
Structural and Serological Characterization of the O Antigen of Proteus mirabilis Clinical Isolates Classified into a New Proteus Serogroup, O84

Abstract
1. Introduction
2. Results and Discussion
2.1. Characterization of Strains
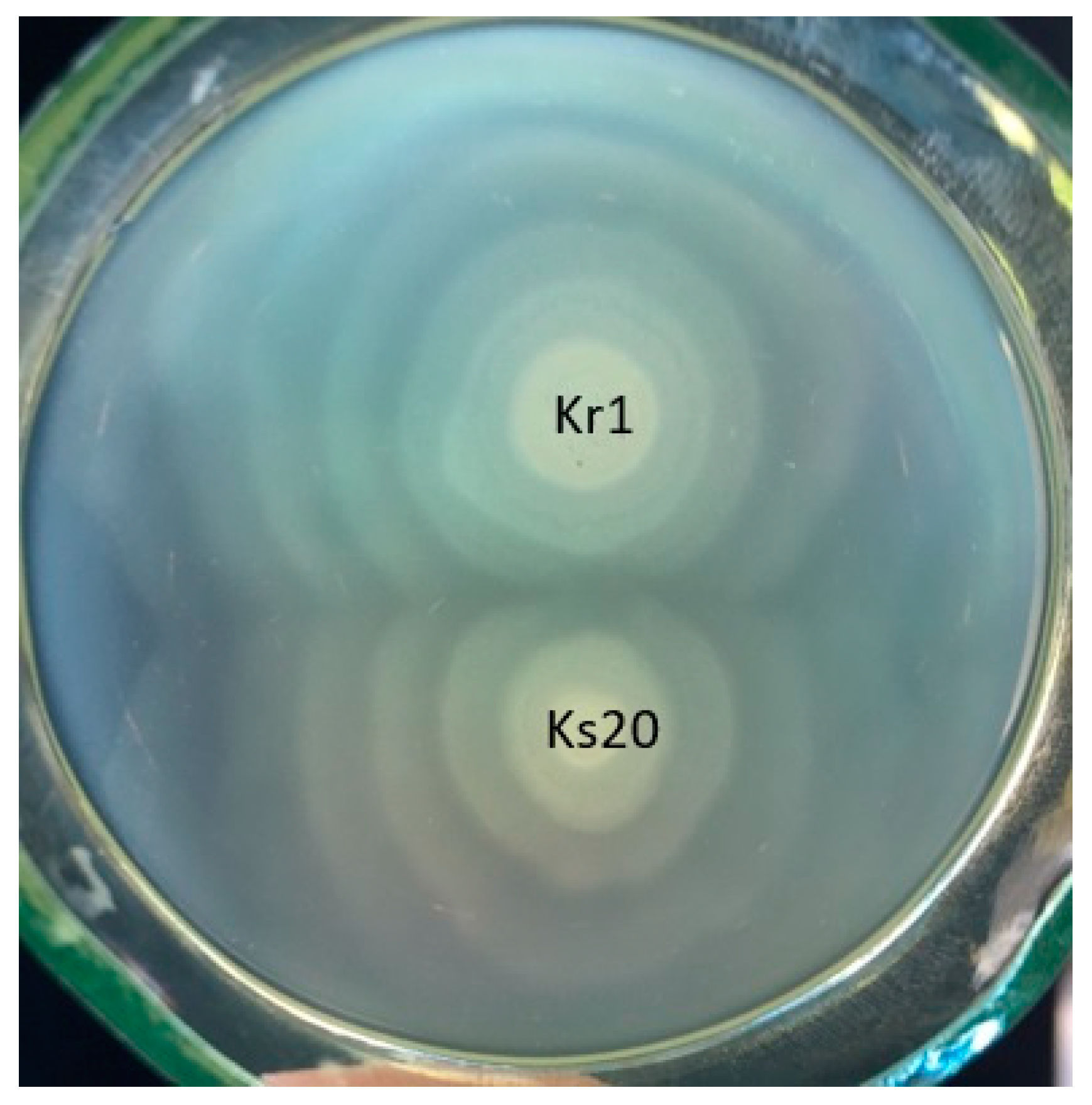
2.2. Serological Studies
2.3. Structural Studies
2.4. Classification of P. mirabilis Kr1 and Ks20
3. Materials and Methods
3.1. Bacterial Strains, Identification, Characterization, Cultivation
3.2. Lipopolysaccharides (LPSs), Antisera
3.3. Serological Assays
3.4. Isolation of O-Specific Polysaccharide Samples
3.5. O-Deacetylation
3.6. Composition Analysis
3.7. Nuclear Magnetic Resonance (NMR) Spectroscopy
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- O’Hara, C.M.; Brenner, F.W.; Miller, J.M. Classification, identification, and clinical significance of Proteus, Providencia, and Morganella. Clin. Microbiol. Rev. 2000, 13, 534–546. [Google Scholar] [CrossRef] [PubMed]
- Różalski, A.; Torzewska, A.; Moryl, M.; Kwil, I.; Maszewska, A.; Ostrowska, K.; Drzewiecka, D.; Zabłotni, A.; Palusiak, A.; Siwińska, M.; et al. Proteus sp.—An opportunistic bacterial pathogen—classification, swarming growth, clinical significance and virulence factors. Acta Univ. Lodziensis. Folia Biol. Oecologica 2012, 8, 1–17. [Google Scholar] [CrossRef]
- Adeolu, M.; Alnajar, S.; Naushad, S.; Gupta, R.S. Genome-based phylogeny and taxonomy of the ‘Enterobacteriales’: Proposal for Enterobacterales ord. nov. divided into the families Enterobacteriaceae, Erwiniaceae fam. nov., Pectobacteriaceae fam. nov., Yersiniaceae fam. nov., Hafniaceae fam. nov., Morganellaceae fam. nov., and Budviciaceae fam. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5575–5599. [Google Scholar] [PubMed]
- Dai, H.; Lu, B.; Li, Z.; Huang, Z.; Cai, H.; Yu, K.; Wang, D. Multilocus sequence analysis for the taxonomic updating and identification of the genus Proteus and reclassification of Proteus genospecies 5 O’Hara et al. 2000, Proteus cibarius Hyun et al. 2016 as later heterotypic synonyms of Proteus terrae Behrendt et al. 2015. BMC Microbiol. 2020, 20, 152. [Google Scholar]
- Drzewiecka, D. Significance and roles of Proteus bacteria in natural environments. Microb. Ecol. 2016, 72, 741–758. [Google Scholar] [CrossRef]
