
Delimiting CD34+ Stromal Cells/Telocytes Are Resident Mesenchymal Cells That Participate in Neovessel Formation in Skin Kaposi Sarcoma
 , , and
, , and
Abstract
:1. Introduction
2. Results
2.1. General Characteristics of KS in Patch, Plaque and Nodular Stages
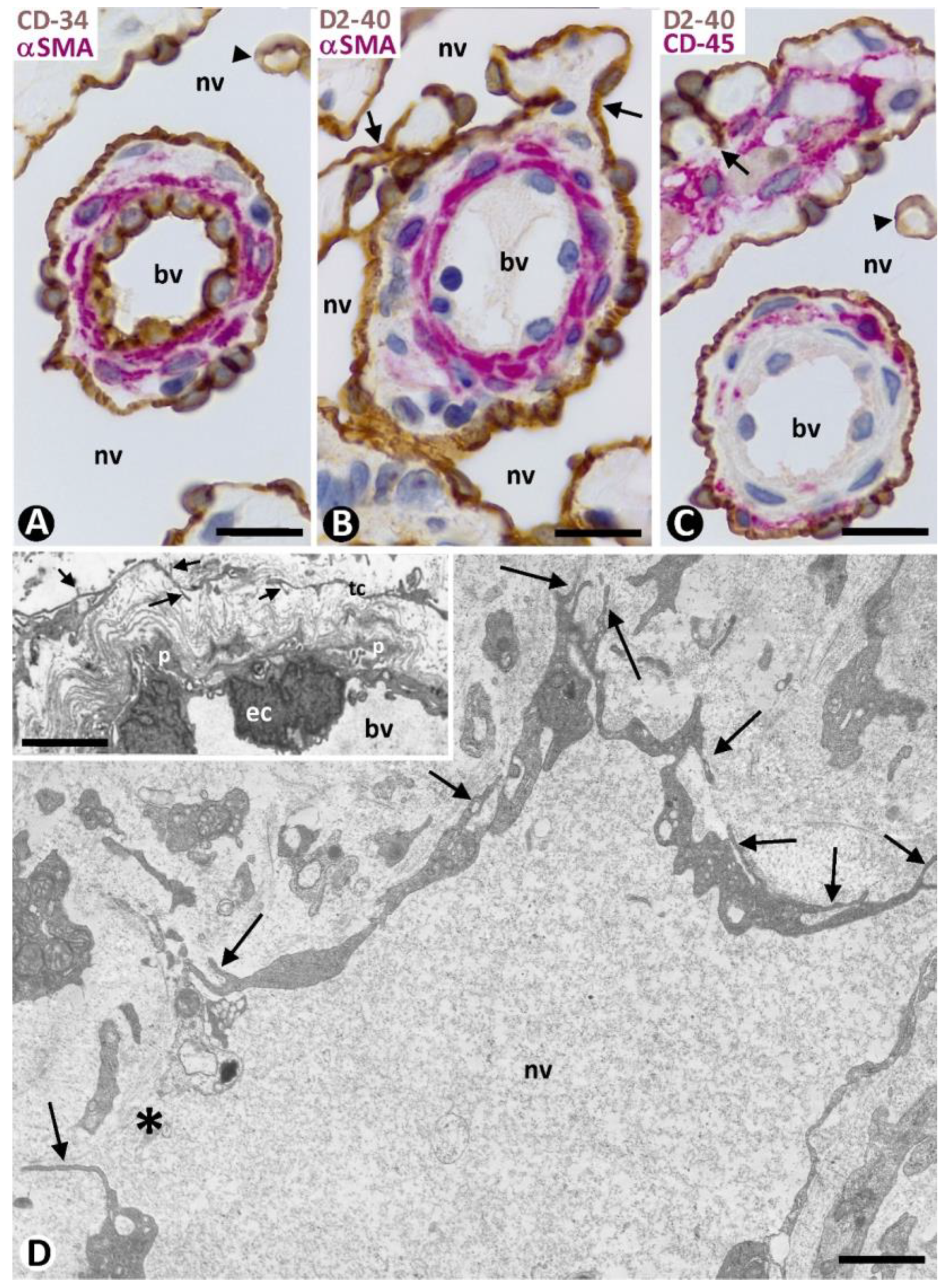
2.2. Pre-Existing (Native) Blood Vessels and Their Relationship with the Inflammatory Infiltrate and Newly Formed Vessels in the Skin Affected by KS in Patch and Plaque Stages
2.2.1. CD34+SCs/TCs in Pre-Existing Blood Vessels without Perivascular Inflammatory Infiltrate or New Vessels (Apparently Non-Affected Blood Vessels)
2.2.2. CD34+SCs/TCs in Pre-Existing Blood Vessels with Perivascular Inflammatory Infiltrate
2.2.3. CD34+SCs/TCs in Pre-Existing Blood Vessels with Perivascular Neovessels
2.3. Initial Neovessels in Early KS Lesions
2.3.1. Characteristics of Type 1 Neovessels
2.3.2. Characteristics of Type 2 Neovessels
2.3.3. Findings Supporting CD34+SC/TC Participation in the Origin of Neovessel ECs
2.3.4. CD34+SCs/TCs during the Formation of Pillars in the Lumen of Neovessels
2.4. Evolution of the Neovessels in KS
2.5. Observations through Serial Histologic Sections
3. Discussion
4. Material and Methods
4.1. Human Tissue Samples
4.2. Light Microscopy
4.3. Immunohistochemistry and Immunofluorescence
4.4. Electron Microscopy
4.5. Quantitative Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaposi, M. Idiopatisches multiples Pigmentsarkom der Haut. Arch. Dermatol. Syph. 1872, 4, 265–273. [Google Scholar] [CrossRef]
- Rabkin, C.S.; Janz, S.; Lash, A.; Coleman, A.E.; Musaba, E.; Liotta, L.; Biggar, R.J.; Zhuang, Z. Monoclonal origin of multicentric Kaposi’s sarcoma lesions. N. Engl. J. Med. 1997, 336, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Duprez, R.; Lacoste, V.; Brière, J.; Couppié, P.; Frances, C.; Sainte-Marie, D.; Kassa-Kelembho, E.; Lando, M.J.; Essame Oyono, J.L.; Nkegoum, B.; et al. Evidence for a multiclonal origin of multicentric advanced lesions of Kaposi sarcoma. J. Natl. Cancer Inst. 2007, 99, 1086–1094. [Google Scholar] [CrossRef]
- Gill, P.S.; Tsai, Y.C.; Rao, A.P.; Spruck, C.H., 3rd; Zheng, T.; Harrington, W.A., Jr.; Cheung, T.; Nathwani, B.; Jones, P.A. Evidence for multiclonality in multicentric Kaposi’s sarcoma. J. Natl. Cancer Inst. 1998, 95, 8257–8261. [Google Scholar] [CrossRef]
- Rusu-Zota, G.; Manole, O.M.; Galeș, C.; Porumb-Andrese, E.; Obadă, O.; Mocanu, C.V. Kaposi Sarcoma, a Trifecta of Pathogenic Mechanisms. Diagnostics 2022, 12, 1242. [Google Scholar] [CrossRef] [PubMed]
- Beckstead, J.H.; Wood, G.S.; Fletcher, V. Evidence for the origin of Kaposi’s sarcoma from lymphatic endothelium. Am. J. Pathol. 1985, 119, 294–300. [Google Scholar]
