
Space Omics and Tissue Response in Astronaut Skeletal Muscle after Short and Long Duration Missions
 , , , , , , , , and
, , , , , , , , and 
Abstract
:1. Introduction
2. Results
2.1. Anthropometric Data
2.2. Training Protocol
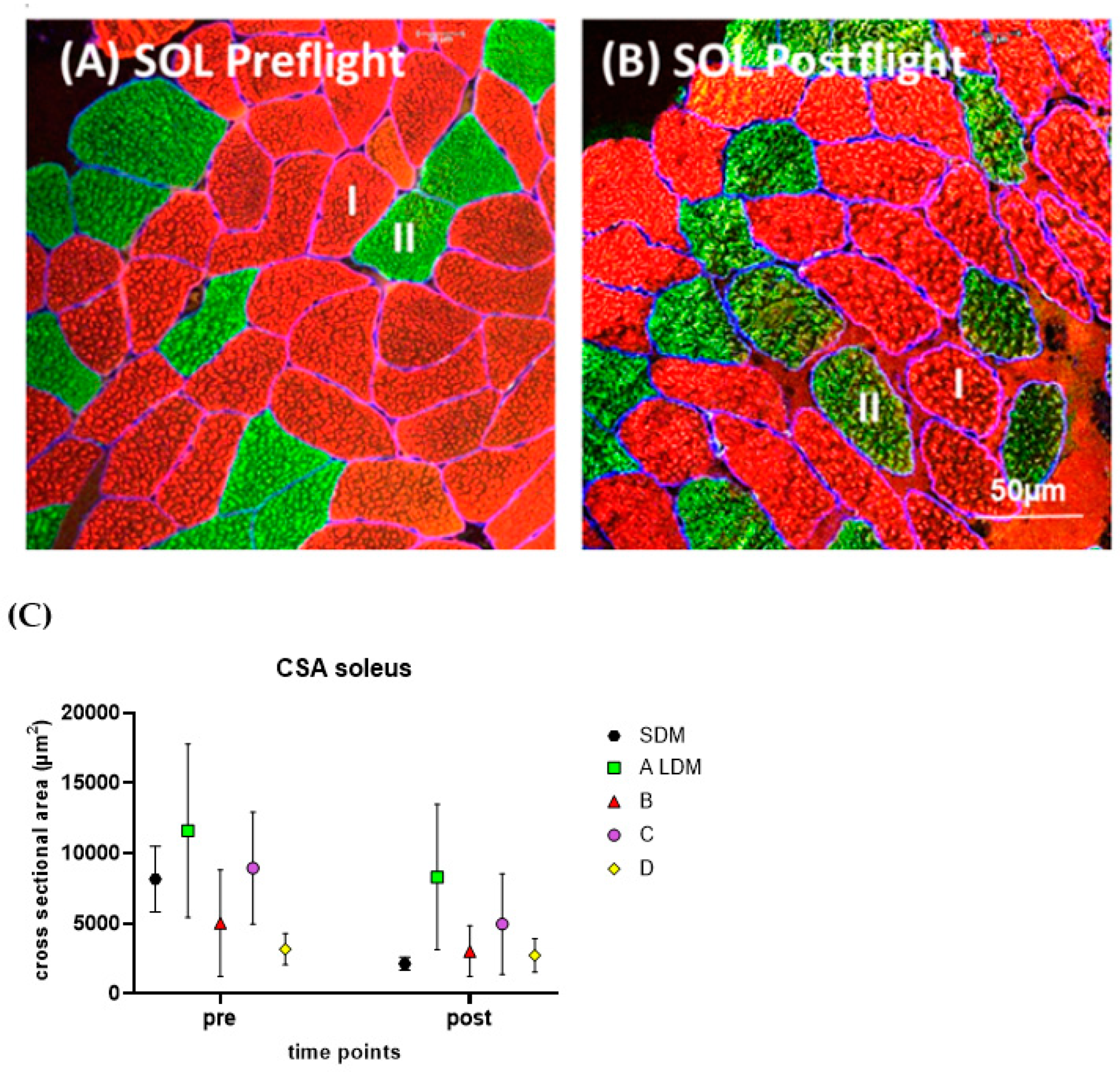
2.3. Myofiber Size (Cross-Sectional Area, CSA) Analysis
2.4. Histology and Extracellular Matrix Immunomarkers
2.5. Proteomic Profile in Acute SDM (9/11 Days ISS Mission) and Chronic LDM Exposure (>6 Months ISS Mission) to Microgravity
2.5.1. Structural/Contractile Proteins and Proteins Involved in Calcium Signaling
2.5.2. Change in Metabolic Protein Abundance According to Glycolysis, TCA Cycle and Mitochondrial Respiratory Chain
2.5.3. Lipid Metabolism
2.5.4. Behavior of Transport, High-Energy Phosphate Interconversion, Stress and Others Proteins
3. Discussion
Study Limitations
4. Materials and Methods
4.1. Study Design
4.2. Muscle Biopsy
4.3. Inflight Countermeasures (CM)
4.4. Histology and Immunohistochemistry
4.5. Protein Extraction for Label-Free LC–ESI–MS/MS Analysis
4.6. Label-Free Liquid Chromatography with Tandem Mass Spectrometry
4.7. Ingenuity Pathway Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACAA2 | 3-ketoacyl-CoA thiolase |
| ACADM | medium-chain specific acyl-CoA dehydrogenase |
| ACADVL | very long-chain specific acyl-CoA dehydrogenase |
| ACAT1 | acetyl-CoA acetyltransferase |
| ACO2 | aconitate hydratase |
| ACOT1 | acyl-coenzyme A thioesterase 1 |
| ACSL1 | long-chain-fatty-acid--CoA ligase 1 |
| ACTA1 | actin, alpha skeletal muscle |
| ACTN2 | alpha-actinin-2 |
| ADSSL1 | adenylosuccinate synthetase isozyme 1 |
| AGL | glycogen debranching enzyme |
| AK1 | adenylate kinase isoenzyme 1 |
| ALDH1A1 | retinal dehydrogenase 1 |
| ALDH4A1 | delta-1-pyrroline-5-carboxylate dehydrogenase |
| ALDOA | fructose-bisphosphate aldolase A |
| ANKRD2 | ankyrin repeat domain-containing protein 2 |
| ANXA2 | annexin A2 |
| APOA1 | apolipoprotein A-I |
| APOA2 | apolipoprotein A-II |
| ARED | advanced resistive exercise device (exercise device, weight lifts) |
| ATP2A1 | sarco(endo)plasmatic reticulum Ca(2+)-ATPase 1 (SERCA 1) pump |
| ATP5A1 | ATP synthase subunit alpha |
| ATP5B | ATP synthase subunit beta |
| ATP5C1 | ATP synthase subunit gamma |
| ATP5D | ATP synthase subunit delta |
| ATP5H | ATP synthase subunit d |
| ATP5J | ATP synthase-coupling factor 6 |
| ATP5J2 | ATP synthase subunit f |
| BCL2L13 | Bcl-2-like protein 13 |
| BMI | body mass index |
| CA3 | carbonic anhydrase 3 |
| CASQ1/CASQ2 | calsequestrin-1/2 |
| CEVIS | cycle ergometer (exercise device, -> heart rate blood circulation) |
| CFL2 | cofilin-2 |
| CISD1 | CDGSH iron-sulfur domain-containing protein |
| CKB | creatine kinase B-type |
| CKM | creatine kinase M-type |
| CKMT2 | creatine kinase S-type |
| CM | countermeasure |
| CMBL | carboxymethylene butenolidase homolog |
| COL6A1 | collagen alpha-1(VI) chain |
| COL6A3 | collagen alpha-3(VI) chain |
| COQ9 | ubiquinone biosynthesis protein COQ9, mitochondrial |
| COX4I1 | cytochrome c oxidase subunit 4 isoform |
| COX5B | cytochrome c oxidase subunit 5B |
| COX6B1 | cytochrome c oxidase subunit 6B1 |
| CS | citrate synthase |
| CSA | cross sectional area (myofiber size) |
| CYB5R1 | NADH-cytochrome b5 reductase 1 |
| CYB5R3 | NADH-cytochrome b5 reductase 3 |
| CYCS | cytochrome C |
| DECR1 | 2,4-dienoyl-CoA reductase |
| DLD | dihydrolipoyl dehydrogenase |
| DLST | dihydrolipoyllysine-residue succinyltransferase component of 2-oxoglutarate dehydrogenase complex |
| DPYSL3 | dihydropyrimidinase-related protein 3 |
