, 10153–10162. [Google Scholar] [CrossRef] [PubMed]
- Yun, W.; Chen, L.; Yi, Z.; Yi, Y.; Tang, Y.J.; Yang, L.Z. Entropy-driven catalytic reaction-induced hairpin structure switching for fluorometric detection of uranyl ions. Microchim. Acta 2019, 186, 6. [Google Scholar] [CrossRef]
- Takezawa, Y.; Hu, L.Y.; Nakama, T.; Shionoya, M. Sharp Switching of DNAzyme Activity through the Formation of a Cu-II-Mediated Carboxyimidazole Base Pair. Angew. Chem. Int. Ed. 2020, 59, 21488–21492. [Google Scholar] [CrossRef]
- Adornetto, G.; Porchetta, A.; Palleschi, G.; Plaxco, K.W.; Ricci, F. A general approach to the design of allosteric, transcription factor-regulated DNAzymes. Chem. Sci. 2015, 6, 3692–3696. [Google Scholar] [CrossRef]
- Zheng, X.D.; Yang, J.; Zhou, C.J.; Zhang, C.; Zhang, Q.; Wei, X.P. Allosteric DNAzyme-based DNA logic circuit: Operations and dynamic analysis. Nucleic Acids Res. 2019, 47, 1097–1109. [Google Scholar] [CrossRef] [Green Version]
- Cui, X.D.; Liu, Y.; Zhang, Q. DNA tile self-assembly driven by antibody-mediated four-way branch migration. Analyst 2022, 147, 2223–2230. [Google Scholar] [CrossRef] [PubMed]
- Pei, H.; Liang, L.; Yao, G.B.; Li, J.; Huang, Q.; Fan, C.H. Reconfigurable Three-Dimensional DNA Nanostructures for the Construction of Intracellular Logic Sensors. Angew. Chem. Int. Ed. 2012, 51, 9020–9024. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, S.J.; Jiang, B.Y.; Xiang, Y.; Yuan, R. Target-induced structure switching of aptamers facilitates strand displacement for DNAzyme recycling amplification detection of thrombin in human serum. Analyst 2019, 144, 2430–2435. [Google Scholar] [CrossRef] [PubMed]
- Zadeh, J.N.; Steenberg, C.D.; Bois, J.S.; Wolfe, B.R.; Pierce, M.B.; Khan, A.R.; Dirks, R.M.; Pierce, N.A. NUPACK: Analysis and Design of Nucleic Acid Systems. J. Comput. Chem. 2011, 32, 170–173. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Zhang, X.; Shi, P.; Cao, B.; Wang, B. A DNA Finite-State Machine Based on the Programmable Allosteric Strategy of DNAzyme. Int. J. Mol. Sci. 2023, 24, 3588. https://doi.org/10.3390/ijms24043588
Wang J, Zhang X, Shi P, Cao B, Wang B. A DNA Finite-State Machine Based on the Programmable Allosteric Strategy of DNAzyme. International Journal of Molecular Sciences. 2023; 24(4):3588. https://doi.org/10.3390/ijms24043588
Chicago/Turabian StyleWang, Jun, Xiaokang Zhang, Peijun Shi, Ben Cao, and Bin Wang. 2023. "A DNA Finite-State Machine Based on the Programmable Allosteric Strategy of DNAzyme" International Journal of Molecular Sciences 24, no. 4: 3588. https://doi.org/10.3390/ijms24043588