
Glucokinase Inactivation Ameliorates Lipid Accumulation and Exerts Favorable Effects on Lipid Metabolism in Hepatocytes


Abstract
:1. Introduction
2. Results
2.1. GCK-MODY Patients Exert Favorable Lipid Profile
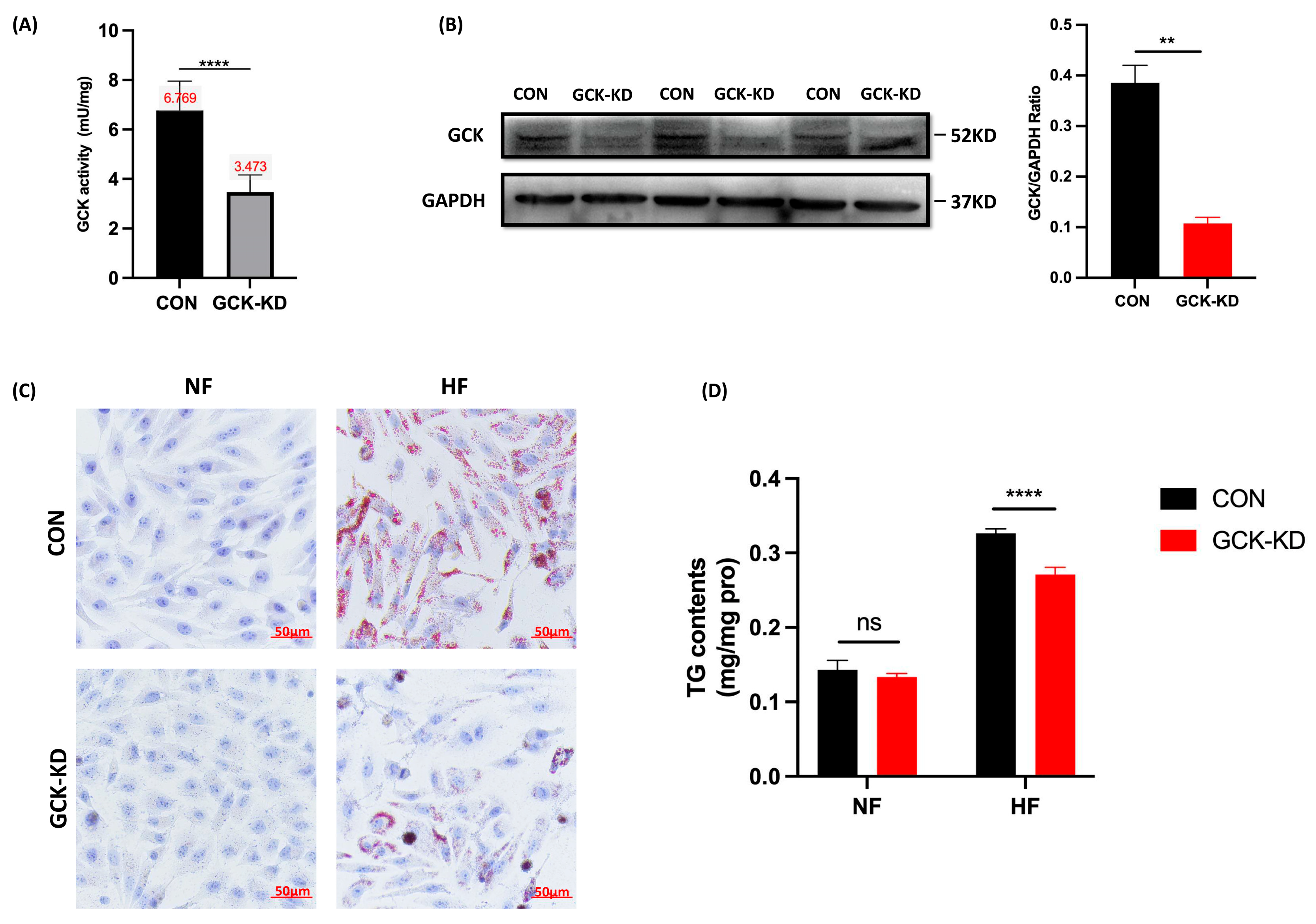
2.2. GCK Knockdown Improved Lipid Accumulation in HFA-Treated HepG2 Cells
2.3. Lipid Profile in GCK Knockdown HepG2 Cells
2.4. GCK Knockdown Altered Hepatic Lipid Metabolism
2.5. Impact of GCK Knockdown on Lipid Metabolism-Related Enzymes and Inflammatory Genes in Human and Mouse Hepatic Cell Lines
3. Discussion
4. Materials and Methods
4.1. Study Population and Data Collection
4.2. Cell Culture and High Fatty Acid Treatment
4.3. Lentivirus Transfection
4.4. SiRNA Transfection
4.5. Oil Red O Staining and Intracellular TAG Levels
4.6. Lipidomic Analysis
4.7. Data Processing
4.8. GCK Enzyme Activity Determination
4.9. Enzyme-Linked Immunosorbent Assay (ELISA)
4.10. Western Blotting
4.11. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Matschinsky, F.M.; Magnuson, M.A.; Zelent, D.; Jetton, T.L.; Doliba, N.; Han, Y.; Taub, R.; Grimsby, J. The network of glucokinase-expressing cells in glucose homeostasis and the potential of glucokinase activators for diabetes therapy. Diabetes 2006, 55, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Iynedjian, P.B. Molecular physiology of mammalian glucokinase. Cell Mol. Life Sci. 2009, 66, 27–42. [Google Scholar] [CrossRef] [PubMed]
- Bishay, R.H.; Greenfield, J.R. A review of maturity onset diabetes of the young (MODY) and challenges in the management of glucokinase-MODY. Med. J. Aust. 2016, 205, 480–485. [Google Scholar] [CrossRef] [PubMed]
- Anik, A.; Catli, G.; Abaci, A.; Bober, E. Maturity-onset diabetes of the young (MODY): An update. J. Pediatr. Endocrinol. Metab. 2015, 28, 251–263. [Google Scholar] [CrossRef]
- Steele, A.M.; Shields, B.M.; Wensley, K.J.; Colclough, K.; Ellard, S.; Hattersley, A.T. Prevalence of vascular complications among patients with glucokinase mutations and prolonged, mild hyperglycemia. JAMA 2014, 311, 279–286. [Google Scholar] [CrossRef]
- Spégel, P.; Ekholm, E.; Tuomi, T.; Groop, L.; Mulder, H.; Filipsson, K. Metabolite profiling reveals normal metabolic control in carriers of mutations in the glucokinase gene (MODY2). Diabetes 2013, 62, 653–661. [Google Scholar] [CrossRef]
- Wang, X.; Lam, S.M.; Cao, M.; Wang, T.; Wang, Z.; Yu, M.; Li, B.; Zhang, H.; Ping, F.; Song, G.; et al. Localized increases in CEPT1 and ATGL elevate plasmalogen phosphatidylcholines in HDLs contributing to atheroprotective lipid profiles in hyperglycemic GCK-MODY. Redox Biol. 2021, 40, 101855. [Google Scholar] [CrossRef]