| ECH1 | delta(3,5)-Delta(2,4)-dienoyl-CoA isomerase |
| ECHS1 | enoyl-CoA hydratase |
| ECI2 | enoyl-CoA delta isomerase 2 |
| ECM | extracellular matrix |
| EDL | extensor digitorum longus (plantarflexor muscle) |
| EHD2 | EH domain-containing protein 2 |
| EIF5A | eukaryotic translation initiation factor 5A-1 |
| ENO1 | alpha-enolase |
| EPHX1 | epoxide hydrolase 1 |
| ER | endoplasmic reticulum (muscle fiber organelle) |
| ETFA/B | electron transfer flavoprotein subunit alpha/beta |
| ETFDH | electron transfer flavoprotein-ubiquinone oxidoreductase |
| FABP3 | fatty-acid binding protein |
| FBN1 | fibrillin 1 |
| FD | flight days |
| FGA/FGB | fibrinogen alpha/beta chain |
| FH | fumarate hydratase |
| FHL1 | four and a half LIM domains protein 1 |
| FLNA | filamin-A |
| FTH1 | ferritin heavy chain |
| GATD3 | glutamine amidotransferase-like class 1 domain-containing protein 3 |
| GC | vitamin D-binding protein |
| GDI2 | Rab GDP dissociation inhibitor beta |
| GLO1 | lactoylglutathione lyase |
| GOT1/GOT2 | aspartate aminotransferase, cytoplasmic/mitochondrial |
| GPD1 | glycerol-3-phosphate dehydrogenase [NAD(+)], cytoplasmic |
| GPD1L | glycerol-3-phosphate dehydrogenase 1-like protein |
| GPT | alanine aminotransferase 1 |
| GSTO1 | Glutathione S-transferase omega-1 |
| GYS1 | glycogen [starch] synthase |
| H&E | hematoxylin-eosin (histology stain) |
| HADH | hydroxyacyl-coenzyme A dehydrogenase |
| HADHA | trifunctional enzyme subunit alpha |
| HHATL | protein-cysteine N-palmitoyltransferase HHAT-like protein |
| HP | haptoglobin |
| HSDL2 | hydroxysteroid dehydrogenase-like protein 2 |
| HSPB6 | heat shock protein beta-6 |
| HSPD1 | 60 kDa heat shock protein |
| IDH2 | isocitrate dehydrogenase [NADP] |
| ILK | integrin-linked kinase |
| IMMT | MICOS complex subunit MIC60 |
| ISS | International Space Station |
| IPA | ingenuity pathway analysis |
| LC–ESI–MS/MS | liquid chromatography electrospray/tandem mass spectrometry |
| µG/0G | microgravity [µG] means micro-fractional and/or zero net gravitational force [0G] |
| LDHA | L-lactate dehydrogenase A chain |
| LDHB | L-lactate dehydrogenase B chain |
| LDM | long-term mission |
| LMCD1 | LIM and cysteine-rich domains protein 1 |
| LUM | lumican |
| MAP4 | microtubule-associated protein 4 |
| MB | myoglobin |
| MDH1 | malate dehydrogenase, cytoplasmic |
| MMP2 | matrix metalloproteinase 2 |
| MSN | moesin |
| MT-CO2 | cytochrome c oxidase subunit 2 |
| MYBPC1 | myosin-binding protein C (slow type) |
| MYH2 | myosin-2 |
| MYH7 | myosin-7 |
| MYL1 | myosin light chain 1/3 |
| MYL2 | myosin regulatory light chain 2 (slow-type myofibers) |
| MYL3 | myosin light chain-3 |
| MYL6B | myosin light chain-6B |
| MYLPF | myosin regulatory light chain 2 |
| MYOM2 | myomesin-2 |
| NDUFA5 | NADH dehydrogenase [ubiquinone] 1 alpha subcomplex subunit 5 |
| NDUFB10 | NADH dehydrogenase [ubiquinone] 1 beta subcomplex subunit 10 |
| NDUFB8 | NADH dehydrogenase [ubiquinone] 1 beta subcomplex subunit 8 |
| NDUFS1 | NADH-ubiquinone oxidoreductase 75 kDa subunit |
| NDUFS2 | NADH dehydrogenase [ubiquinone] iron-sulfur protein 2 |
| NDUFS5 | NADH dehydrogenase [ubiquinone] iron-sulfur protein 5 |
| NDUFS8 | NADH dehydrogenase [ubiquinone] iron-sulfur protein 8 |
| NEB | nebulin |
| NME2 | nucleoside diphosphate kinase B |
| NNT | NAD(P) transhydrogenase |
| OGDH | 2-oxoglutarate dehydrogenase |
| PDHA1 | pyruvate dehydrogenase E1 component subunit alpha |
| PDLIM3 | PDZ LIM domain protein 3 |
| PFKM | ATP-dependent 6-phosphofructokinase |
| PHB | prohibitin |
| PKM | pyruvate kinase |
