
Dysfunction of Prkcaa Links Social Behavior Defects with Disturbed Circadian Rhythm in Zebrafish
 ,
,
Abstract
:1. Introduction
2. Results
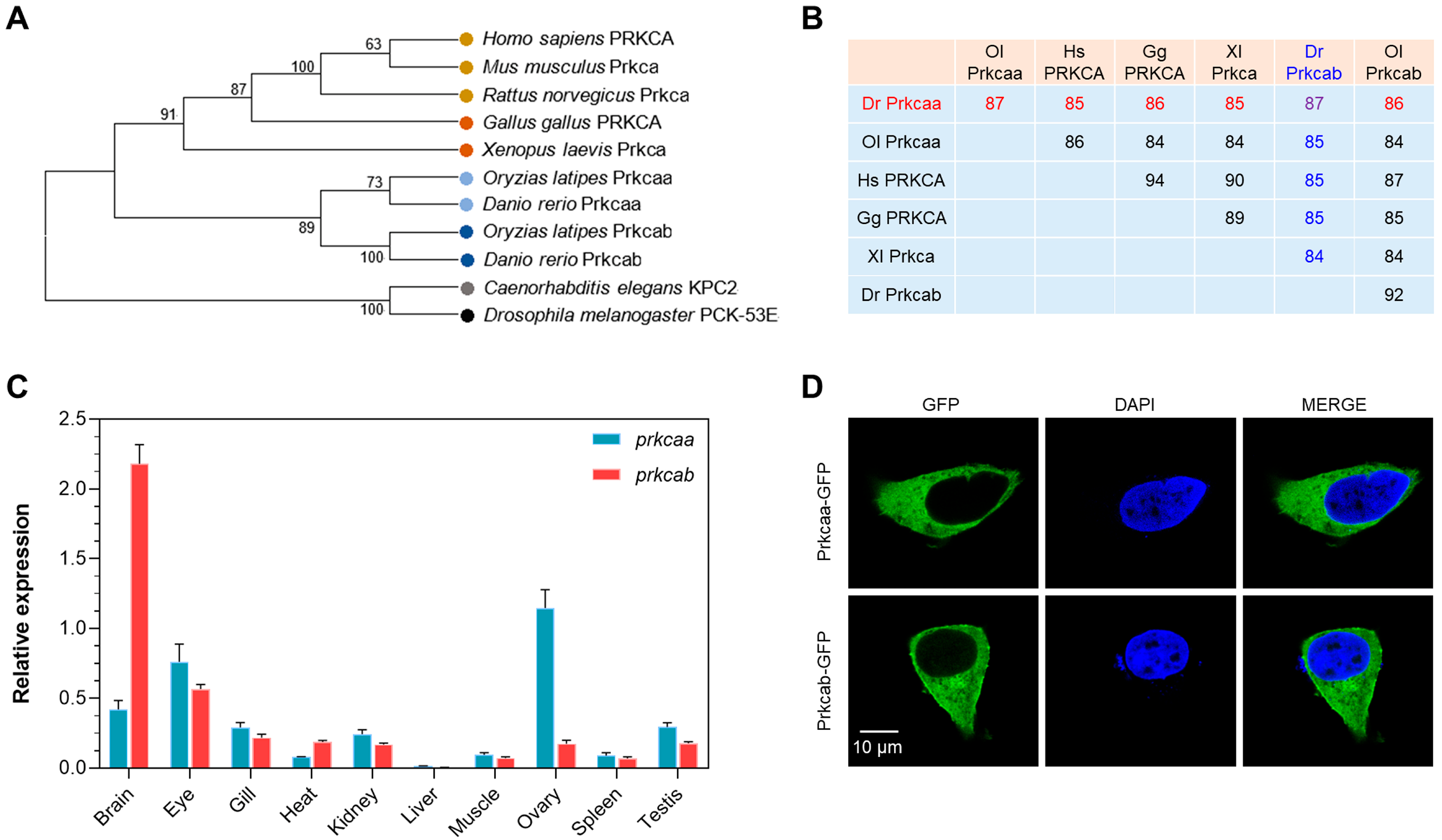
2.1. Molecular Characterization of Zebrafish Prkcaa and Prkcab
2.2. Generation of the prkcaa-Knockout Zebrafish Lines
2.3. Dysfunction of Prkcaa Led to Behavioral Defects in Zebrafish
2.3.1. The prkcaa−/− Mutants Exhibited Anxiety-Like Behavior
2.3.2. The prkcaa−/− Mutants Displayed Impaired Aggressive Behavior
2.3.3. The prkcaa−/− Mutants Demonstrated Decreased Social Preference
2.3.4. The prkcaa−/− Mutants Had Defects in Shoaling Behavior
2.4. Effects of prkcaa Mutation on Gene Expression
2.5. Functional Enrichments for the Differentially Expressed Genes
2.6. The Mutation of prkcaa Affected Genes Involved in Neural Activities
2.7. The 3145A Mutants Had Inverse Rhythm of Locomotion Activity
3. Discussion
4. Materials and Methods
4.1. Zebrafish Maintenance and Measurement
4.2. Phylogenetic Analysis
4.3. Total RNA Extraction
4.4. Real-Time Quantitative PCR Assay
4.5. Molecular Cloning
4.6. Cell Culture, Transfection and Microphotography
4.7. Generation of Gene-Knockout Zebrafish Lines
4.8. Behavioral Assays
4.8.1. Novel Tank Test
4.8.2. Mirror Biting Test
4.8.3. Social Preference Test
4.8.4. Shoaling Test
4.8.5. Locomotion Activity Assay
4.9. RNA Sequencing and Data Analysis
4.10. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | Approach area |
| ASD | Autism spectrum disorder |
| CRISPR | Clustered regularly interspaced palindromic repeats |
| CS | Conspecific sector |
| CZRC | China Zebrafish Resource Center |
| DAG | Diacylglycerol |
| DEG | Differentially expressed genes |
| ES | Empty sector |
| FPKM | Fragments per kilobase per million mapped fragments |
| KEGG | Kyoto encyclopedia of genes and genomes |
| PCA | Principal component analysis |
| PKC | Protein kinase C |
| qPCR | Quantitative real-time PCR |
| RNA-seq | RNA sequencing |
References
- Oancea, E.; Meyer, T. Protein Kinase C as a Molecular Machine for Decoding Calcium and Diacylglycerol Signals. Cell 1998, 95, 307–318. [Google Scholar] [CrossRef]
- Webb, B.L.; Hirst, S.J.; Giembycz, M.A. Protein kinase C isoenzymes: A review of their structure, regulation and role in regulating airways smooth muscle tone and mitogenesis. Br. J. Pharmacol. 2000, 130, 1433–1452. [Google Scholar] [CrossRef]
- Newton, A.C. Protein-Kinase-C—Structure, Function, and Regulation. J. Biol. Chem. 1995, 270, 28495–28498. [Google Scholar]
- Nakanishi, H.; Exton, J.H. Purification and characterization of the zeta isoform of protein kinase C from bovine kidney. J. Biol. Chem. 1992, 267, 16347–16354. [Google Scholar] [CrossRef]
