
Comparative Analysis of Phenotypic and Molecular Data on Response to Main Pear Diseases and Pest Attack in a Germplasm Collection


 ,
,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Results
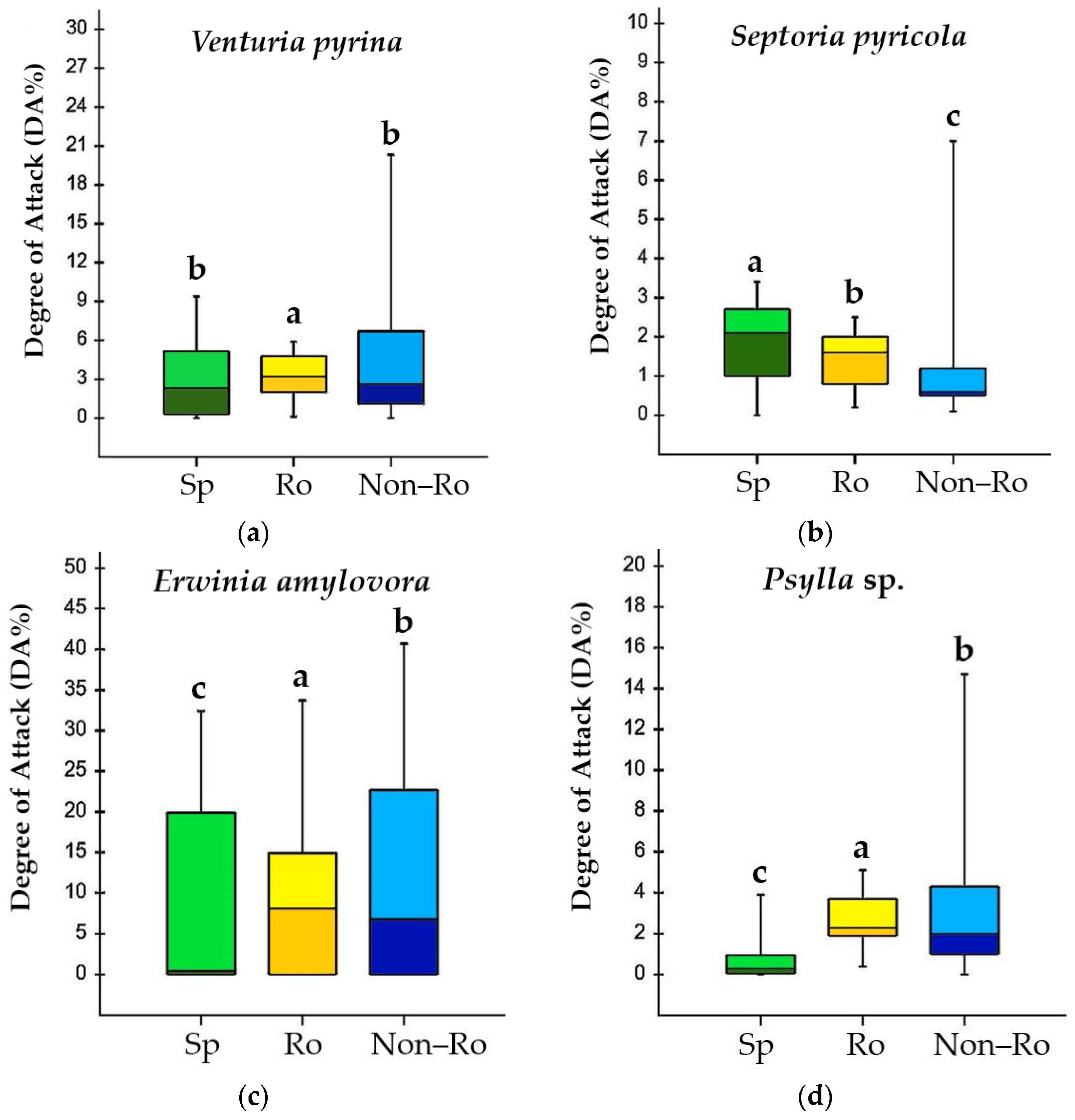
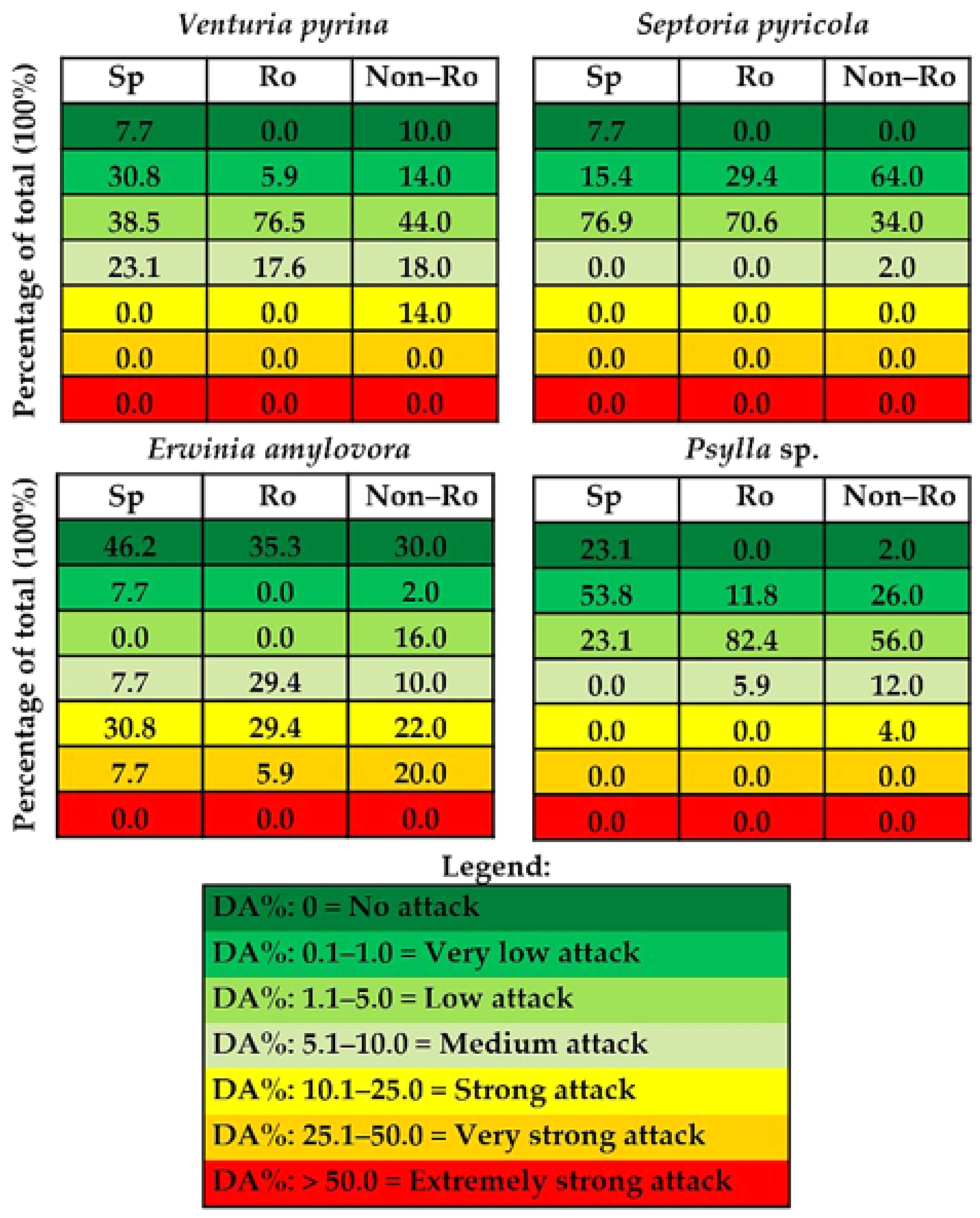
2.1. Field Evaluation
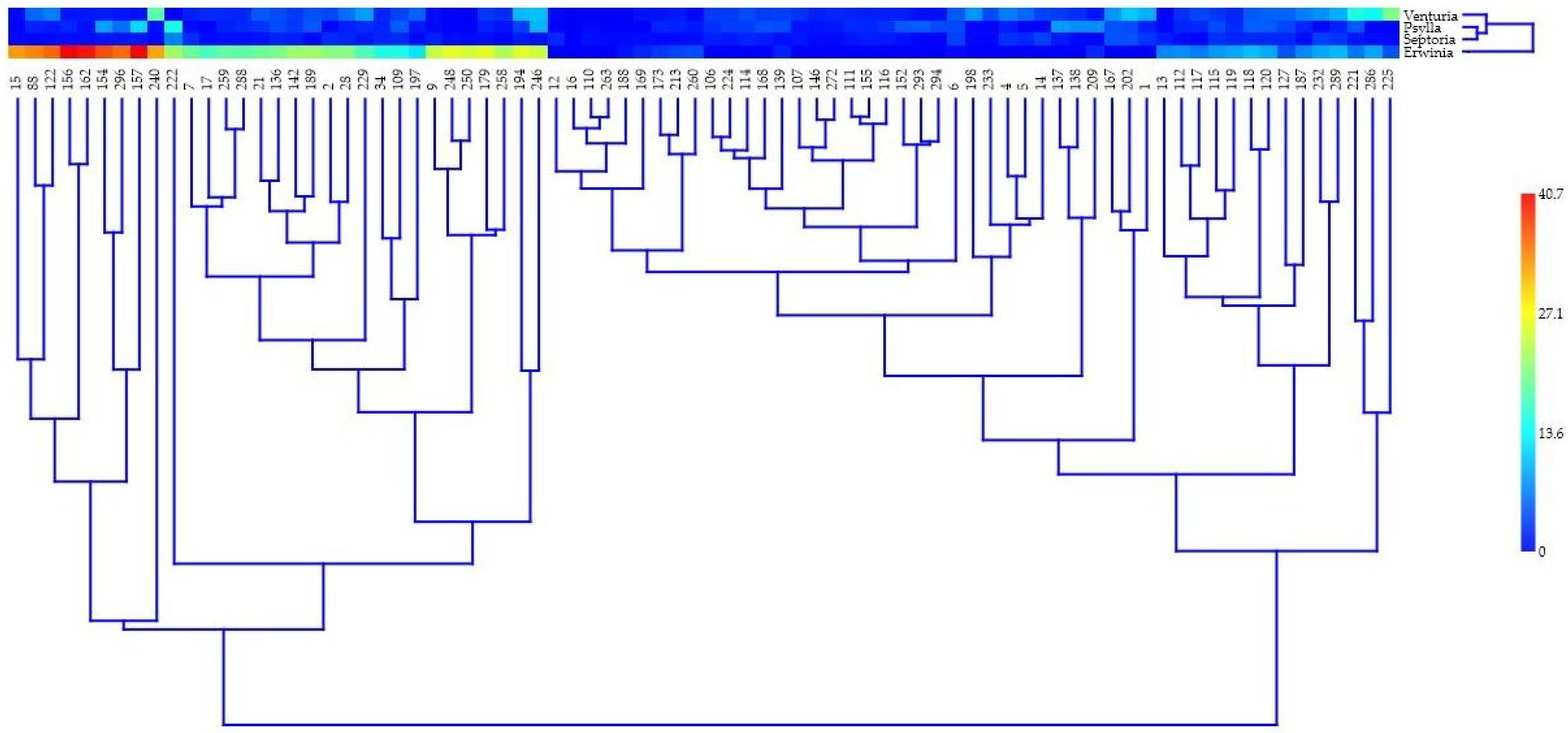
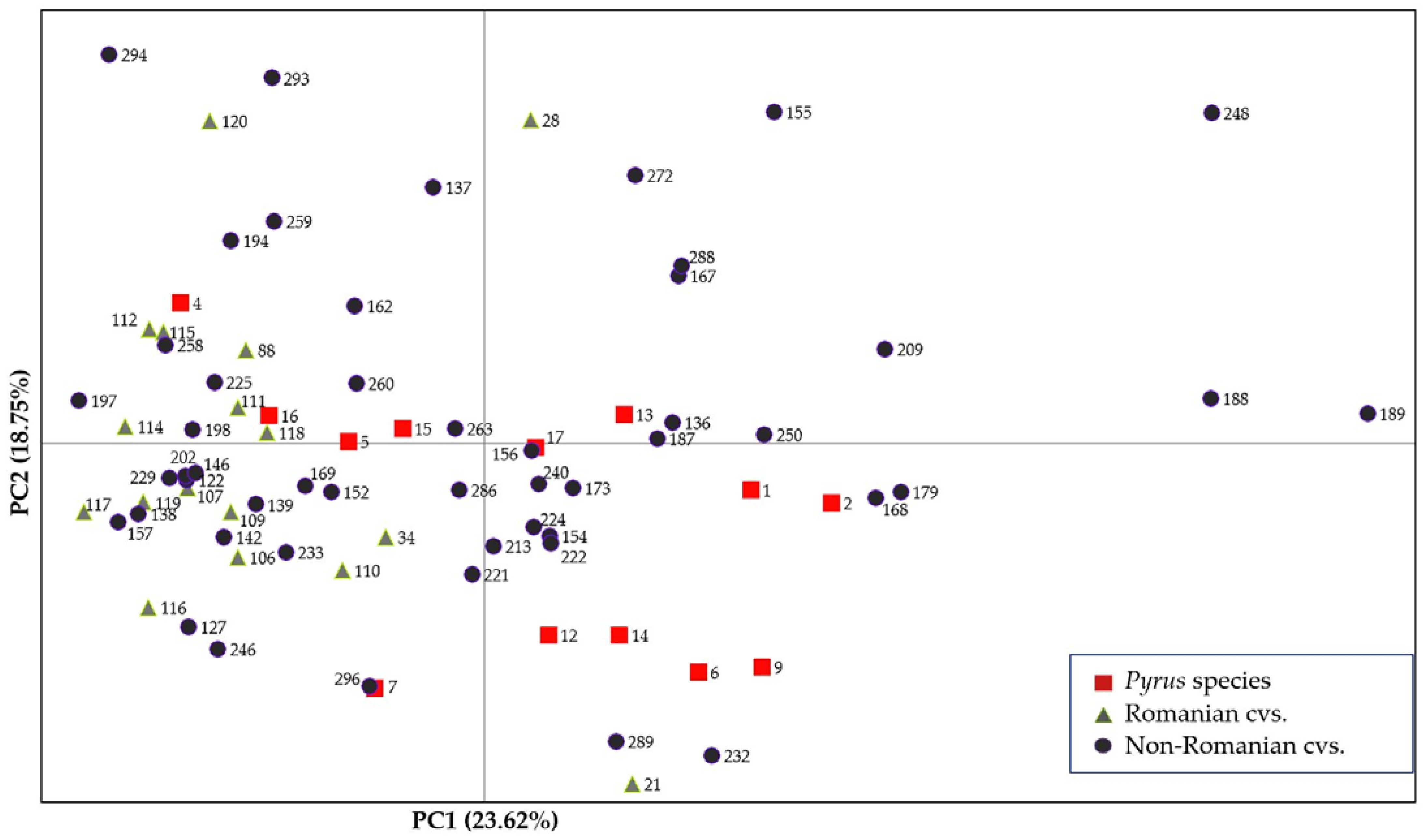
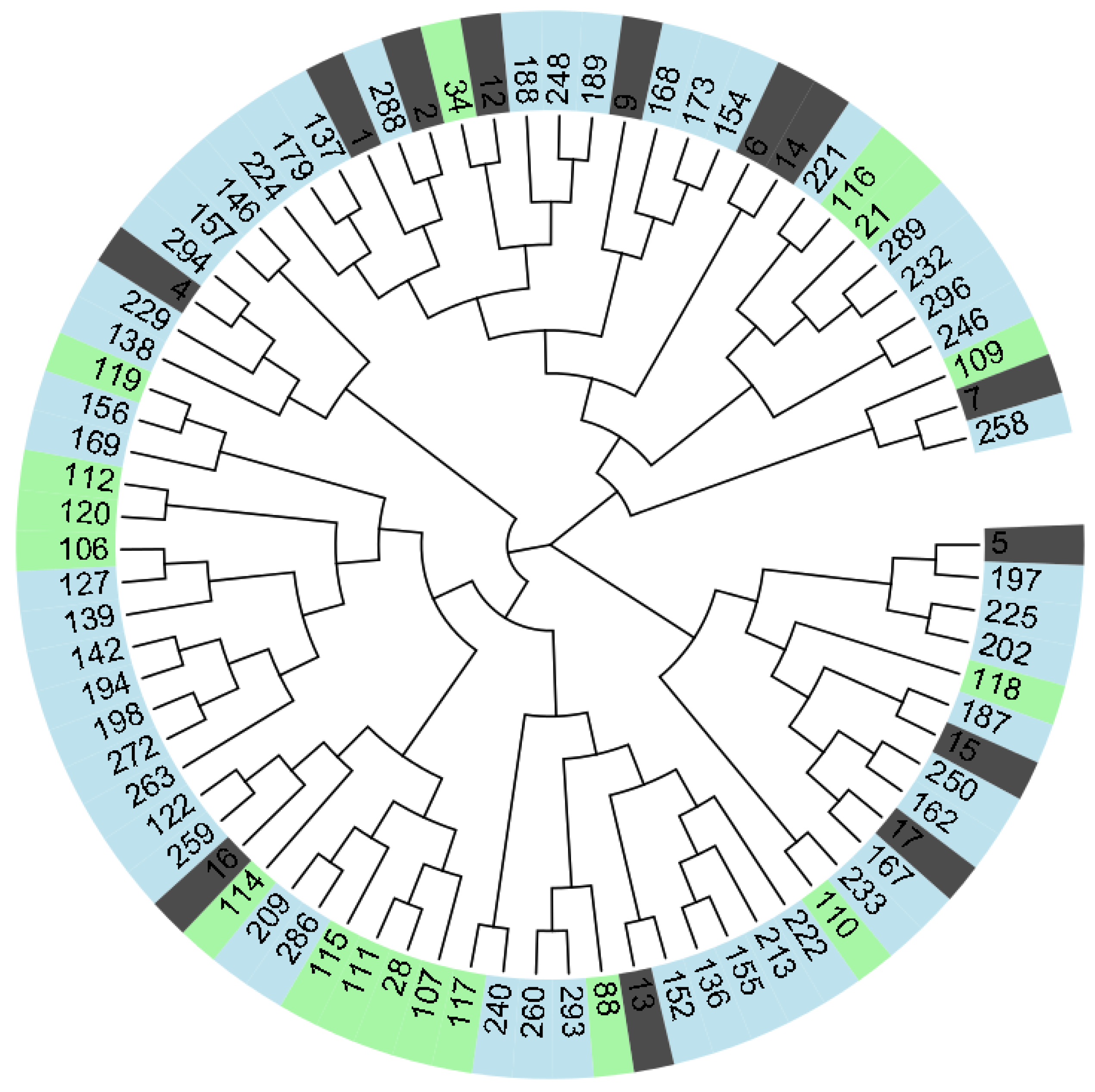
2.2. Genetic Analysis
3. Discussion
4. Materials and Methods
4.1. Description of the Study Site and Biological Material
4.2. Assessment of Diseases and Pests
4.3. Genetic Diversity Analysis
4.4. Statistical Analysis
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Filipović, D.; Obradović, Đ.; Tripković, B. Plant storage in Neolithic southeast Europe: Synthesis of the archaeological and archaeobotanical evidence from Serbia. Veg. Hist. Archaeobot. 2018, 27, 31–44. [Google Scholar] [CrossRef]
- Miladinović-Radmilović, N.; Vitezović, S. Archaeobotany at neolithic sites in Serbia: A critical overview of the methods and results. In Bioarchaeology in the Balkans; Miladinović-Radmilović, N., Vitezović, S., Eds.; Srpsko Arheološko Društvo and Blago Sirmijuma: Beograd, Serbia; Sremska Mitrovica, Serbia, 2013; pp. 25–55. [Google Scholar]