| PLEC | plectin |
| PPIA | peptidyl-prolyl cis-trans isomerase A |
| PRDX1 | peroxiredoxin-1 |
| PRDX3 | thioredoxin-dependent peroxide reductase |
| PRE | preflight |
| PRKACA | cAMP-dependent protein kinase catalytic subunit alpha |
| PSMA7 | proteasome subunit alpha type-7 |
| PTGES2 | prostaglandin E synthase 2 |
| R+0 | day of return to Earth |
| RHOA | Rho A GTPase |
| RTN4 | reticulon-4 |
| RYR1 | ryanodine receptor 1 |
| S100A1 | protein S100-A1 |
| SDHA | succinate dehydrogenase [ubiquinone] flavoprotein subunit |
| SDM | short-term mission |
| SERPINC1 | antithrombin-III |
| SH3BGR | SH3 domain-binding glutamic acid-rich protein |
| SLC25A11 | Mitochondrial 2-oxoglutarate/malate carrier protein |
| SLC25A12 | Calcium-binding mitochondrial carrier protein Aralar1 |
| SLC25A4 | ADP/ATP translocase 1 |
| SLC4A1 | band 3 anion transport protein |
| SOD2 | superoxid dismutase [Mn] 2 |
| SOL | soleus muscle (deep calf postural muscle) |
| SRL | sarcalumenin |
| SUCLA2 | succinyl-CoA ligase [ADP-forming] subunit beta |
| SYNM | synemin |
| SYPL2 | synaptophysin-like protein 2 |
| T2 | treadmill 2 (exercise device, endurance) |
| TCA | tricarboxylic acid |
| TF | serotransferrin |
| TNNI1 | troponin I, slow skeletal muscle |
| TPI1 | triosephosphate isomerase |
| TPM2 | tropomyosin beta chain |
| TRIM72 | tripartite motif-containing protein 72 |
| TTN | titin |
| TUBB | tubulin beta chain |
| TUBB4B | tubulin beta-4B chain |
| TUFM | elongation factor Tu |
| UGP2 | UTP--glucose-1-phosphate uridylyltransferase |
| UQCRC1/UQCRC2 | cytochrome b-c1 complex subunit 1/2 |
| UQCRQ | cytochrome b-c1 complex subunit 8 |
| VCL | vinculin |
| VDAC1/2/3 | voltage-dependent anion-selective channel protein 1/2/3 |
| VIM | vimentin |
| w/o | without |
| YBX3 | Y-box-binding protein 3 |
| YWHAZ | 14-3-3 protein zeta/delta |
References
- Demontis, G.C.; Germani, M.M.; Caiani, E.G.; Barravecchia, I.; Passino, C.; Angeloni, D. Human Pathophysiological Adaptations to the Space Environment. Front. Physiol. 2017, 8, 547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juhl, O.J.; Buettmann, E.G.; Friedman, M.A.; DeNapoli, R.C.; Hoppock, G.A.; Donahue, H.J. Update on the effects of microgravity on the musculoskeletal system. npj Microgravity 2021, 7, 28. [Google Scholar] [CrossRef] [PubMed]
- Grimm, D.; Grosse, J.; Wehland, M.; Mann, V.; Reseland, J.E.; Sundaresan, A.; Corydon, T.J. The impact of microgravity on bone in humans. Bone 2016, 87, 44–56. [Google Scholar] [CrossRef] [PubMed]
- Tagliaferri, C.; Wittrant, Y.; Davicco, M.-J.; Walrand, S.; Coxam, V. Muscle and bone, two interconnected tissues. Ageing Res. Rev. 2015, 21, 55–70. [Google Scholar] [CrossRef] [PubMed]
- Vandenburgh, H.; Chromiak, J.; Shansky, J.; Del Tatto, M.; Lemaire, J. Space travel directly induces skeletal muscle atrophy. FASEB J. 1999, 13, 1031–1038. [Google Scholar] [CrossRef] [PubMed]
- English, K.L.; Lee, S.M.; Loehr, J.A.; Ploutz–Snyder, R.J.; Ploutz–Snyder, L.L. Isokinetic Strength Changes Following Long-Duration Spaceflight on the ISS. Aerosp. Med. Hum. Perform. 2015, 86, A68–A77. [Google Scholar] [CrossRef]
- Fitts, R.H.; Trappe, S.W.; Costill, D.L.; Gallagher, P.M.; Creer, A.C.; Colloton, P.A.; Peters, J.R.; Romatowski, J.G.; Bain, J.L.; Riley, D.A. Prolonged space flight-induced alterations in the structure and function of human skeletal muscle fibres. J. Physiol. 2010, 588, 3567–3592. [Google Scholar] [CrossRef]
- Genc, K.; Gopalakrishnan, R.; Kuklis, M.; Maender, C.; Rice, A.; Bowersox, K.; Cavanagh, P. Foot forces during exercise on the International Space Station. J. Biomech. 2010, 43, 3020–3027. [Google Scholar] [CrossRef]