- Flore, O.; Rafii, S.; Ely, S.; O’Leary, J.J.; Hyjek, E.M.; Cesarman, E. Transformation of primary human endothelial cells by Kaposi’s sarcoma-associated herpesvirus. Nature 1998, 394, 588–592. [Google Scholar] [CrossRef]
- Lee, S.; Kim, J.E.; Johnson, B.A.; Andukuri, A.; Yoon, Y.S. Direct reprogramming into endothelial cells: A new source for vascular regeneration. Regen. Med. 2017, 12, 317–320. [Google Scholar] [CrossRef]
- Mathison, M.; Sanagasetti, D.; Singh, V.P.; Pugazenthi, A.; Pinnamaneni, J.P.; Ryan, C.T.; Yang, J.; Rosengart, T.K. Fibroblast transition to an endothelial “trans” state improves cell reprogramming efficiency. Sci. Rep. 2021, 11, 22605. [Google Scholar] [CrossRef]
- Junker, J.P.; Lönnqvist, S.; Rakar, J.; Karlsson, L.K.; Grenegård, M.; Kratz, G. Differentiation of human dermal fibroblasts towards endothelial cells. Differentiation 2013, 85, 67–77. [Google Scholar] [CrossRef]
- Kurian, L.; Sancho-Martinez, I.; Nivet, E.; Aguirre, A.; Moon, K.; Pendaries, C.; Volle-Challier, C.; Bono, F.; Herbert, J.M.; Pulecio, J.; et al. Conversion of human fibroblasts to angioblast-like progenitor cells. Nat. Methods 2013, 10, 77–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Margariti, A.; Winkler, B.; Karamariti, E.; Zampetaki, A.; Tsai, T.N.; Baban, D.; Ragoussis, J.; Huang, Y.; Han, J.D.; Zeng, L.; et al. Direct reprogramming of fibroblasts into endothelial cells capable of angiogenesis and reendothelialization in tissue-engineered vessels. Proc. Natl. Acad. Sci. USA 2012, 109, 13793–13798. [Google Scholar] [CrossRef] [PubMed]
- Han, J.K.; Chang, S.H.; Cho, H.J.; Choi, S.B.; Ahn, H.S.; Lee, J.; Jeong, H.; Youn, S.W.; Lee, H.J.; Kwon, Y.W.; et al. Direct conversion of adult skin fibroblasts to endothelial cells by defined factors. Circulation 2014, 130, 1168–1178. [Google Scholar] [CrossRef] [PubMed]
- Ding, Y.; Chen, W.; Lu, Z.; Wang, Y.; Yuan, Y. Kaposi’s sarcoma-associated herpesvirus promotes mesenchymal to- endothelial transition by resolving the bivalent chromatin of PROX1 gene. PLoS Pathog. 2021, 17, e1009847. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhong, C.; Liu, D.; Yu, W.; Chen, W.; Wang, Y.; Shi, S.; Yuan, Y. Evidence for Kaposi Sarcoma Originating from Mesenchymal Stem Cell through KSHV-induced Mesenchymal-to-Endothelial Transition. Cancer Res. 2018, 78, 230–245. [Google Scholar] [CrossRef] [PubMed]
- Naipauer, J.; Rosario, S.; Gupta, S.; Premer, C.; Méndez-Solís, O.; Schlesinger, M.; Ponzinibbio, V.; Jain, V.; Gay, L.; Renne, R.; et al. PDGFRA defines the mesenchymal stem cell Kaposi’s sarcoma progenitors by enabling KSHV oncogenesis in an angiogenic environment. PLoS Pathog. 2019, 15, e1008221. [Google Scholar] [CrossRef]