- Filipović, D.; Tasić, N. Vinča-Belo Brdo, a late neolithic site in Serbia consideration of the macro-botanical remains as indicators of dietary habits. Balcanica 2012, 43, 7–27. [Google Scholar] [CrossRef]
- Silva, G.J.; Souza, T.M.; Barbieri, R.L.; Costa de Oliveira, A. Origin, domestication, and dispersing of pear (Pyrus spp.). Adv. Agric. 2014, 2014, 541097. [Google Scholar] [CrossRef] [Green Version]
- Quinet, M.; Wesel, J.-P. Botany and Taxonomy of Pear. In The Pear Genome; Korban, S.S., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 1–33. [Google Scholar]
- da Silva, G.J.; Villa, F.; Grimaldi, F.; da Silva, P.S.; Welter, J.F. Pear (Pyrus spp.) breeding. In Advances in Plant Breeding Strategies: Fruits: Volume 3; Al-Khayri, J.M., Jain, S.M., Johnson, D.V., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 131–163. [Google Scholar]
- Janick, J. The pear in history, literature, popular culture, and art. Acta Hortic. 2002, 596, 41–52. [Google Scholar] [CrossRef]
- Jackson, J.E. The Biology of Apples and Pears; Cambridge University Press: New York, NY, USA, 2003. [Google Scholar]
- Mudge, K.; Janick, J.; Scofield, S.; Goldschmidt, E.E. A history of grafting. Hortic. Rev. 2009, 35, 437–493. [Google Scholar] [CrossRef]