- LeBlanc, A.; Lin, C.; Shackelford, L.; Sinitsyn, V.; Evans, H.; Belichenko, O.; Schenkman, B.; Kozlovskaya, I.; Oganov, V.; Bakulin, A.; et al. Muscle volume, MRI relaxation times (T2), and body composition after spaceflight. J. Appl. Physiol. 2000, 89, 2158–2164. [Google Scholar] [CrossRef]
- Trappe, S.; Costill, D.; Gallagher, P.; Creer, A.; Peters, J.R.; Evans, H.; Riley, D.A.; Fitts, R.H.; Begue, G.; Raue, U.; et al. Exercise in space: Human skeletal muscle after 6 months aboard the International Space Station. J. Appl. Physiol. 2009, 106, 1159–1168. [Google Scholar] [CrossRef] [Green Version]
- Leblanc, A.; Schneider, V.; Shackelford, L.; West, S.; Oganov, V.; Bakulin, A.; Voronin, L. Bone mineral and lean tissue loss after long duration space flight. J. Musculoskelet. Neuronal Interact. 2000, 1, 157–160. [Google Scholar] [PubMed]
- Blaber, E.; Dvorochkin, N.; Torres, M.; Yousuf, R.; Burns, B.; Globus, R.; Almeida, E. Mechanical unloading of bone in microgravity reduces mesenchymal and hematopoietic stem cell-mediated tissue regeneration. Stem Cell Res. 2014, 13, 181–201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cowin, S. On Mechanosensation in Bone under Microgravity. Bone 1998, 22, 119S–125S. [Google Scholar] [CrossRef] [PubMed]
- Lang, T.; Van Loon, J.J.; Bloomfield, S.; Vico, L.; Chopard, A.; Rittweger, J.; Kyparos, A.; Blottner, D.; Vuori, I.; Gerzer, R.; et al. Towards human exploration of space: The THESEUS review series on muscle and bone research priorities. npj Microgravity 2017, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Belavý, D.L.; Bock, O.; Börst, H.; Armbrecht, G.; Gast, U.; Degner, C.; Beller, G.; Soll, H.; Salanova, M.; Habazettl, H.; et al. The 2nd Berlin BedRest Study: Protocol and implementation. J. Musculoskelet. Neuronal Interact. 2010, 10, 207–219. [Google Scholar]
- Bergström, J.; Hermansen, L.; Hultman, E.; Saltin, B. Diet, Muscle Glycogen and Physical Performance. Acta Physiol. Scand. 1967, 71, 140–150. [Google Scholar] [CrossRef]
- Evans, W.J.; Phinney, S.D.; Young, V.R. Suction applied to a muscle biopsy maximizes sample size. Med. Sci. Sports Exerc. 1982, 14, 101–102. [Google Scholar] [CrossRef]
- Baldwin, K.M. Effect of spaceflight on the functional, biochemical, and metabolic properties of skeletal muscle. Med. Sci. Sports Exerc. 1996, 28, 983–987. [Google Scholar] [CrossRef]
- Fitts, R.H.; Riley, D.R.; Widrick, J.J. Functional and structural adaptations of skeletal muscle to microgravity. J. Exp. Biol. 2001, 204, 3201–3208. [Google Scholar] [CrossRef]
- Rittweger, J.; Albracht, K.; Flück, M.; Ruoss, S.; Brocca, L.; Longa, E.; Moriggi, M.; Seynnes, O.; Di Giulio, I.; Tenori, L.; et al. Sarcolab pilot study into skeletal muscle’s adaptation to long-term spaceflight. npj Microgravity 2018, 4, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Bs, B.B.K.; Riley, D.A. Distinguishing unloading. Versus reloading-induced changes in rat soleus muscle. Muscle Nerve 1993, 16, 99–108. [Google Scholar] [CrossRef]
- Hackney, K.J.; Scott, J.M.; Hanson, A.M.; English, K.L.; Downs, M.E.; Ploutz-Snyder, L.L. The Astronaut-Athlete: Optimizing Human Performance in Space. J. Strength Cond. Res. 2015, 29, 3531–3545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Padilha, C.S.; Figueiredo, C.; Deminice, R.; Krüger, K.; Seelaender, M.; Rosa-Neto, J.C.; Lira, F.S. Costly immunometabolic remodelling in disused muscle buildup through physical exercise. Acta Physiol. 2022, 234, e13782. [Google Scholar] [CrossRef] [PubMed]