- Eid, S.; Sas, K.M.; Abcouwer, S.F.; Feldman, E.L.; Gardner, T.W.; Pennathur, S.; Fort, P.E. New insights into the mechanisms of diabetic complications: Role of lipids and lipid metabolism. Diabetologia 2019, 62, 1539–1549. [Google Scholar] [CrossRef]
- Agius, L. Lessons from glucokinase activators: The problem of declining efficacy. Expert. Opin. Ther. Pat. 2014, 24, 1155–1159. [Google Scholar] [CrossRef]
- Meininger, G.E.; Scott, R.; Alba, M.; Shentu, Y.; Luo, E.; Amin, H.; Davies, M.J.; Kaufman, K.D.; Goldstein, B.J. Effects of MK-0941, a novel glucokinase activator, on glycemic control in insulin-treated patients with type 2 diabetes. Diabetes Care 2011, 34, 2560–2566. [Google Scholar] [CrossRef]
- Katz, L.; Manamley, N.; Snyder, W.J.; Dodds, M.; Agafonova, N.; Sierra-Johnson, J.; Cruz, M.; Kaur, P.; Mudaliar, S.; Raskin, P.; et al. AMG 151 (ARRY-403), a novel glucokinase activator, decreases fasting and postprandial glycaemia in patients with type 2 diabetes. Diabetes Obes. Metab. 2016, 18, 191–195. [Google Scholar] [CrossRef]
- De Ceuninck, F.; Kargar, C.; Ilic, C.; Caliez, A.; Rolin, J.O.; Umbdenstock, T.; Vinson, C.; Combettes, M.; de Fanti, B.; Harley, E.; et al. Small molecule glucokinase activators disturb lipid homeostasis and induce fatty liver in rodents: A warning for therapeutic applications in humans. Br. J. Pharmacol. 2013, 168, 339–353. [Google Scholar] [CrossRef]
- Kawata, S.; Nakamura, A.; Miyoshi, H.; Yang, K.; Shigesawa, I.; Yamauchi, Y.; Tsuchida, K.; Omori, K.; Takahashi, K.; Nomoto, H.; et al. Glucokinase activation leads to an unsustained hypoglycaemic effect with hepatic triglyceride accumulation in db/db mice. Diabetes Obes. Metab 2022, 24, 391–401. [Google Scholar] [CrossRef]
- Palomer, X.; Pizarro-Delgado, J.; Barroso, E.; Vázquez-Carrera, M. Palmitic and Oleic Acid: The Yin and Yang of Fatty Acids in Type 2 Diabetes Mellitus. Trends Endocrinol. Metab. 2018, 29, 178–190. [Google Scholar] [CrossRef]
- Joshi-Barve, S.; Barve, S.S.; Amancherla, K.; Gobejishvili, L.; Hill, D.; Cave, M.; Hote, P.; McClain, C.J. Palmitic acid induces production of proinflammatory cytokine interleukin-8 from hepatocytes. Hepatology 2007, 46, 823–830. [Google Scholar] [CrossRef]
- Zhao, N.; Tan, H.; Wang, L.; Han, L.; Cheng, Y.; Feng, Y.; Li, T.; Liu, X. Palmitate induces fat accumulation via repressing FoxO1-mediated ATGL-dependent lipolysis in HepG2 hepatocytes. PLoS ONE 2021, 16, e0243938. [Google Scholar] [CrossRef]
- Afshinnia, F.; Rajendiran, T.M.; Soni, T.; Byun, J.; Wernisch, S.; Sas, K.M.; Hawkins, J.; Bellovich, K.; Gipson, D.; Michailidis, G.; et al. Impaired -Oxidation and Altered Complex Lipid Fatty Acid Partitioning with Advancing CKD. J. Am. Soc. Nephrol. 2018, 29, 295–306. [Google Scholar] [CrossRef]