- Pal, N.; Sharma, N.; Sharma, R.; Hooja, S.; Maheshwari, R.K. Prevalence of multidrug (MDR) and extensively drug resistant (XDR) Proteus species in a tertiary care hospital. India Int. J. Curr. Microbiol. App. Sci. 2014, 3, 243–252. [Google Scholar]
- Reśliński, A.; Gospodarek, E.; Mikucka, A. Prevalence of multidrug-resistant Proteus spp. strains in clinical specimens and their susceptibility to antibiotics. Med. Dośw. Mikrobiol. 2005, 57, 175–184. [Google Scholar]
- Armbruster, C.E.; Mobley, H.L.T.; Pearson, M.M. Pathogenesis of Proteus mirabilis infection. EcoSal. Plus 2018, 8, 1–123. [Google Scholar] [CrossRef]
- Yuan, F.; Huang, Z.; Yang, T.; Wang, G.; Li, P.; Yang, B.; Li, J. Pathogenesis of Proteus mirabilis in catheter-associated urinary tract infections. Urol. Int. 2021, 105, 354–361. [Google Scholar] [CrossRef]
- Jacobsen, S.M.; Shirtliff, M.E. Proteus mirabilis biofilms and catheter-associated urinary tract infections. Virulence 2011, 2, 460–465. [Google Scholar] [CrossRef]
- Filipiak, A.; Chrapek, M.; Literacka, E.; Wawszczak, M.; Głuszek, S.; Majchrzak, M.; Wróbel, G.; Łysek-Gładysińska, M.; Gniadkowski, M.; Adamus-Białek, W. Pathogenic factors correlate with antimicrobial resistance among clinical Proteus mirabilis strains. Front. Microbiol. 2020, 11, 579389. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Zhang, Y.; Zhang, X.; Li, J. An evaluation of multidrug-resistant (MDR) bacteria in patients with urinary stone disease: Data from a high-volume stone management center. World J. Urol. 2020, 38, 425–432. [Google Scholar] [CrossRef] [PubMed]
- Bessa, L.J.; Fazii, P.; Di Giulio, M.; Cellini, L. Bacterial isolates from infected wounds and their antibiotic susceptibility pattern: Some remarks about wound infection. Int. Wound J. 2015, 12, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Godebo, G.; Kibru, G.; Tassew, H. Multidrug-resistant bacterial isolates in infected wounds at Jimma University Specialized Hospital, Ethiopia. Ann. Clin. Microbiol. Antimicrob. 2013, 12, 17. [Google Scholar] [CrossRef]
- Fadeyibi, I.O.; Raji, M.A.; Ibrahim, N.A.; Ugburo, A.O.; Ademiluyi, S. Bacteriology of infected burn wounds in the burn wards of a teaching hospital in Southwest Nigeria. Burns 2013, 39, 168–173. [Google Scholar] [CrossRef]