- Gundersen, K. Excitation-transcription coupling in skeletal muscle: The molecular pathways of exercise. Biol. Rev. 2011, 86, 564–600. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zhang, L.; Zhang, L.; Han, R. Autophagy in striated muscle diseases. Front. Cardiovasc. Med. 2022, 9, 1000067. [Google Scholar] [CrossRef]
- Franco-Romero, A.; Sandri, M. Role of autophagy in muscle disease. Mol. Asp. Med. 2021, 82, 101041. [Google Scholar] [CrossRef]
- Yamashita, S.; Kyuuma, M.; Inoue, K.; Hata, Y.; Kawada, R.; Yamabi, M.; Fujii, Y.; Sakagami, J.; Fukuda, T.; Furukawa, K.; et al. Mitophagy reporter mouse analysis reveals increased mitophagy activity in disuse-induced muscle atrophy. J. Cell Physiol. 2021, 236, 7612–7624. [Google Scholar] [CrossRef]
- Suetta, C.; Frandsen, U.; Jensen, L.; Jensen, M.M.; Jespersen, J.G.; Hvid, L.G.; Bayer, M.; Petersson, S.J.; Schrøder, H.D.; Andersen, J.L.; et al. Aging Affects the Transcriptional Regulation of Human Skeletal Muscle Disuse Atrophy. PLoS ONE 2012, 7, e51238. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Yuan, W.; Chen, Y.; Lin, B.; Wang, S.; Deng, Z.; Zheng, Q.; Li, Q. Transcription and proteome changes involved in re-innervation muscle following nerve crush in rats. BMC Genom. 2022, 23, 666. [Google Scholar] [CrossRef]
- Dirksen, R.T.; Eisner, D.A.; Ríos, E.; Sipido, K.R. Excitation–contraction coupling in cardiac, skeletal, and smooth muscle. J. Gen. Physiol. 2022, 154, e202213244. [Google Scholar] [CrossRef]
- Rossi, D.; Gamberucci, A.; Pierantozzi, E.; Amato, C.; Migliore, L.; Sorrentino, V. Calsequestrin, a key protein in striated muscle health and disease. J. Muscle Res. Cell Motil. 2020, 42, 267–279. [Google Scholar] [CrossRef]
- Allen, D.G.; Lamb, G.D.; Westerblad, H. Impaired calcium release during fatigue. J. Appl. Physiol. 2008, 104, 296–305. [Google Scholar] [CrossRef] [PubMed]
- Khamoui, A.V.; Tokmina-Roszyk, D.; Feresin, R.G.; Fields, G.B.; Visavadiya, N.P. Skeletal muscle proteome expression differentiates severity of cancer cachexia in mice and identifies loss of fragile X mental retardation syndrome-related protein 1. Proteomics 2022, 22, e2100157. [Google Scholar] [CrossRef] [PubMed]
- Moriggi, M.; Vasso, M.; Fania, C.; Capitanio, D.; Bonifacio, G.; Salanova, M.; Blottner, D.; Rittweger, J.; Felsenberg, D.; Cerretelli, P.; et al. Long term bed rest with and without vibration exercise countermeasures: Effects on human muscle protein dysregulation. Proteomics 2010, 10, 3756–3774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Castro-Sepulveda, M.; Jannas-Vela, S.; Fernández-Verdejo, R.; Ávalos-Allele, D.; Tapia, G.; Villagrán, C.; Quezada, N.; Zbinden-Foncea, H. Relative lipid oxidation associates directly with mitochondrial fusion phenotype and mitochondria-sarcoplasmic reticulum interactions in human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2020, 318, E848–E855. [Google Scholar] [CrossRef]
- Al Saedi, A.; Debruin, D.A.; Hayes, A.; Hamrick, M. Lipid metabolism in sarcopenia. Bone 2022, 164, 116539. [Google Scholar] [CrossRef] [PubMed]
- Coen, P.M.; Goodpaster, B.H. Role of intramyocelluar lipids in human health. Trends Endocrinol. Metab. 2012, 23, 391–398. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salanova, M.; Gambara, G.; Moriggi, M.; Vasso, M.; Ungethuem, U.; Belavý, D.L.; Felsenberg, D.; Cerretelli, P.; Gelfi, C.; Blottner, D. Vibration mechanosignals superimposed to resistive exercise result in baseline skeletal muscle transcriptome profiles following chronic disuse in bed rest. Sci. Rep. 2015, 5, 17027. [Google Scholar] [CrossRef] [Green Version]