- Dalby, A. Food in the Ancient World from A to Z; Routledge: London, UK; New York, NY, USA, 2003. [Google Scholar]
- Morgan, J. The Book of Pears: The Definitive History and Guide to over 500 Varieties; Chelsea Green Publishing: Totnes, UK, 2015. [Google Scholar]
- Volk, G.M.; Cornille, A. Genetic diversity and domestication history in Pyrus. In The Pear Genome. Compendium of Plant Genomes; Korban, S., Ed.; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Fischer, M. Pear breeding. In Breeding Plantation Tree Crops: Temperate Species; Mohan Jain, S.M., Priyadarshan, P.M., Eds.; Springer ScienceþBusiness Media: New York, NY, USA, 2009; pp. 135–160. [Google Scholar]
- Janick, J. The origins of fruits, fruit growing, and fruit breeding. Plant Breed. Rev. 2005, 25, 255–320. [Google Scholar]
- Hedrick, U.; Howe, G.; Taylor, O.; Francis, E.; Tukey, H. The Pears of New York. J. B. LYON Company: Albany, NY, USA, 1921. [Google Scholar]
- Quinet, M.; Wesel, J. Botany and taxonomy of pear. In The Pear Genome. Compendium of Plant Genomes; Korban, S., Ed.; Springer: Cham, Switzerland, 2019. [Google Scholar]
- Eccher Zerbini, P. The quality of pear fruit. Acta Hortic. 2002, 596, 805–810. [Google Scholar] [CrossRef]