- Drzewiecka, D.; Palusiak, A.; Siwińska, M.; Zabłotni, A. The prevailing O serogroups among the serologically differentiated clinical Proteus spp. strains in central Poland. Sci. Rep. 2021, 11, 18982. [Google Scholar] [CrossRef]
- Manos, J.; Belas, R. The Genera Proteus Providencia, and Morganella. In The Procaryotes; Dworkin, M., Falkow, S., Rosenberg, E., Schleifer, K.H., Stackebrandt, E., Eds.; Springer: New York, NY, USA, 2006; Volume 6, pp. 245–269. [Google Scholar]
- Armbruster, C.E.; Mobley, H.L. Merging mythology and morphology: The multifaceted lifestyle of Proteus mirabilis. Nat. Rev. Microbiol. 2012, 10, 743–754. [Google Scholar] [CrossRef]
- Little, K.; Austerman, J.; Zheng, J.; Gibbs, K.A. Cell shape and population migration are distinct steps of Proteus mirabilis swarming that are decoupled on high-percentage agar. J. Bacteriol. 2019, 201, e00726-18. [Google Scholar] [CrossRef]
- Schaffer, J.N.; Pearson, M.M. Proteus mirabilis and urinary tract infections. Microbiol. Spectr. 2015, 3, 1–66. [Google Scholar] [CrossRef]
- Palusiak, A. Proteus mirabilis and Klebsiella pneumoniae as pathogens capable of causing co-infections and exhibiting similarities in their virulence factors. Front. Cell. Infect. Microbiol. 2022, 20, 991657. [Google Scholar] [CrossRef]
- Knirel, Y.A.; Perepelov, A.V.; Kondakova, A.N.; Senchenkova, S.N.; Sidorczyk, Z.; Różalski, A.; Kaca, W. Structure and serology of O-antigens as the basis for classification of Proteus strains. Innate Immun. 2011, 17, 70–96. [Google Scholar] [CrossRef]
- Siwińska, M.; Zabłotni, A.; Levina, E.A.; Shashkov, A.S.; Ovchinnikova, O.G.; Różalski, A.; Knirel, Y.A. The unique structure of bacterial polysaccharides. Immunochemical studies on the O-antigen of Proteus penneri 4034-85 clinical strain classified into a new O83 Proteus serogroup. Int. J. Biol. Macromol. 2020, 163, 1168–1174. [Google Scholar] [CrossRef] [PubMed]
- Larsson, P. Serology of Proteus mirabilis and Proteus vulgaris. Methods Microbiol. 1984, 14, 187–214. [Google Scholar]
- Senior, B.W. Media and tests to simplify the recognition and identification of members of the Proteeae. J. Med. Microbiol. 1997, 46, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Pfaller, M.A.; Mujeeb, I.; Hollis, R.J.; Jones, R.N.; Doern, G.V. Evaluation of the discriminatory powers of the Dienes test and ribotyping as typing methods for Proteus mirabilis. J. Clin. Microbiol. 2000, 38, 1077–1080. [Google Scholar] [CrossRef] [PubMed]