- Newton, A.C. Protein kinase C: Perfectly balanced. Crit. Rev. Biochem. Mol. Biol. 2018, 53, 208–230. [Google Scholar]
- Nakashima, S. Protein Kinase C (PKC): Regulation and Biological Function. J. Biochem. 2002, 132, 669–675. [Google Scholar] [CrossRef]
- Konopatskaya, O.; Poole, A.W. Protein kinase C alpha: Disease regulator and therapeutic target. Trends Pharmacol. Sci. 2010, 31, 8–14. [Google Scholar]
- Letiges, M. Knockout of PKC Enhances Insulin Signaling Through PI3K. Mol. Endocrinol. 2002, 16, 847–858. [Google Scholar] [CrossRef]
- Ruether, K.; Feigenspan, A.; Pirngruber, J.; Leitges, M.; Baehr, W.; Strauss, O. PKC alpha Is Essential for the Proper Activation and Termination of Rod Bipolar Cell Response. Investig. Oph. Thalmology Vis. Sci. 2010, 51, 6051–6058. [Google Scholar]
- Meisel, M.; Hermann-Kleiter, N.; Hinterleitner, R.; Gruber, T.; Wachowicz, K.; Pfeifhofer-Obermair, C.; Fresser, F.; Leitges, M.; Soldani, C.; Viola, A.; et al. The kinase PKCalpha selectively upregulates interleukin-17A during Th17 cell immune responses. Immunity 2013, 38, 41–52. [Google Scholar]
- Jakubcakova, V.; Oster, H.; Tamanini, F.; Cadenas, C.; Leitges, M.; van der Horst, G.T.; Eichele, G. Light Entrainment of the Mammalian Circadian Clock by a PRKCA-Dependent Posttranslational Mechanism. Neuron 2007, 54, 831–843. [Google Scholar] [CrossRef]
- Robles, M.S.; Boyault, C.; Knutti, D.; Padmanabhan, K.; Weitz, C.J. Identification of RACK1 and Protein Kinase C alpha as Integral Components of the Mammalian Circadian Clock. Science 2010, 327, 463–466. [Google Scholar]
- Nam, H.J.; Boo, K.; Kim, D.; Han, D.H.; Choe, H.K.; Kim, C.R.; Sun, W.; Kim, H.; Kim, K.; Lee, H.; et al. Phosphorylation of LSD1 by PKC alpha Is Crucial for Circadian Rhythmicity and Phase Resetting. Mol. Cell 2014, 53, 791–805. [Google Scholar]
- Walker, W.H.; Walton, J.C.; DeVries, A.C.; Nelson, R.J. Circadian rhythm disruption and mental health. Transl. Psychiatry 2020, 10, 28. [Google Scholar]
- Ahmad, M.; Din, N.S.B.; Tharumalay, R.D.; Din, N.C.; Ibrahim, N.; Amit, N.; Farah, N.M.; Osman, R.A.; Hamid, M.F.A.; Ibrahim, I.A.; et al. The Effects of Circadian Rhythm Disruption on Mental Health and Physiological Responses among Shift Workers and General Population. Int. J. Environ. Res. Public Health 2020, 17, 7156. [Google Scholar] [CrossRef]
- Coe, B.P.; Stessman, H.A.F.; Sulovari, A.; Geisheker, M.R.; Bakken, T.E.; Lake, A.M.; Dougherty, J.D.; Lein, E.S.; Hormozdiari, F.; Bernier, R.A.; et al. Neurodevelopmental disease genes implicated by de novo mutation and copy number variation morbidity. Nat. Genet. 2018, 51, 106–116. [Google Scholar] [CrossRef]