- Komiya, Y.; Sawano, S.; Mashima, D.; Ichitsubo, R.; Nakamura, M.; Tatsumi, R.; Ikeuchi, Y.; Mizunoya, W. Mouse soleus (slow) muscle shows greater intramyocellular lipid droplet accumulation than EDL (fast) muscle: Fiber type-specific analysis. J. Muscle Res. Cell Motil. 2017, 38, 163–173. [Google Scholar] [CrossRef]
- Seibert, J.T.; Najt, C.P.; Heden, T.D.; Mashek, D.G.; Chow, L.S. Muscle Lipid Droplets: Cellular Signaling to Exercise Physiology and Beyond. Trends Endocrinol. Metab. 2020, 31, 928–938. [Google Scholar] [CrossRef]
- Chambers, M.A.; Moylan, J.S.; Reid, M.B. Physical inactivity and muscle weakness in the critically ill. Crit. Care Med. 2009, 37, S337–S346. [Google Scholar] [CrossRef] [PubMed]
- Barbacini, P.; Blottner, D.; Capitanio, D.; Trautmann, G.; Block, K.; Torretta, E.; Moriggi, M.; Salanova, M.; Gelfi, C. Effects of Omega-3 and Antioxidant Cocktail Supplement on Prolonged Bed Rest: Results from Serum Proteome and Sphingolipids Analysis. Cells 2022, 11, 2120. [Google Scholar] [CrossRef] [PubMed]
- Libera, L.D.; Ravara, B.; Gobbo, V.; Tarricone, E.; Vitadello, M.; Biolo, G.; Vescovo, G.; Gorza, L. A transient antioxidant stress response accompanies the onset of disuse atrophy in human skeletal muscle. J. Appl. Physiol. 2009, 107, 549–557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baba, S.; Smith, T.; Hellmann, J.; Bhatnagar, A.; Carter, K.; Vanhoover, A.; Caruso, J. Space Flight Diet-Induced Deficiency and Response to Gravity-Free Resistive Exercise. Nutrients 2020, 12, 2400. [Google Scholar] [CrossRef] [PubMed]
- Bergouignan, A.; Stein, T.P.; Habold, C.; Coxam, V.; O’Gorman, D.; Blanc, S. Towards human exploration of space: The THESEUS review series on nutrition and metabolism research priorities. npj Microgravity 2016, 2, 16029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heer, M.; Boerger, A.; Kamps, N.; Mika, C.; Korr, C.; Drummer, C. Nutrient supply during recent European missions. Pflug. Arch. 2000, 441, R8–R14. [Google Scholar] [CrossRef]
- Smith, S.M.; Zwart, S.R.; Block, G.; Rice, B.L.; Davis-Street, J.E. The Nutritional Status of Astronauts Is Altered after Long-Term Space Flight Aboard the International Space Station. J. Nutr. 2005, 135, 437–443. [Google Scholar] [CrossRef] [Green Version]
- Leblanc, A.; Rowe, R.; Schneider, V.; Evans, H.; Hedrick, T. Regional muscle loss after short duration spaceflight. Aviat. Space Environ. Med. 1995, 66, 1151–1154. [Google Scholar]
- Purslow, P.P. The Structure and Role of Intramuscular Connective Tissue in Muscle Function. Front. Physiol. 2020, 11, 495. [Google Scholar] [CrossRef]
- Fede, C.; Fan, C.; Pirri, C.; Petrelli, L.; Biz, C.; Porzionato, A.; Macchi, V.; De Caro, R.; Stecco, C. The Effects of Aging on the Intramuscular Connective Tissue. Int. J. Mol. Sci. 2022, 23, 11061. [Google Scholar] [CrossRef]
- Wynn, T.A. Common and unique mechanisms regulate fibrosis in various fibroproliferative diseases. J. Clin. Investig. 2007, 117, 524–529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schoser, B.G.H.; Blottner, D.; Stuerenburg, H.-J. Matrix metalloproteinases in inflammatory myopathies: Enhanced immunoreactivity near atrophic myofibers. Acta Neurol. Scand. 2002, 105, 309–313. [Google Scholar] [CrossRef] [PubMed]
- Thot, G.K.; Berwanger, C.; Mulder, E.; Lee, J.K.; Lichterfeld, Y.; Ganse, B.; Giakoumaki, I.; Degens, H.; Duran, I.; Schönau, E.; et al. Effects of long-term immobilisation on endomysium of the soleus muscle in humans. Exp. Physiol. 2021, 106, 2038–2045. [Google Scholar] [CrossRef] [PubMed]