- Baniwal, P.; Hathan, B.S. Physico-chemical, nutritional, functional, textural and morphological characterization of sand pear fruit (Pyrus pyrifolia L.) from northern region of India. Asian J. Chem. 2017, 29, 805–809. [Google Scholar] [CrossRef]
- Reiland, H.; Slavin, J. Systematic review of pears and health. Nutr. Today 2015, 50, 301–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akagić, A.; Oras, A.; Gaši, F.; Meland, M.; Drkenda, P.; Memić, S.; Spaho, N.; Žuljević, S.O.; Jerković, I.; Musić, O.; et al. A comparative study of ten pear (Pyrus communis L.) cultivars in relation to the content of sugars, organic acids, and polyphenol compounds. Foods 2022, 11, 3031. [Google Scholar] [CrossRef]
- Wang, Z.; Barrow, C.J.; Dunshea, F.R.; Suleria, H.A.R. A comparative investigation on phenolic composition, characterization and antioxidant potentials of five different Australian grown pear varieties. Antioxidants 2021, 10, 151. [Google Scholar] [CrossRef]
- Hong, S.-Y.; Lansky, E.; Kang, S.-S.; Yang, M. A review of pears (Pyrus spp.), ancient functional food for modern times. BMC Complement. Med. Ther. 2021, 21, 219. [Google Scholar] [CrossRef]