- Torshizi, A.D.; Wang, K. Tissue-wide cell-specific proteogenomic modeling reveals novel candidate risk genes in autism spectrum disorders. NPJ Syst. Biol. Appl. 2022, 8, 31. [Google Scholar] [CrossRef]
- Carroll, L.S.; Williams, N.M.; Moskvina, V.; Russell, E.; Norton, N.; Williams, H.; Peirce, T.; Georgieva, L.; Dwyer, S.; Grozeva, D.; et al. Evidence for rare and common genetic risk variants for schizophrenia at protein kinase C, alpha. Mol. Psychiatry 2009, 15, 1101–1111. [Google Scholar] [CrossRef] [Green Version]
- Stewart, A.M.; Nguyen, M.; Wong, K.; Poudel, M.K.; Kalueff, A.V. Developing zebrafish models of autism spectrum disorder (ASD). Prog. Neuro Psychopharmacol. Biol. Psychiatry 2014, 50, 27–36. [Google Scholar] [CrossRef]
- Meshalkina, D.A.; Kizlyk, M.N.; Kysil, E.V.; Collier, A.D.; Echevarria, D.J.; de Abreu, M.S.; Barcellos, L.J.G.; Song, C.; Warnick, J.E.; Kyzar, E.J.; et al. Zebrafish models of autism spectrum disorder. Exp. Neurol. 2018, 299, 207–216. [Google Scholar] [CrossRef]
- Tayanloo-Beik, A.; Hamidpour, S.K.; Abedi, M.; Shojaei, H.; Tavirani, M.R.; Namazi, N.; Larijani, B.; Arjmand, B. Zebrafish Modeling of Autism Spectrum Disorders, Current Status and Future Prospective. Front. Psychiatry 2022, 13, 1514. [Google Scholar] [CrossRef]
- Pensado-López, A.; Veiga-Rúa, S.; Carracedo, Á.; Allegue, C.; Sánchez, L. Experimental Models to Study Autism Spectrum Disorders: hiPSCs, Rodents and Zebrafish. Genes 2020, 11, 1376. [Google Scholar] [CrossRef]
- Cai, L.; Fisher, A.L.; Huang, H.; Xie, Z. CRISPR-mediated genome editing and human diseases. Genes Dis. 2016, 3, 244–251. [Google Scholar] [CrossRef]
- Guissart, C.; Latypova, X.; Rollier, P.; Khan, T.N.; Stamberger, H.; McWalter, K.; Cho, M.T.; Kjaergaard, S.; Weckhuysen, S.; Lesca, G.; et al. Dual Molecular Effects of Dominant RORA Mutations Cause Two Variants of Syndromic Intellectual Disability with Either Autism or Cerebellar Ataxia. Am. J. Hum. Genet. 2018, 102, 744–759. [Google Scholar] [CrossRef]
- Hamilton, T.J.; Morrill, A.; Lucas, K.; Gallup, J.; Harris, M.; Healey, M.; Pitman, T.; Schalomon, M.; Digweed, S.; Tresguerres, M. Establishing zebrafish as a model to study the anxiolytic effects of scopolamine. Sci. Rep. 2017, 7, 15081. [Google Scholar] [CrossRef]
- Cachat, J.M.; Canavello, P.R.; Elegante, M.F.; Bartels, B.K.; Elkhayat, S.I.; Hart, P.C.; Tien, A.K.; Tien, D.H.; Beeson, E.; Mohnot, S.; et al. Modeling Stress and Anxiety in Zebrafish. In Zebrafish Models in Neurobehavioral Research; Kalueff, A.V., Cachat, J.M., Eds.; Human Press: Saskatoon, SK, Canada, 2011; pp. 73–88. [Google Scholar]
- Supek, F.; Bošnjak, M.; Škunca, N.; Smuc, T. REVIGO Summarizes and Visualizes Long Lists of Gene Ontology Terms. PLoS ONE 2011, 6, e21800. [Google Scholar] [CrossRef]