- Gramolelli, S.; Ojala, P.M. Kaposi’s sarcoma herpesvirus-induced endothelial cell reprogramming supports viral persistence and contributes to Kaposi’s sarcoma tumorigenesis. Curr. Opin. Virol. 2017, 26, 156–162. [Google Scholar] [CrossRef]
- Grayson, W.; Pantanowitz, L. Histological variants of cutaneous Kaposi sarcoma. Diagn. Pathol. 2008, 3, 31. [Google Scholar] [CrossRef]
- O’Donnell, P.J.; Pantanowitz, L.; Grayson, W. Unique histologic variants of cutaneous Kaposi sarcoma. Am. J. Dermatopathol. 2010, 32, 244–250. [Google Scholar] [CrossRef]
- Faussone Pellegrini, M.S.; Popescu, L.M. Telocytes. Biomol. Concepts 2011, 2, 481–489. [Google Scholar] [CrossRef]
- Popescu, L.M.; Faussone-Pellegrini, M.S. Telocytes—A case of serendipity: The winding way from Interstitial Cells of Cajal (ICC), via Interstitial Cajal-Like Cells (ICLC) to Telocytes. J. Cell. Mol. Med. 2010, 14, 729–740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; Sáez, F.J.; Díaz-Flores, L., Jr.; Valladares, F.; Madrid, J.F. CD34+ stromal cells/fibroblasts/fibrocytes/telocytes as a tissue reserve and a principal source of mesenchymal cells. Location, morphology, function and role in pathology. Histol. Histopathol. 2014, 29, 831–870. [Google Scholar] [CrossRef] [PubMed]
- Ceafalan, L.; Gherghiceanu, M.; Popescu, L.M.; Simionescu, O. Telocytes in human skin—Are they involved in skin regeneration? J. Cell. Mol. Med. 2012, 16, 1405–1420. [Google Scholar] [CrossRef] [PubMed]
- Cretoiu, D.; Gherghiceanu, M.; Hummel, E.; Zimmermann, H.; Simionescu, O.; Popescu, L.M. FIB-SEM tomography of human skin telocytes and their extracellular vesicles. J. Cell. Mol. Med. 2015, 19, 714–722. [Google Scholar] [CrossRef]
- Manole, C.G.; Simionescu, O. The cutaneous telocytes. Adv. Exp. Med. Biol. 2016, 913, 303–323. [Google Scholar] [CrossRef]
- Manetti, M.; Guiducci, S.; Ruffo, M.; Rosa, I.; Faussone-Pellegrini, M.S.; Matucci-Cerinic, M.; Ibba-Manneschi, L. Evidence for progressive reduction and loss of telocytes in the dermal cellular network of systemic sclerosis. J. Cell. Mol. Med. 2013, 17, 482–496. [Google Scholar] [CrossRef]
- Rusu, M.; Mirancea, N.; Mănoiu, V.; Vâlcu, M.; Nicolescu, M.; Păduraru, D. Skin telocytes. Ann. Anat. 2012, 194, 359–367. [Google Scholar] [CrossRef]
- Wang, L.; Xiao, L.; Zhang, R.; Jin, H.; Shi, H. Ultrastructural and immunohistochemical characteristics of telocytes in human scalp tissue. Sci. Rep. 2020, 10, 1693. [Google Scholar] [CrossRef]
- Arafat, E.A. Ultrastructural and immunohistochemical characteristics of telocytes in the skin and skeletal muscle of newborn rats. Acta Histochem. 2016, 118, 574–580. [Google Scholar] [CrossRef]