- James-Martin, G.; Williams, G.; Stonehouse, W.; O’Callaghan, N.; Noakes, M. Health and Nutritional Properties of Pears (Pyrus): A Literature Review; South Australian Health and Medical Research Institute, North Terrace: Adelaide, Australia, 2015. [Google Scholar]
- Manfredini, R.; De Giorgi, A.; Storari, A.; Fabbian, F. Pears and renal stones: Possible weapon for prevention? A comprehensive narrative review. Eur. Rev. Med. Pharmacol. Sci. 2016, 20, 414–425. [Google Scholar]
- Zhang, Y.; Cao, Y.-F.; Huo, H.-L.; Xu, J.-Y.; Tian, L.-M.; Dong, X.-G.; Qi, D.; Liu, C. An assessment of the genetic diversity of pear (Pyrus L.) germplasm resources based on the fruit phenotypic traits. J. Integr. Agric. 2022, 21, 2275–2290. [Google Scholar] [CrossRef]
- Wertheim, S.J.; Wagenmakers, P.S.; Bootsma, J.H.; Groot, M.J. Orchard systems for apple and pear: Conditions for success. Acta Hortic. 2001, 557, 209–227. [Google Scholar] [CrossRef]
- Sestras, R. Ameliorarea Speciilor Horticole; AcademicPres: Cluj-Napoca, Romania, 2004. [Google Scholar]
- Bell, R.L.; Quamme, H.A.; Layne, R.E.C.; Skirvin, R.M. Pears. In Fruit Breeding; Janick, J., Moore, J.N., Eds.; John Wiley and Sons: New York, NY, USA, 1996; pp. 441–514. [Google Scholar]
- Teng, Y. The pear industry and research in China. Acta Hortic. 2011, 909, 161–170. [Google Scholar] [CrossRef]