- Elmann-Larsen, B.; Schmitt, D. Staying in bed to benefit ESA’s astronauts and Europe’s citizens. ESA Bull 2003, 113, 34–39. [Google Scholar]
- Kenny, H.C.; Tascher, G.; Ziemianin, A.; Rudwill, F.; Zahariev, A.; Chery, I.; Gauquelin-Koch, G.; Barielle, M.-P.; Heer, M.; Blanc, S.; et al. Effectiveness of Resistive Vibration Exercise and Whey Protein Supplementation Plus Alkaline Salt on the Skeletal Muscle Proteome Following 21 Days of Bed Rest in Healthy Males. J. Proteome Res. 2020, 19, 3438–3451. [Google Scholar] [CrossRef]
- Nielsen, R.O.; Schjerling, P.; Tesch, P.; Stål, P.; Langberg, H. Collagen content in the vastus lateralis and the soleus muscle following a 90-day bed rest period with or without resistance exercises. Muscles Ligaments Tendons J. 2015, 5, 305–309. [Google Scholar] [CrossRef]
- Parry, S.M.; Puthucheary, Z.A. The impact of extended bed rest on the musculoskeletal system in the critical care environment. Extreme Physiol. Med. 2015, 4, 16. [Google Scholar] [CrossRef] [Green Version]
- Argilés, J.M.; Campos, N.; Lopez-Pedrosa, J.M.; Rueda, R.; Rodriguez-Mañas, L. Skeletal Muscle Regulates Metabolism via Interorgan Crosstalk: Roles in Health and Disease. J. Am. Med. Dir. Assoc. 2016, 17, 789–796. [Google Scholar] [CrossRef] [Green Version]
- Lemoine, J.K.; Haus, J.M.; Trappe, S.W.; Trappe, T.A. Muscle proteins during 60-day bedrest in women: Impact of exercise or nutrition. Muscle Nerve 2009, 39, 463–471. [Google Scholar] [CrossRef]
- Evans, W.J. Skeletal muscle loss: Cachexia, sarcopenia, and inactivity. Am. J. Clin. Nutr. 2010, 91, 1123S–1127S. [Google Scholar] [CrossRef] [Green Version]
- Capri, M.; Morsiani, C.; Santoro, A.; Moriggi, M.; Conte, M.; Martucci, M.; Bellavista, E.; Fabbri, C.; Giampieri, E.; Albracht, K.; et al. Recovery from 6-month spaceflight at the International Space Station: Muscle-related stress into a proinflammatory setting. FASEB J. 2019, 33, 5168–5180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blottner, D.; Bosutti, A.; Degens, H.; Schiffl, G.; Gutsmann, M.; Buehlmeier, J.; Rittweger, J.; Ganse, B.; Heer, M.; Salanova, M. Whey protein plus bicarbonate supplement has little effects on structural atrophy and proteolysis marker immunopatterns in skeletal muscle disuse during 21 days of bed rest. J. Musculoskelet. Neuronal Interact. 2014, 14, 432–444. [Google Scholar] [PubMed]
- Loehr, J.A.; Guilliams, M.E.; Petersen, N.; Hirsch, N.; Kawashima, S.; Ohshima, H. Physical Training for Long-Duration Spaceflight. Aerosp. Med. Hum. Perform. 2015, 86, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Petersen, N.; Jaekel, P.; Rosenberger, A.; Weber, T.; Scott, J.; Castrucci, F.; Lambrecht, G.; Ploutz-Snyder, L.; Damann, V.; Kozlovskaya, I.; et al. Exercise in space: The European Space Agency approach to in-flight exercise countermeasures for long-duration missions on ISS. Extreme Physiol. Med. 2016, 5, 9. [Google Scholar] [CrossRef] [Green Version]
- Blottner, D.; Hastermann, M.; Weber, R.; Lenz, R.; Gambara, G.; Limper, U.; Rittweger, J.; Bosutti, A.; Degens, H.; Salanova, M. Reactive Jumps Preserve Skeletal Muscle Structure, Phenotype, and Myofiber Oxidative Capacity in Bed Rest. Front. Physiol. 2020, 10, 1527. [Google Scholar] [CrossRef] [Green Version]
- Wiśniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef]
- Capitanio, D.; Moriggi, M.; Barbacini, P.; Torretta, E.; Moroni, I.; Blasevich, F.; Morandi, L.; Mora, M.; Gelfi, C. Molecular Fingerprint of BMD Patients Lacking a Portion in the Rod Domain of Dystrophin. Int. J. Mol. Sci. 2022, 23, 2624. [Google Scholar] [CrossRef]