- Kang, Y.; Zhu, Z.; Zheng, Y.; Wan, W.; Manole, C.G.; Zhang, Q. Skin telocytes versus fibroblasts: Two distinct dermal cell populations. J. Cell. Mol. Med. 2015, 19, 2530–2539. [Google Scholar] [CrossRef]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; González-Gómez, M.; Rodríguez-Rodriguez, R.; Hernández-León, N.; Díaz- Flores, L., Jr.; Carrasco, J.L. Cd34+ Stromal Cells/Telocytes in Normal and Pathological Skin. Int. J. Mol. Sci. 2021, 22, 7342. [Google Scholar] [CrossRef]
- Romano, E.; Rosa, I.; Fioretto, B.S.; Lucattelli, E.; Innocenti, M.; Ibba-Manneschi, L.; Matucci-Cerinic, M.; Manetti, M. A Two-Step Immunomagnetic Microbead-Based Method for the Isolation of Human Primary Skin Telocytes/CD34+ Stromal Cells. Int. J. Mol. Sci. 2020, 21, 5877. [Google Scholar] [CrossRef] [PubMed]
- Rosa, I.; Faussone-Pellegrini, M.S.; Romano, E.; Ibba-Manneschi, L.; Matucci-Cerinic, M.; Manetti, M. Impairment in the telocyte/CD34+ stromal cell network in human rheumatoid arthritis synovium. Int. J. Mol. Sci. 2021, 25, 2274–2278. [Google Scholar] [CrossRef]
- Braun, J.; Kurtz, A.; Barutcu, N.; Bodo, J.; Thiel, A.; Dong, J. Concerted regulation of CD34 and CD105 accompanies mesenchymal stromal cell derivation from human adventitial stromal cell. Stem Cells Dev. 2013, 22, 815–827. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; González, M.; Sáez, F.J.; Aparicio, F.; Díaz-Flores, L., Jr.; Madrid, J.F. Human resident CD34+ stromal cells/telocytes have progenitor capacity and are a source of αSMA+ cells during repair. Histol. Histopathol. 2015, 30, 615–627. [Google Scholar] [CrossRef]
- Zimmerlin, L.; Donnenberg, V.S.; Rubin, J.P.; Donnenberg, A.D. Mesenchymal markers on human adipose stem/progenitor cells. Cytometry A 2013, 83, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Flores, L.; Gutiérrez, R.; Lizartza, K.; Goméz, M.G.; García, M.P.; Sáez, F.J.; Díaz-Flores, L., Jr.; Madrid, J.F. Behavior of in situ human native adipose tissue CD34+ stromal/progenitor cells during different stages of repair. Tissue- resident CD34+ stromal cells as a source of myofibroblasts. Anat. Rec. 2015, 298, 917–930. [Google Scholar] [CrossRef]
- Cretoiu, D.; Radu, B.M.; Banciu, A.; Banciu, D.D.; Cretoiu, S.M. Telocytes heterogeneity: From cellular morphology to functional evidence. Semin. Cell Dev. Biol. 2017, 64, 26–39. [Google Scholar] [CrossRef]
- Kondo, A.; Kaestner, K.H. Emerging diverse roles of telocytes. Development 2019, 146, 14. [Google Scholar] [CrossRef]
- Bani, D.; Formigli, L.; Gherghiceanu, M.; Faussone-Pellegrini, M.-S. Telocytes as supporting cells for myocardial tissue organization in developing and adult heart. J. Cell. Mol. Med. 2010, 14, 2531–2538. [Google Scholar] [CrossRef]
- Cretoiu, D.; Roatesi, S.; Bica, I.; Plesca, C.; Stefan, A.; Bajenaru, O.; Condrat, C.E.; Cretoiu, S.M. Simulation and modeling of telocytes behavior in signaling and intercellular communication processes. Int. J. Mol. Sci. 2020, 21, 2615. [Google Scholar] [CrossRef] [Green Version]