- Sestras, R.E.; Sestras, A.F. Quantitative traits of interest in apple breeding and their implications for selection. Plants 2023, 12, 903. [Google Scholar] [CrossRef]
- Puskás, M.; Höfer, M.; Sestraş, R.E.; Peil, A.; Sestraş, A.F.; Hanke, M.-V.; Flachowsky, H. Molecular and flow cytometric evaluation of pear (Pyrus L.) genetic resources of the German and Romanian national fruit collections. Genet. Resour. Crop Evol. 2016, 63, 1023–1033. [Google Scholar] [CrossRef]
- Kadkhodaei, S.; Arzani, K.; Yadollahi, A.; Karimzadeh, G.; Abdollahi, H. Genetic diversity and similarity of Asian and European pears (Pyrus spp.) revealed by genome size and morphological traits prediction. Int. J. Fruit Sci. 2021, 21, 619–633. [Google Scholar] [CrossRef]
- Bell, R.L. Pears (Pyrus). Acta Hortic. 1991, 290, 657–700. [Google Scholar] [CrossRef]
- Xu, X.; Fountain, M. Integrated Management of Diseases and Insect Pests of Tree Fruit; Burleigh Dodds Science Publishing Limited: London, UK, 2019. [Google Scholar]
- Vanneste, J.L. Fire Blight: The Disease and Its Causative Agent, Erwinia Amylovora; Cabi Publishing: Wallingford, UK, 2000. [Google Scholar]
- Van Der Zwet, T.; Keil, H.L. Fire Blight: A Bacterial Disease of Rosaceous Plants; US Department of Agriculture: Washington, DC, USA, 1979.