- Rodriguez-Cuenca, S.; Pellegrinelli, V.; Campbell, M.; Oresic, M.; Vidal-Puig, A. Sphingolipids and glycerophospholipids—The “ying and yang” of lipotoxicity in metabolic diseases. Prog. Lipid. Res. 2017, 66, 14–29. [Google Scholar] [CrossRef]
- McMaster, C.R. From yeast to humans—Roles of the Kennedy pathway for phosphatidylcholine synthesis. FEBS Lett. 2018, 592, 1256–1272. [Google Scholar] [CrossRef]
- Cole, L.K.; Vance, J.E.; Vance, D.E. Phosphatidylcholine biosynthesis and lipoprotein metabolism. Biochim. Biophys. Acta 2012, 1821, 754–761. [Google Scholar] [CrossRef]
- Maeba, R.; Kojima, K.-I.; Nagura, M.; Komori, A.; Nishimukai, M.; Okazaki, T.; Uchida, S. Association of cholesterol efflux capacity with plasmalogen levels of high-density lipoprotein: A cross-sectional study in chronic kidney disease patients. Atherosclerosis 2018, 270, 102–109. [Google Scholar] [CrossRef] [PubMed]
- Cao, P.; Wu, Y.; Li, Y.; Xiang, L.; Cheng, B.; Hu, Y.; Jiang, X.; Wang, Z.; Wu, S.; Si, L.; et al. The important role of glycerophospholipid metabolism in the protective effects of polyphenol-enriched Tartary buckwheat extract against alcoholic liver disease. Food Funct. 2022, 13, 10415–10425. [Google Scholar] [CrossRef] [PubMed]
- Piras, I.S.; Raju, A.; Don, J.; Schork, N.J.; Gerhard, G.S.; DiStefano, J.K. Hepatic PEMT Expression Decreases with Increasing NAFLD Severity. Int. J. Mol. Sci. 2022, 23, 9296. [Google Scholar] [CrossRef] [PubMed]
- Luukkonen, P.K.; Zhou, Y.; Sädevirta, S.; Leivonen, M.; Arola, J.; Orešič, M.; Hyötyläinen, T.; Yki-Järvinen, H. Hepatic ceramides dissociate steatosis and insulin resistance in patients with non-alcoholic fatty liver disease. J. Hepatol. 2016, 64, 1167–1175. [Google Scholar] [CrossRef]
- Puri, P.; Baillie, R.A.; Wiest, M.M.; Mirshahi, F.; Choudhury, J.; Cheung, O.; Sargeant, C.; Contos, M.J.; Sanyal, A.J. A lipidomic analysis of nonalcoholic fatty liver disease. Hepatology 2007, 46, 1081–1090. [Google Scholar] [CrossRef]
- Summers, S.A. Editorial: The Role of Ceramides in Diabetes and Cardiovascular Disease. Front. Endocrinol. 2021, 12, 667885. [Google Scholar] [CrossRef]
- Ou-Yang, Q.; Lin, X.-M.; Zhu, Y.-J.; Zheng, B.; Li, L.; Yang, Y.-C.; Hou, G.-J.; Chen, X.; Luo, G.-J.; Huo, F.; et al. Distinct role of nuclear receptor corepressor 1 regulated de novo fatty acids synthesis in liver regeneration and hepatocarcinogenesis in mice. Hepatology 2018, 67, 1071–1087. [Google Scholar] [CrossRef]