- Diaz-Flores, L.; Gutiérrez, R.; García, M.P.; González, M.; Madrid, J.F. Telocytes as a source of progenitor cells in regeneration and repair through granulation tissue. Curr. Stem Cell Res. Ther. 2016, 11, 395–403. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; Sáez, F.; Aparicio, F.; Madrid, J.F. Uptake and intracytoplasmic storage of pigmented particles by human CD34+ stromal cells/telocytes: Endocytic property of telocytes. J. Cell. Mol. Med. 2014, 18, 2478–2487. [Google Scholar] [CrossRef] [PubMed]
- Faussone-Pellegrini, M.-S.; Bani, D. Relationships between telocytes and cardiomyocytes during pre- and post-natal life. J. Cell. Mol. Med. 2010, 14, 1061–1063. [Google Scholar] [CrossRef] [PubMed]
- Gherghiceanu, M.; Popescu, L.M. Cardiac telocytes—Their junctions and functional implications. Cell Tissue Res. 2012, 348, 265–279. [Google Scholar] [CrossRef]
- Manetti, M.; Tani, A.; Rosa, I.; Chellini, F.; Squecco, R.; Idrizaj, E.; Zecchi-Orlandini, S.; Ibba-Manneschi, L.; Sassoli, C. Morphological evidence for telocytes as stromal cells supporting satellite cell activation in eccentric contraction-induced skeletal muscle injury. Sci. Rep. 2019, 9, 14515. [Google Scholar] [CrossRef]
- Nicolescu, M.; Bucur, A.; Dinca, O.; Rusu, M.; Popescu, L.M. Telocytes in parotid glands. Anat. Rec. 2012, 295, 378–385. [Google Scholar] [CrossRef]
- Popescu, L.M.; Manole, E.; Şerboiu, C.S.; Manole, C.G.; Suciu, L.C.; Gherghiceanu, M.; Popescu, B.O. Identification of telocytes in skeletal muscle interstitium: Implication for muscle regeneration. J. Cell. Mol. Med. 2011, 15, 1379–1392. [Google Scholar] [CrossRef]
- Vannucchi, M.-G.; Bani, D.; Faussone-Pellegrini, M.-S. Telocytes contribute as cell progenitors and differentiation inductors in tissue regeneration. Curr. Stem Cell Res. Ther. 2016, 11, 383–389. [Google Scholar] [CrossRef]
- Zhao, B.; Chen, S.; Liu, J.; Yuan, Z.; Qi, X.; Qin, J.; Zheng, X.; Shen, X.; Yu, Y.; Qnin, T.J.; et al. Cardiac telocytes were decreased during myocardial infarction and their therapeutic effects for ischaemic heart in rat. J. Cell. Mol. Med. 2012, 17, 123–133. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhang, M.; Qian, M.; Wang, L.; Cismasiu, V.B.; Bai, C.; Popescu, L.M.; Wang, X. Genetic comparison of mouse lung telocytes with mesenchymal stem cells and fibroblasts. J. Cell. Mol. Med. 2013, 17, 567–577. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Wang, Y.; Zhu, P.; Sun, H.; Mou, Y.; Duan, C.; Yao, A.; Lv, S.; Wang, C. Distribution, and characteristics of telocytes as nurse cells in the architectural organization of engineered heart tissues. Sci. China Life Sci. 2014, 57, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Song, D.; Wei, C.; Chen, C.; Yang, Y.; Deng, X.; Gu, J. Telocytes inhibited inflammatory factor expression and enhanced cell migration in LPS-induced skin wound healing models in vitro and in vivo. J. Transl. Med. 2020, 18, 60. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, N.R.J.; Reilly, N.S.; Schrock, D.C.; Hocking, D.C.; Oakes, P.W.; Fowell, D.J. CD4+ T Cell Interstitial Migration Controlled by Fibronectin in the Inflamed Skin. Front. Immunol. 2020, 11, 1501. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Flores, L.; Gutiérrez, R.; Díaz-Flores, L., Jr.; Goméz, M.G.; Sáez, F.J.; Madrid, J.F. Behaviour of telocytes during physiopathological activation. Semin. Cell Dev. Biol. 2016, 55, 50–61. [Google Scholar] [CrossRef]
- Gaylo, A.; Schrock, D.C.; Fernandes, N.R.; Fowell, D.J. T Cell Interstitial Migration: Motility Cues from the Inflamed Tissue for Micro- and Macro-Positioning. Front. Immunol. 2016, 7, 428. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.P.; González-Gómez, M.; Díaz-Flores, L., Jr.; Carrasco, J.L.; Madrid, J.F.; Rodríguez Bello, A. Comparison of the Behavior of Perivascular Cells (Pericytes and CD34+ Stromal Cell/Telocytes) in Sprouting and Intussusceptive Angiogenesis. Int. J. Mol. Sci. 2022, 23, 9010. [Google Scholar] [CrossRef]
- Patan, S.; Tanda, S.; Roberge, S.; Jones, R.C.; Jain, R.K.; Munn, L.L. Vascular morphogenesis and remodeling in a human tumor xenograft: Blood vessel formation and growth after ovariectomy and tumor implantation. Circ. Res. 2001, 89, 732–739. [Google Scholar] [CrossRef]
- Patan, S.; Munn, L.L.; Tanda, S.; Roberge, S.; Jain, R.K.; Jones, R.C. Vascular morphogenesis and remodeling in a model of tissue repair: Blood vessel formation and growth in the ovarian pedicle after ovariectomy. Circ. Res. 2001, 89, 723–731. [Google Scholar] [CrossRef]
- Djonov, V.G.; Kurz, H.; Burri, P.H. Optimality in the developing vascular system: Branching remodeling by means of intussusception as an efficient adaptation mechanism. Dev. Dyn. 2002, 224, 391–402. [Google Scholar] [CrossRef]
- Burri, P.H.; Tarek, M.R. A novel mechanism of capillary growth in the rat pulmonary microcirculation. Anat. Rec. 1990, 228, 35–45. [Google Scholar] [CrossRef] [PubMed]
- Burri, P.H.; Hlushchuk, R.; Djonov, V. Intussusceptive angiogenesis: Its emergence, its characteristics, and its significance. Dev. Dyn. 2004, 231, 474–488. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Flores, L.; Gutiérrez, R.; González-Gómez, M.; García, P.; Sáez, F.J.; Díaz-Flores, L., Jr.; Carrasco, J.L.; Madrid, J.F. Segmentation of Dilated Hemorrhoidal Veins in Hemorrhoidal Disease. Cells Tissues Organs 2018, 205, 120–128. [Google Scholar] [CrossRef]