- Gmiter, D.; Kaca, W. Into the understanding the multicellular lifestyle of Proteus mirabilis on solid surfaces. Front. Cell. Infect. Microbiol. 2022, 12, 864305. [Google Scholar] [CrossRef]
- Budding, A.E.; Ingham, C.J.; Bitter, W.; Vandenbroucke-Grauls, C.M.; Schneeberger, P.M. The Dienes phenomenon: Competition and territoriality in swarming Proteus mirabilis. J. Bacteriol. 2009, 191, 3892–3900. [Google Scholar] [CrossRef]
- Drzewiecka, D.; Arbatsky, N.P.; Shashkov, A.S.; Stączek, P.; Knirel, Y.A.; Sidorczyk, Z. Structure and serological properties of the O-antigen of two clinical Proteus mirabilis strains classified into a new Proteus O77 serogroup. FEMS Immunol. Med. Microbiol. 2008, 54, 185–194. [Google Scholar] [CrossRef]
- Drzewiecka, D.; Arbatsky, N.P.; Stączek, P.; Shashkov, A.S.; Knirel, Y.A.; Sidorczyk, Z. Structural and serological studies of the O-polysaccharide of strains from a newly created Proteus O78 serogroup prevalent in Polish patients. FEMS Immunol. Med. Microbiol. 2010, 58, 269–276. [Google Scholar] [CrossRef]
- Munson, E.L.; Pfaller, M.A.; Doern, G.V. Modification of Dienes mutual inhibition test for epidemiological characterization of Pseudomonas aeruginosa isolates. J. Clin. Microbiol. 2002, 40, 4285–4288. [Google Scholar] [CrossRef]
- Nagano, N.; Shibata, N.; Saitou, Y.; Nagano, Y.; Arakawa, Y. Nosocomial outbreak of infections by Proteus mirabilis that produces extended-spectrum CTX-M-2 type beta-lactamase. J. Clin. Microbiol. 2003, 41, 5530–5536. [Google Scholar] [CrossRef]
- Jamil, R.T.; Foris, L.A.; Snowden, J. Proteus mirabilis Infections. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar] [PubMed]
- Westphal, O.; Jann, K. Bacterial lipopolysaccharides. Extraction with phenol-water and further applications of the procedure. Methods Carbohydr. Chem. 1965, 5, 83–89. [Google Scholar]
- Sawardeker, J.S.; Sloneker, J.H.; Jeanes, A. Quantitative determination of monosaccharides as their alditol acetates by gas liquid chromatography. Anal. Chem. 1965, 37, 1602–1603. [Google Scholar] [CrossRef]
- Lipkind, G.M.; Shashkov, A.S.; Knirel, Y.A.; Vinogradov, E.V.; Kochetkov, N.K. A computer-assisted structural analysis of regular polysaccharides on the basis of 13C-n.m.r. data. Carbohydr. Res. 1988, 175, 59–75. [Google Scholar] [CrossRef] [PubMed]