- Pang, Y.; Xu, X.; Xiang, X.; Li, Y.; Zhao, Z.; Li, J.; Gao, S.; Liu, Q.; Mai, K.; Ai, Q. High Fat Activates O-GlcNAcylation and Affects AMPK/ACC Pathway to Regulate Lipid Metabolism. Nutrients 2021, 13, 1740. [Google Scholar] [CrossRef]
- Naiman, S.; Huynh, F.K.; Gil, R.; Glick, Y.; Shahar, Y.; Touitou, N.; Nahum, L.; Avivi, M.Y.; Roichman, A.; Kanfi, Y.; et al. SIRT6 Promotes Hepatic Beta-Oxidation via Activation of PPARα. Cell Rep. 2019, 29, 4127–4143e8. [Google Scholar] [CrossRef]
- Xu, Y.-H.; Luo, Z.; Wu, K.; Fan, Y.-F.; You, W.-J.; Zhang, L.-H. Structure and Functional Analysis of Promoters from Two Liver Isoforms of CPT I in Grass Carp Ctenopharyngodon idella. Int. J. Mol. Sci. 2017, 18, 2405. [Google Scholar] [CrossRef]
- Kim, S.-J.; Tang, T.; Abbott, M.; Viscarra, J.A.; Wang, Y.; Sul, H.S. AMPK Phosphorylates Desnutrin/ATGL and Hormone-Sensitive Lipase To Regulate Lipolysis and Fatty Acid Oxidation within Adipose Tissue. Mol Cell Biol. 2016, 36, 1961–1976. [Google Scholar] [CrossRef]
- McMaster, C.R.; Bell, R.M. CDP-choline:1,2-diacylglycerol cholinephosphotransferase. Biochim. Biophys. Acta 1997, 1348, 100–110. [Google Scholar] [CrossRef]
- Kawana, M.; Miyamoto, M.; Ohno, Y.; Kihara, A. Comparative profiling and comprehensive quantification of stratum corneum ceramides in humans and mice by LC/MS/MS. J. Lipid. Res. 2020, 61, 884–895. [Google Scholar] [CrossRef]
- Osbak, K.K.; Colclough, K.; Saint-Martin, C.; Beer, N.L.; Bellanné-Chantelot, C.; Ellard, S.; Gloyn, A.L. Update on mutations in glucokinase (GCK), which cause maturity-onset diabetes of the young, permanent neonatal diabetes, and hyperinsulinemic hypoglycemia. Hum. Mutat. 2009, 30, 1512–1526. [Google Scholar] [CrossRef]
- Dunn, W.B.; Broadhurst, D.; Begley, P.; Zelena, E.; Francis-McIntyre, S.; Anderson, N.; Brown, M.; Knowles, J.D.; Halsall, A.; Haselden, J.N.; et al. Procedures for large-scale metabolic profiling of serum and plasma using gas chromatography and liquid chromatography coupled to mass spectrometry. Nat. Protoc. 2011, 6, 1060–1083. [Google Scholar] [CrossRef]
- Want, E.J.; Wilson, I.D.; Gika, H.; Theodoridis, G.; Plumb, R.S.; Shockcor, J.; Holmes, E.; Nicholson, J.K. Global metabolic profiling procedures for urine using UPLC-MS. Nat. Protoc. 2010, 5, 1005–1018. [Google Scholar] [CrossRef]
- Smith, C.A.; Want, E.J.; O’Maille, G.; Abagyan, R.; Siuzdak, G. XCMS: Processing mass spectrometry data for metabolite profiling using nonlinear peak alignment, matching, and identification. Anal. Chem. 2006, 78, 779–787. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | MODY2 (n = 33) | T1DM (n = 34) | T2DM (n = 34) | Control (n = 30) | p |
|---|---|---|---|---|---|
| Gender (M/F) | 12/21 | 14/20 | 13/21 | 14/16 | 0.853 |