- Díaz-Flores, L.; Gutiérrez, R.; Pino García, M.; González-Gómez, M.; Díaz-Flores, L., Jr.; Carrasco, J.L. Intussusceptive lymphangiogenesis in the sinuses of developing human foetal lymph nodes. Ann. Anat. 2019, 226, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Paku, S.; Dezso, K.; Bugyik, E.; Tóvári, J.; Tímár, J.; Nagy, P.; Laszlo, V.; Klepetko, W.; Döme, B. A new mechanism for pillar formation during tumor-induced intussusceptive angiogenesis: Inverse sprouting. Am. J. Pathol. 2011, 179, 1573–1585. [Google Scholar] [CrossRef]
- Díaz-Flores, L.; Gutiérrez, R.; González-Gómez, M.; García, M.A.P.; Carrasco, J.L.; Díaz-Flores, L., Jr.; Madrid, J.F.; Álvarez-Argüelles, H. Participation of Intussusceptive Angiogenesis in the Morphogenesis of Lobular Capillary Hemangioma. Sci. Rep. 2020, 10, 4987. [Google Scholar] [CrossRef]
- Masson, P. Hémangiendothéliome Végétant Intravasculaire. Bull. Soc. Anat. 1923, 93, 517–523. [Google Scholar]
- Dabska, M. Malignant endovascular papillary angioendothelioma of the skin in childhood. Clinicopathologic study of 6 cases. Cancer 1969, 24, 503–510. [Google Scholar] [CrossRef]
- Clearkin, K.P.; Enzinger, F.M. Intravascular papillary endothelial hyperplasia. Arch. Pathol. Lab. Med. 1976, 100, 441–444. [Google Scholar]
- Díaz-Flores, L.; Gutiérrez, R.; García-Suárez, M.P.; Sáez, F.J.; Gutiérrez, E.; Valladares, F.; Carrasco, J.L.; Díaz-Flores, L., Jr.; Madrid, J.F. Morphofunctional basis of the different types of angiogenesis and formation of postnatal angiogenesis- related secondary structures. Histol. Histopathol. 2017, 32, 1239–1279. [Google Scholar] [CrossRef]
- Kuo, T.; Sayers, C.P.; Rosai, J. Masson’s “vegetant intravascular hemangioendothelioma:” A lesion often mistaken for angiosarcoma: Study of seventeen cases located in the skin and soft tissues. Cancer 1976, 38, 1227–1236. [Google Scholar] [CrossRef] [PubMed]
- Llamas-Velasco, M.; Angulo, J.; Durán, R.; Fraitag, S.; Kutzner, H.; Requena, L. Hemangioma with dabskoid features: A rare histopathologic variant of acquired hemangioma. J. Cutan. Pathol. 2016, 43, 258–262. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Flores, L.; Gutiérrez, R.; García, M.D.P.; Carrasco, J.L.; Sáez, F.J.; Díaz-Flores, L., Jr.; González-Gómez, M.; Madrid, J.F. Intussusceptive Lymphangiogenesis in Lymphatic Malformations/Lymphangiomas. Anat. Rec. 2019, 302, 2003–2013. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Flores, L.; Gutiérrez, R.; González-Gómez, M.; García, M.P.; Carrasco, J.L.; Díaz-Flores, L., Jr.; Madrid, J.F. Myriad pillars formed by intussusceptive angiogenesis as the basis of intravascular papillary endothelial hyperplasia (IPEH). IPEH is intussusceptive angiogenesis made a lesion. Histol. Histopathol. 2021, 36, 217–228. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Flores, A.; Rodriguez, R. Promontory sign in a reactive benign vascular proliferation. Am. J. Dermatopathol. 2010, 32, 700–703. [Google Scholar] [CrossRef]