- Jansson, P.-E.; Kenne, L.; Widmalm, G. Computer-assisted structural analysis of polysaccharides with an extended version of CASPER using 1H- and 13C-n.m.r. data. Carbohydr. Res. 1989, 188, 169–191. [Google Scholar] [CrossRef]
- Siwińska, M.; Levina, E.A.; Ovchinnikova, O.G.; Drzewiecka, D.; Shashkov, A.S.; Różalski, A.; Knirel, Y.A. Classification of a Proteus penneri clinical isolate with a unique O-antigen structure to a new Proteus serogroup, O80. Carbohydr Res. 2015, 407, 131–136. [Google Scholar] [CrossRef]
- Zabłotni, A.; Arbatsky, N.P.; Drzewiecka, D.; Shashkov, A.S.; Knirel, Y.A. Chemical characterization and serological properties of a unique O-polysaccharide of the Proteus mirabilis Sm 99 clinical strain. Identification of a new, O81, serotype. Int. J. Biol. Macromol. 2018, 118, 1131–1135. [Google Scholar] [CrossRef]
- Siwińska, M.; Levina, E.A.; Shashkov, A.S.; Kalinchuk, N.A.; Drzewiecka, D.; Knirel, Y.A. Structural and serological characterization of the O82 antigen of a Proteus mirabilis strain isolated from a patient in Poland. Carbohydr. Res. 2019, 486, 107831. [Google Scholar] [CrossRef]
- Yu, X.; Torzewska, A.; Zhang, X.; Yin, Z.; Drzewiecka, D.; Cao, H.; Liu, B.; Knirel, Y.A.; Różalski, A.; Wang, L. Genetic diversity of the O antigens of Proteus species and the development of a suspension array for molecular serotyping. PLoS ONE 2017, 12, e0183267. [Google Scholar] [CrossRef]
- Lüderitz, O.; Westphal, O.; Staub, A.M.; Nikaido, H. Isolation and chemical and immunological characterization of bacterial lipopolysaccharides. In Microbial Toxins; Weinbaum, G., Kadis, S., Ajl, S.J., Eds.; Academic Press Inc.: New York, NY, USA; London, UK, 1971; Volume 4, pp. 145–233. [Google Scholar]
- Arbatsky, N.P.; Shashkov, A.S.; Widmalm, G.; Knirel, Y.A.; Zych, K.; Sidorczyk, Z. Structure of the O-specific polysaccharide of Proteus penneri strain 25 containing N-(l-alanyl) and multiple O-acetyl groups in a tetrasaccharide repeating unit. Carbohydr. Res. 1997, 298, 229–235. [Google Scholar] [CrossRef]
- Sidorczyk, Z.; Zych, K.; Toukach, F.V.; Arbatsky, N.P.; Zabłotni, A.; Shashkov, A.S.; Knirel, Y.A. Structure of the O-polysaccharide and classification of Proteus mirabilis G1 in Proteus serogroup O3. Eur. J. Biochem. 2002, 269, 1406–1412. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