- Sestras, A.F.; Sestras, R.E.; Barbos, A.; Militaru, M. The differences among pear genotypes to fire blight (Erwinia amylovora) attack, based on observations of natural infection. Not. Bot. Horti Agrobot. Cluj-Napoca 2008, 36, 97–103. [Google Scholar] [CrossRef]
- Sardella, D.; Muscat, A.; Brincat, J.-P.; Gatt, R.; Decelis, S.; Valdramidis, V. A comprehensive review of the pear fungal diseases. Int. J. Fruit Sci. 2016, 16, 351–377. [Google Scholar] [CrossRef]
- Horton, D.R. Monitoring of pear psylla for pest management decisions and research. J. Integr. Pest Manag. 1999, 4, 1–20. [Google Scholar] [CrossRef]
- Nin, S.; Giordani, E.; Sacchetti, P.; Ferri, A. Pear resistance to psilla (Cacopsylla pyri L.): A review. Adv. Hortic. Sci. 2012, 26, 59–74. [Google Scholar]
- Simionca Mărcășan, L.I.; Hulujan, I.B.; Florian, T.; Somsai, P.A.; Militaru, M.; Sestras, A.F.; Oltean, I.; Sestras, R.E. The importance of assessing the population structure and biology of psylla species for pest monitoring and management in pear orchards. Not. Bot. Horti Agrobot. Cluj-Napoca 2022, 50, 13022. [Google Scholar] [CrossRef]
- Sestras, A.; Somsai, P.; Militaru, M.; Mitre, V.; Ercişli, S.; Sestras, R. The response of pear cultivars and wild species of Pyrus to Psylla sp. attack, depending on genotype, based on eggs’ and nymphs’ presence on the leaves, before and after the treatment with insecticide. Acta Hortic. 2020, 1289, 79–90. [Google Scholar] [CrossRef]
- Samoilova, A.V.; Leclerque, A. PCR-based Identification of Erwinia amylovora bacteriophages isolated in the Republic of Moldova. J. Virol. Microb. 2014, 1–9. [Google Scholar] [CrossRef]
- Shaltiel-Harpaz, L.; Soroker, V.; Kedoshim, R.; Hason, R.; Sokalsky, T.; Hatib, K.; Bar-Ya’akov, I.; Holland, D. Two pear accessions evaluated for susceptibility to pear psylla Cacopsylla bidens (Šulc) in Israel. Pest Manag. Sci. 2014, 70, 234–239. [Google Scholar] [CrossRef]
- Cimpoieş, G. Pomicultura Speciala; Colograf-Com: Chişinău, Moldova, 2002; p. 336. [Google Scholar]
- Militaru, M.; Braniste, N.; Butac, M.; Sestras, A.; Sotiropoulos, T.; Lukić, M.; Ambrozič Turk, B.; Dzhuvinov, V. Review of pome fruit breeding in Balkan. Acta Hortic. 2013, 981, 177–184. [Google Scholar] [CrossRef]
- Lespinasse, Y.; Chevalier, M.; Durel, C.E.; Guérif, P.H.; Tellier, M.; Denancé, C.; Belouin, A.; Robert, P.H. Pear breeding for scab and psylla resistance. Acta Hortic. 2008, 800, 475–482. [Google Scholar] [CrossRef]
- Sestras, A.F.; Pamfil, D.; Dan, C.; Bolboaca, S.D.; Jäntschi, L.; Sestras, R.E. Possibilities to improve apple scab (Venturia inaequalis (Cke.) Wint.) and powdery mildew [Podosphaera leucotricha (Ell. et Everh.) Salm.] resistance on apple by increasing genetic diversity using potentials of wild species. Aust. J. Crop Sci. 2011, 5, 748–755. [Google Scholar]
- Sestras, R.E.; Pamfil, D.; Ardelean, M.; Botez, C.; Sestras, A.F.; Mitre, I.; Dan, C.; Mihalte, L. Use of phenotypic and MAS selection based on bulk segregant analysis to reveal the genetic variability induced by artificial hybridization in apple. Not. Bot. Horti Agrobot. Cluj-Napoca 2009, 37, 273–277. [Google Scholar] [CrossRef]
- Dan, C.; Sestras, A.; Bozdog, C.; Sestras, R. Investigation of wild species potential to increase genetic diversity useful for apple breeding. Genetika 2015, 47, 993–1011. [Google Scholar] [CrossRef]
- Brewer, L.R.; Palmer, J.W. Global pear breeding programmes: Goals, trends and progress for new cultivars and new rootstocks. Acta Hortic. 2011, 909, 105–119. [Google Scholar] [CrossRef]
- Bell, R.L.; Itai, A. Pyrus. In Wild Crop Relatives: Genomic and Breeding Resources: Temperate Fruits; Kole, C., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 147–177. [Google Scholar]
- Brewer, L.; Volz, R. Genetics and breeding of pear. In The Pear Genome; Korban, S.S., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 63–101. [Google Scholar]