- Lazova, R.; McNiff, J.M.; Glusac, E.J.; Godic, A. Promontory sign—Present in patch and plaque stage of angiosarcoma. Am. J. Dermatopathol. 2009, 31, 132–136. [Google Scholar] [CrossRef]
- Chen, X.; Zeng, J.; Huang, Y.; Gong, M.; Ye, Y.; Zhao, H.; Chen, Z.; Zhang, H. Telocytes and their structural relationships with surrounding cell types in the skin of silky fowl by immunohistochemistrical, transmission electron microscopical and morphometric analysis. Poult. Sci. 2021, 100, 101367. [Google Scholar] [CrossRef]
- Ravalli, S.; Federico, C.; Lauretta, G.; Saccone, S.; Pricoco, E.; Roggio, F.; Di Rosa, M.; Maugeri, G.; Musumeci, G. Morphological Evidence of Telocytes in Skeletal Muscle Interstitium of Exercised and Sedentary Rodents. Biomedicines 2021, 9, 807. [Google Scholar] [CrossRef]

















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Differential Findings | Common Findings | ||||
|---|---|---|---|---|---|
| Endothelial Cell Characteristics | Vessel Lumen | Percentages | and Relationship with Pre-Existing Blood Vessels | ||
| Early Stages | Advanced Stages | ||||
| Type 1 neovessel | Flattened | Irregular |
| ||
| Discontinuities | Lymphangiomatous aspect in advanced stages | 56.77% | 13.70% | ||
| Processes surrounding perivascular collagen. | Frequent folds, pillars and promontory sign | ||||
| Type 2 neovessel | Fusiform (spindle cell-type) | Elongated, virtual or small | 43.23% | 86.93% | |
| Formation of rows (Fascicular aspect) | Frequent intraluminal red blood cells in advanced stages | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Díaz-Flores, L.; Gutiérrez, R.; González-Gómez, M.; García, M.d.P.; Palmas, M.; Carrasco, J.L.; Madrid, J.F.; Díaz-Flores, L., Jr. Delimiting CD34+ Stromal Cells/Telocytes Are Resident Mesenchymal Cells That Participate in Neovessel Formation in Skin Kaposi Sarcoma. Int. J. Mol. Sci. 2023, 24, 3793. https://doi.org/10.3390/ijms24043793
Díaz-Flores L, Gutiérrez R, González-Gómez M, García MdP, Palmas M, Carrasco JL, Madrid JF, Díaz-Flores L Jr. Delimiting CD34+ Stromal Cells/Telocytes Are Resident Mesenchymal Cells That Participate in Neovessel Formation in Skin Kaposi Sarcoma. International Journal of Molecular Sciences. 2023; 24(4):3793. https://doi.org/10.3390/ijms24043793
Chicago/Turabian StyleDíaz-Flores, Lucio, Ricardo Gutiérrez, Miriam González-Gómez, Maria del Pino García, Marta Palmas, Jose Luis Carrasco, Juan Francisco Madrid, and Lucio Díaz-Flores, Jr. 2023. "Delimiting CD34+ Stromal Cells/Telocytes Are Resident Mesenchymal Cells That Participate in Neovessel Formation in Skin Kaposi Sarcoma" International Journal of Molecular Sciences 24, no. 4: 3793. https://doi.org/10.3390/ijms24043793