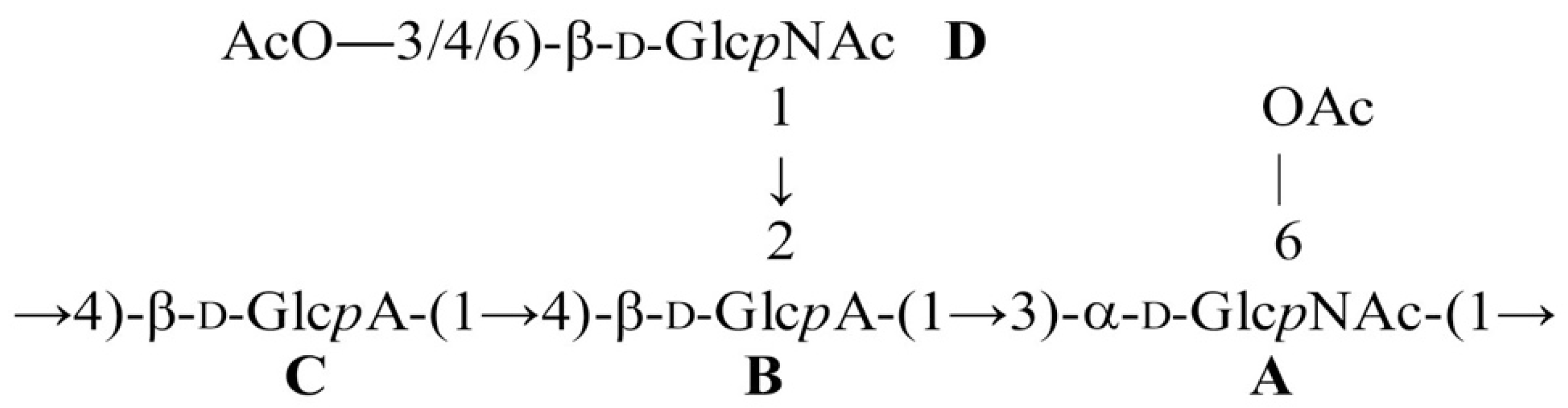{kind=link}
| P. mirabilis Kr1 O Antiserum | Reciprocal Titer in the Reaction with | |
|---|---|---|
| Kr1 LPS | Ks20 Biomass | |
| not adsorbed | 128,000 | 256,000 |
| adsorbed with P. mirabilis Ks20 biomass | <1000 | <1000 |
| Monosaccharide Residue | C-1 H-1 | C-2 H-2 | C-3 H-3 | C-4 H-4 | C-5 H-5 | C-6 H-6 (6a, 6b) |
|---|---|---|---|---|---|---|
| →3)-α-d-GlcpNAc-(1→ A | 98.5 | 53.2 | 81.5 | 69.4 | 73.3 | 61.6 |
| 5.37 | 4.02 | 3.87 | 3.58 | 3.68 | 3.78, 3.78 | |
| →2,4)-β-d-GlcpA-(1→ B | 101.3 | 80.8 | 76.7 | 82.0 | 75.5 | 174.5 a |
| 4.66 | 3.56 | 3.67 | 3.74 | 3.94 | ||
| →4)-β-d-GlcpA-(1→ C | 103.8 | 74.7 | 77.4 | 77.6 | 77.0 | 174.4 a |
| 4.52 | 3.34 | 3.71 | 3.79 | 3.93 | ||
| β-d-GlcpNAc-(1→ D | 102.9 | 57.0 | 75.4 | 71.2 | 77.2 | 62.3 |
| 4.78 | 3.70 | 3.52 | 3.42 | 3.37 | 3.71, 3.89 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Drzewiecka, D.; Siwińska, M.; Senchenkova, S.N.; Levina, E.A.; Shashkov, A.S.; Knirel, Y.A. Structural and Serological Characterization of the O Antigen of Proteus mirabilis Clinical Isolates Classified into a New Proteus Serogroup, O84. Int. J. Mol. Sci. 2023, 24, 4699. https://doi.org/10.3390/ijms24054699
Drzewiecka D, Siwińska M, Senchenkova SN, Levina EA, Shashkov AS, Knirel YA. Structural and Serological Characterization of the O Antigen of Proteus mirabilis Clinical Isolates Classified into a New Proteus Serogroup, O84. International Journal of Molecular Sciences. 2023; 24(5):4699. https://doi.org/10.3390/ijms24054699
Chicago/Turabian StyleDrzewiecka, Dominika, Małgorzata Siwińska, Sof’ya N. Senchenkova, Evgeniya A. Levina, Alexander S. Shashkov, and Yuriy A. Knirel. 2023. "Structural and Serological Characterization of the O Antigen of Proteus mirabilis Clinical Isolates Classified into a New Proteus Serogroup, O84" International Journal of Molecular Sciences 24, no. 5: 4699. https://doi.org/10.3390/ijms24054699
APA StyleDrzewiecka, D., Siwińska, M., Senchenkova, S. N., Levina, E. A., Shashkov, A. S., & Knirel, Y. A. (2023). Structural and Serological Characterization of the O Antigen of Proteus mirabilis Clinical Isolates Classified into a New Proteus Serogroup, O84. International Journal of Molecular Sciences, 24(5), 4699. https://doi.org/10.3390/ijms24054699