- Cox, J.; Mann, M. MaxQuant enables high peptide identification rates, individualized p.p.b.-range mass accuracies and proteome-wide protein quantification. Nat. Biotechnol. 2008, 26, 1367–1372. [Google Scholar] [CrossRef]
- Cox, J.; Hein, M.Y.; Luber, C.A.; Paron, I.; Nagaraj, N.; Mann, M. Accurate Proteome-wide Label-free Quantification by Delayed Normalization and Maximal Peptide Ratio Extraction, Termed MaxLFQ. Mol. Cell Proteom. 2014, 13, 2513–2526. [Google Scholar] [CrossRef] [Green Version]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]
- Krämer, A.; Green, J.; Pollard, J., Jr.; Tugendreich, S. Causal analysis approaches in Ingenuity Pathway Analysis. Bioinformatics 2014, 30, 523–530. [Google Scholar] [CrossRef] [PubMed]
- Antonsen, E.L.; Myers, J.G.; Boley, L.; Arellano, J.; Kerstman, E.; Kadwa, B.; Buckland, D.M.; Van Baalen, M. Estimating medical risk in human spaceflight. npj Microgravity 2022, 8, 8. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Astronaut (LDM) | Body Weight (kg) | Height (cm) | BMI * |
|---|---|---|---|
| SDM | 89.4 | 186.0 | 25.8 |
| A | 69.6 | 172.2 | 23.5 |
| B | 87.42 | 184.0 | 25.8 |
| C | 89.37 | 187.97 | 25.3 |
| D | 85.51 | 183.50 | 25.3 |
| Astronaut (LDM) | CEVIS (Days ISS) | T2 (Days ISS) | ARED (Days ISS) |
|---|---|---|---|
| SDM | N/A | N/A | N/A |
| A | 45 (FD4 to FD184) | 88 (FD10 to FD175) | 107 (FD8 to FD181) |
| B | 44 (FD6 to FD118 | 75 (FD8 to FD128) | 41 (FD8 to FD175) |
| C | 30 (FD7 to Fd128) | 64 (FD4 to Fd137) | 29 (FD8 to FD175) |
| D | 40 (FD6 to FD 153) | 68 (FD 5 to FD 133) | 39 (FD8 to FD 175) |
| Canonical Pathways | R+0/PRE SDM | R+0/PRE LDM | Molecules |
|---|---|---|---|
| Oxidative Phosphorylation | −4.69 | −0.333 | ATP5F1A, ATP5F1B, ATP5F1C, ATP5F1D, ATP5MF, ATP5PD, ATP5PF, COX4I1, COX5B, COX6B1, CYCS, MT-CO2, NDUFA5, NDUFB10, NDUFB8, NDUFS1, NDUFS2, NDUFS5, NDUFS8, SDHA, UQCRC1, UQCRC2 |
| TCA Cycle II (Eukaryotic) | −2.646 | −2 | ACO2, CS, FH, MDH1, OGDH, SDHA, SUCLA2, DLD, DLST |
| Necroptosis Signaling Pathway | −2 | N/A | SLC25A4, VDAC1, VDAC2, VDAC3 |
| GP6 Signaling Pathway | 2 | N/A | COL6A1, COL6A3, FGA, FGB |
| Fatty Acid β-oxidation I | N/A | −2.646 | ACAA2, ACADM, ACSL1, ECHS1, ECI2, HADH, HADHA |
| ILK Signaling | 0 | −2.449 | ACTN2, CFL2, MYH7, MYL2, MYL3, MYL6B |
| RHOA Signaling | 0 | −2 | CFL2, MYL2, MYL3, MYL6B |
| Dilated Cardiomyopathy Signaling Pathway | 0 | 2.236 | MYH7, MYL2, MYL3, MYL6B, PRKACA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blottner, D.; Moriggi, M.; Trautmann, G.; Hastermann, M.; Capitanio, D.; Torretta, E.; Block, K.; Rittweger, J.; Limper, U.; Gelfi, C.; et al. Space Omics and Tissue Response in Astronaut Skeletal Muscle after Short and Long Duration Missions. Int. J. Mol. Sci. 2023, 24, 4095. https://doi.org/10.3390/ijms24044095
Blottner D, Moriggi M, Trautmann G, Hastermann M, Capitanio D, Torretta E, Block K, Rittweger J, Limper U, Gelfi C, et al. Space Omics and Tissue Response in Astronaut Skeletal Muscle after Short and Long Duration Missions. International Journal of Molecular Sciences. 2023; 24(4):4095. https://doi.org/10.3390/ijms24044095
Chicago/Turabian StyleBlottner, Dieter, Manuela Moriggi, Gabor Trautmann, Maria Hastermann, Daniele Capitanio, Enrica Torretta, Katharina Block, Joern Rittweger, Ulrich Limper, Cecilia Gelfi, and et al. 2023. "Space Omics and Tissue Response in Astronaut Skeletal Muscle after Short and Long Duration Missions" International Journal of Molecular Sciences 24, no. 4: 4095. https://doi.org/10.3390/ijms24044095