- Buske, C.; Gerlai, R. Maturation of shoaling behavior is accompanied by changes in the dopaminergic and serotoninergic systems in zebrafish. Dev. Psychobiol. 2011, 54, 28–35. [Google Scholar] [CrossRef]
- Fu, C.-W.; Horng, J.-L.; Chou, M.-Y. Fish Behavior as a Neural Proxy to Reveal Physiological States. Front. Physiol. 2022, 13, 1420. [Google Scholar] [CrossRef]
- Santos, P.L.; Brito, R.G.; Matos, J.P.S.C.F.; Quintans, J.S.S.; Quintans-Júnior, L.J. Fos Protein as a Marker of Neuronal Activity: A Useful Tool in the Study of the Mechanism of Action of Natural Products with Analgesic Activity. Mol. Neurobiol. 2017, 55, 4560–4579. [Google Scholar] [CrossRef]
- Hoffman, G.E.; Smith, M.S.; Verbalis, J.G. C-Fos and Related Immediate-Early Gene-Products as Markers of Activity in Neuro-endocrine Systems. Front. Neuroendocrinol. 1993, 14, 173–213. [Google Scholar]
- Sun, X.; Lin, Y. Npas4: Linking Neuronal Activity to Memory. Trends Neurosci. 2016, 39, 264–275. [Google Scholar] [CrossRef]
- Ma, Y.; Usuwanthim, K.; Munawara, U.; Quach, A.; Gorgani, N.N.; Abbott, C.A.; Hii, C.S.; Ferrante, A. Protein Kinase C alpha Regulates the Expression of Complement Receptor Ig in Human Monocyte-Derived Macrophages. J. Immunol. 2015, 194, 2855–2861. [Google Scholar]
- Haus, E.; Smolensky, M.H. Biologic Rhythms in the Immune System. Chrono. Int. 1999, 16, 581–622. [Google Scholar] [CrossRef]
- Wang, C.; Lutes, L.K.; Barnoud, C.; Scheiermann, C. The circadian immune system. Sci. Immunol. 2022, 7, eabm2465. [Google Scholar] [CrossRef]
- Lazado, C.C.; Skov, P.V.; Pedersen, P.B. Innate immune defenses exhibit circadian rhythmicity and differential temporal sensitivity to a bacterial endotoxin in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 2016, 55, 613–622. [Google Scholar] [CrossRef]
- Topilko, P.; Schneider-Maunoury, S.; Levi, G.; Evercooren, A.B.-V.; Chennoufi, A.B.Y.; Seitanidou, T.; Babinet, C.; Charnay, P. Krox-20 controls myelination in the peripheral nervous system. Nature 1994, 371, 796–799. [Google Scholar] [CrossRef]
- Kim, S.H.; Song, J.Y.; Joo, E.J.; Lee, K.Y.; Shin, S.Y.; Lee, Y.H.; Ahn, Y.M.; Kim, Y.S. Genetic association of the EGR2 gene with bipolar disorder in Korea. Exp. Mol. Med. 2012, 44, 121–129. [Google Scholar] [CrossRef]
- Van der Veen, D.R.; van der Pol-Meijer, M.M.T.; Jansen, K.; Smeets, M.; Zee, E.A.V.D.; Gerkema, M.P. Circadian rhythms of C-FOS expression in the suprachiasmatic nuclei of the common vole (Microtus arvalis). Chronobiol. Int. 2008, 25, 481–499. [Google Scholar]