| Age (Year) | 25.4 ± 16.42 | 18.6 ± 9.54 | 24.7 ± 7.91 | 16.8 ± 10.14 & | <0.01 |
| BMI (kg/m2) | 19.1 ± 4.44 | 19.0 ± 4.97 | 25.7 ± 4.53 * | 19.6 ± 3.11 | <0.0001 |
| FBG (mmol/L) | 6.7 (6.4, 7.1) | 7.8 (6.4, 10.5) # | 7.6 (6.4, 9.9) * | 4.7 (4.5, 5.1) & | <0.0001 |
| Fasting insulin (mIU/L) | 5.4 (3.9, 8.9) | 16.4 (10.6, 24.2) # | 13.7 (8.1, 21.8) | 7.9 (4.9, 10.2) | 0.0019 |
| Fasting C-peptide (ng/mL) | 0.89 (0.72, 1.36) | 0.37 (0.18, 0.52) # | 1.36 (0.85, 2.02) * | 1.13 (0.71, 1.25) | <0.0001 |
| HbA1c (%) | 6.3 ± 0.37 | 8.7 ± 2.29 # | 8.4 ± 2.3 * | 4.7 ± 0.4 & | <0.001 |
| GA (%) | 18.0 ± 1.46 | 23.5 ± 6.39 # | 22.3 ± 6.37 * | 13.6 ± 1.19 & | <0.0001 |
| TAG (mmol/L) | 0.47 (0.39, 0.86) | 0.64 (0.46, 0.81) | 1.59 (1.36, 1.8) * | 0.73 (0.48, 1.02) | <0.0001 |
| TC (mmol/L) | 4.4 ± 0.77 | 4.5 ± 0.86 | 5.3 ± 1.05 * | 4.0 ± 0.67 | <0.0001 |
| LDL-C (mmol/L) | 2.1 ± 0.57 | 2.6 ± 0.76 | 3.3 ± 1.01 * | 2.2 ± 0.68 | <0.0001 |
| HDL-C (mmol/L) | 1.6 ± 0.25 | 1.3 ± 0.23 # | 1.2 ± 0.42 * | 1.5 ± 0.21 | <0.0001 |
| ALT (mmol/L) | 14 (11, 18) | 14 (10, 18.5) | 21 (16, 33) * | 11 (9, 15.5) | <0.0001 |
| Cr (umol/L) | 55.9 ± 18.10 | 52.6 ± 16.64 | 60.8 ± 26.3 | 53.6 ± 11.32 | 0.3371 |
| UA (umol/L) | 258.7 ± 73.32 | 255.6 ± 64.20 | 362.3 ± 109. 9 * | 307.5 ± 93.07 | <0.0001 |
| CRP (mg/L) | 0.19 (0.16, 0.36) | 0.66 (0.28, 1.58) # | 1.3 (0.49, 1.95) * | 0.26 (0.17, 0.44) | 0.0065 |
| Hcy (umol/L) | 9.3 (8.5, 11.1) | 9.7 (8, 10.3) | 16.1 (10.1, 17.1) | 11 (9.5, 13.8) | 0.0087 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, Z.; Xie, T.; Liu, J.; Zhang, Q.; Xiao, X. Glucokinase Inactivation Ameliorates Lipid Accumulation and Exerts Favorable Effects on Lipid Metabolism in Hepatocytes. Int. J. Mol. Sci. 2023, 24, 4315. https://doi.org/10.3390/ijms24054315
Xie Z, Xie T, Liu J, Zhang Q, Xiao X. Glucokinase Inactivation Ameliorates Lipid Accumulation and Exerts Favorable Effects on Lipid Metabolism in Hepatocytes. International Journal of Molecular Sciences. 2023; 24(5):4315. https://doi.org/10.3390/ijms24054315
Chicago/Turabian StyleXie, Ziyan, Ting Xie, Jieying Liu, Qian Zhang, and Xinhua Xiao. 2023. "Glucokinase Inactivation Ameliorates Lipid Accumulation and Exerts Favorable Effects on Lipid Metabolism in Hepatocytes" International Journal of Molecular Sciences 24, no. 5: 4315. https://doi.org/10.3390/ijms24054315
APA StyleXie, Z., Xie, T., Liu, J., Zhang, Q., & Xiao, X. (2023). Glucokinase Inactivation Ameliorates Lipid Accumulation and Exerts Favorable Effects on Lipid Metabolism in Hepatocytes. International Journal of Molecular Sciences, 24(5), 4315. https://doi.org/10.3390/ijms24054315