- Challet, E.; Jacob, N.; Vuillez, P.; Pévet, P.; Malan, A. Fos-like immunoreactivity in the circadian timing system of calorie-restricted rats fed at dawn: Daily rhythms and light pulse-induced changes. Brain Res. 1997, 770, 228–236. [Google Scholar] [CrossRef]
- Honrado, G.I.; Johnson, R.S.; Golombek, D.A.; Spiegelman, B.M.; Papaioannou, V.E.; Ralph, M.R. The circadian system of c-fos deficient mice. J. Comp. Physiol. A Neuroethol. Sens. Neural Behav. Physiol. 1996, 178, 563–570. [Google Scholar]
- Robins, M.T.; Li, J.; Ryabinin, A.E. Effects of Housing Conditions and Circadian Time on Baseline c-Fos Immunoreactivity in C57BL/6J Mice. Neuroscience 2020, 431, 143–151. [Google Scholar] [CrossRef]
- Xu, P.; Berto, S.; Kulkarni, A.; Jeong, B.; Joseph, C.; Cox, K.H.; Greenberg, M.E.; Kim, T.-K.; Konopka, G.; Takahashi, J.S. NPAS4 regulates the transcriptional response of the suprachiasmatic nucleus to light and circadian behavior. Neuron 2021, 109, 3268–3282. [Google Scholar] [CrossRef]
- Lissek, T.; Andrianarivelo, A.; Saint-Jour, E.; Allichon, M.; Bauersachs, H.G.; Nassar, M.; Piette, C.; Pruunsild, P.; Tan, Y.; Forget, B.; et al. Npas4 regulates medium spiny neuron physiology and gates cocaine-induced hyperlocomotion. EMBO Rep. 2021, 22, e51882. [Google Scholar] [CrossRef]
- Braz, J.C.; Gregory, K.; Pathak, A.; Zhao, W.; Sahin, B.; Klevitsky, R.; Kimball, T.F.; Lorenz, J.N.; Nairn, A.; Liggett, S.B.; et al. PKC-α regulates cardiac contractility and propensity toward heart failure. Nat. Med. 2004, 10, 248–254. [Google Scholar] [CrossRef]
- Ren, J.; Long, Y.; Liu, R.; Song, G.; Li, Q.; Cui, Z. Characterization of Biological Pathways Regulating Acute Cold Resistance of Zebrafish. Int. J. Mol. Sci. 2021, 22, 3028. [Google Scholar] [CrossRef]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef]
- Long, Y.; Li, X.; Li, F.; Ge, G.; Liu, R.; Song, G.; Li, Q.; Qiao, Z.; Cui, Z. Transcriptional Programs Underlying Cold Acclimation of Common Carp (Cyprinus carpio L.). Front. Genet. 2020, 11, 556418. [Google Scholar] [CrossRef]
- Liu, R.; Long, Y.; Liu, R.; Song, G.; Li, Q.; Yan, H.; Cui, Z. Understanding the Function and Mechanism of Zebrafish Tmem39b in Regulating Cold Resistance. Int. J. Mol. Sci. 2022, 23, 11442. [Google Scholar]
- Varshney, G.K.; Pei, W.; LaFave, M.C.; Idol, J.; Xu, L.; Gallardo, V.; Carrington, B.; Bishop, K.; Jones, M.; Li, M.; et al. High-throughput gene targeting and phenotyping in zebrafish using CRISPR/Cas9. Genome Res. 2015, 25, 1030–1042. [Google Scholar] [CrossRef]
- Cachat, J.; Stewart, A.; Grossman, L.; Gaikwad, S.; Kadri, F.; Chung, K.M.; Wu, N.; Wong, K.; Roy, S.; Suciu, C.; et al. Measuring behavioral and endocrine responses to novelty stress in adult zebrafish. Nat. Protoc. 2010, 5, 1786–1799. [Google Scholar] [CrossRef]
- Ikeda, A.; Komamizu, M.; Hayashi, A.; Yamasaki, C.; Okada, K.; Kawabe, M.; Komatsu, M.; Shiozaki, K. Neu1 deficiency induces abnormal emotional behavior in zebrafish. Sci. Rep. 2021, 11, 13477. [Google Scholar] [CrossRef]
- Rueden, C.T.; Schindelin, J.; Hiner, M.C.; Dezonia, B.E.; Walter, A.E.; Arena, E.T.; Eliceiri, K.W. ImageJ2: ImageJ for the next generation of scientific image data. BMC Bioinform. 2017, 18, 529. [Google Scholar] [CrossRef] [Green Version]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef]
- Liao, Y.; Smyth, G.K.; Shi, W. featureCounts: An efficient general purpose program for assigning sequence reads to ge-nomic features. Bioinformatics 2014, 30, 923–930. [Google Scholar]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar]
- Xu, J.; Kelly, R.; Fang, H.; Tong, W. ArrayTrack: A free FDA bioinformatics tool to support emerging biomedical research an update. Hum. Genom. 2010, 4, 428–434. [Google Scholar] [CrossRef]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef]
- Chin, C.-H.; Chen, S.-H.; Wu, H.-H.; Ho, C.-W.; Ko, M.-T.; Lin, C.-Y. cytoHubba: Identifying hub objects and sub-networks from complex interactome. BMC Syst. Biol. 2014, 8, S11. [Google Scholar] [CrossRef]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiment | Purpose |
|---|---|
| Novel tank test | To analyze anxiety-like behavior and exploratory activity of the experimental fish |
| Mirror biting test | To analyze emotional activities such as the sociality, aggressiveness and boldness of the experimental fish |
| Social preference test | To analyze the preference of the experimental fish to conspecifics |
| Shoaling test | To assess the social cohesion of the homogeneous group of the experimental fish |
| Locomotion activity assay | To assess the locomotion rhythmicity of the experimental fish |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, H.; Long, Y.; Song, G.; Chen, S.; Xu, Z.; Li, Q.; Wu, Z. Dysfunction of Prkcaa Links Social Behavior Defects with Disturbed Circadian Rhythm in Zebrafish. Int. J. Mol. Sci. 2023, 24, 3849. https://doi.org/10.3390/ijms24043849
Hu H, Long Y, Song G, Chen S, Xu Z, Li Q, Wu Z. Dysfunction of Prkcaa Links Social Behavior Defects with Disturbed Circadian Rhythm in Zebrafish. International Journal of Molecular Sciences. 2023; 24(4):3849. https://doi.org/10.3390/ijms24043849
Chicago/Turabian StyleHu, Han, Yong Long, Guili Song, Shaoxiong Chen, Zhicheng Xu, Qing Li, and Zhengli Wu. 2023. "Dysfunction of Prkcaa Links Social Behavior Defects with Disturbed Circadian Rhythm in Zebrafish" International Journal of Molecular Sciences 24, no. 4: 3849. https://doi.org/10.3390/ijms24043849