- Khan, A.; Korban, S.S. Breeding and genetics of disease resistance in temperate fruit trees: Challenges and new opportunities. Theor. Appl. Genet. 2022, 135, 3961–3985. [Google Scholar] [CrossRef] [PubMed]
- Dondini, L.; Sansavini, S. European Pear. In Fruit Breeding; Badenes, M.L., Byrne, D.H., Eds.; Springer: Boston, MA, USA, 2012; pp. 369–413. [Google Scholar]
- Laurens, F.; Aranzana, M.J.; Arús, P.; Bassi, D.; Bonany, J.; Corelli, L.; Durel, C.E.; Mes, J.; Pascal, T.; Patocchi, A.; et al. Review of fruit genetics and breeding programmes and a new European initiative to increase fruit breeding efficiency. Acta Hortic. 2012, 929, 95–102. [Google Scholar] [CrossRef]
- Postman, J.; Volk, G.; Aldwinckle, H. Standardized plant disease evaluations will enhance resistance gene discovery. HortScience 2010, 45, 1317–1320. [Google Scholar] [CrossRef]
- Cociu, V.; Oprea, S. Metode de Cercetare în Ameliorarea Plantelor Pomicole; Dacia: Cluj-Napoca, Romania, 1989. [Google Scholar]
- Pârvu, M. Ghid Practic de Fitopatologie; Presa Universitară Clujeană: Cluj-Napoca, Romania, 2010. [Google Scholar]
- Lateur, M.; Populer, C. Screening fruit tree genetic resources in Belgium for disease resistance and other desirable characters. Euphytica 1994, 77, 147–153. [Google Scholar] [CrossRef]
- Patocchi, A.; Frei, A.; Frey, J.E.; Kellerhals, M. Towards improvement of marker assisted selection of apple scab resistant cultivars: Venturia inaequalis virulence surveys and standardization of molecular marker alleles associated with resistance genes. Mol. Breed. 2009, 24, 337–347. [Google Scholar] [CrossRef]
- Patocchi, A.; Wehrli, A.; Dubuis, P.-H.; Auwerkerken, A.; Leida, C.; Cipriani, G.; Passey, T.; Staples, M.; Didelot, F.; Philion, V.; et al. Ten years of VINQUEST: First insight for breeding new apple cultivars with durable apple scab resistance. Am. Phytopathol. Soc. (APS) 2020, 104, 2074–2081. [Google Scholar] [CrossRef] [Green Version]
- Nutter, F.J.; Teng, P.; Shokes, F. Disease assessment terms and concepts. Plant Dis. Rep. 1991, 75, 1187–1188. [Google Scholar]
- Bock, C.H.; El Jarroudi, M.; Kouadio, A.L.; Mackels, C.; Chiang, K.S.; Delfosse, P. Disease severity estimates—Effects of rater accuracy and assessment methods for comparing treatments. Plant Dis. 2015, 99, 1104–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bock, C.H.; Chiang, K.-S.; Del Ponte, E.M. Accuracy of plant specimen disease severity estimates: Concepts, history, methods, ramifications and challenges for the future. CAB Rev. 2016, 11, 1–21. [Google Scholar] [CrossRef]
- Chiang, K.S.; Liu, H.I.; Chen, Y.L.; El Jarroudi, M.; Bock, C.H. Quantitative ordinal scale estimates of plant disease severity: Comparing treatments using a proportional odds model. Phytopathology 2020, 110, 734–743. [Google Scholar] [CrossRef]
- Chiang, K.-S.; Bock, C.H. Understanding the ramifications of quantitative ordinal scales on accuracy of estimates of disease severity and data analysis in plant pathology. Trop. Plant Pathol. 2022, 47, 58–73. [Google Scholar] [CrossRef] [PubMed]
- Postman, J.D.; Spotts, R.A.; Calabro, J. Scab resistance in Pyrus germplasm. Acta Hortic. 2005, 671, 601–608. [Google Scholar] [CrossRef]
- Perchepied, L.; Leforestier, D.; Ravon, E.; Guérif, P.; Denancé, C.; Tellier, M.; Terakami, S.; Yamamoto, T.; Chevalier, M.; Lespinasse, Y.; et al. Genetic mapping and pyramiding of two new pear scab resistance QTLs. Mol. Breed. 2015, 35, 197. [Google Scholar] [CrossRef]
- Bouvier, L.; Bourcy, M.; Boulay, M.; Tellier, M.; Guérif, P.; Denancé, C.; Durel, C.-E.; Lespinasse, Y. A new pear scab resistance gene Rvp1 from the European pear cultivar ‘Navara’ maps in a genomic region syntenic to an apple scab resistance gene cluster on linkage group 2. Tree Genet. Genomes 2012, 8, 53–60. [Google Scholar] [CrossRef] [Green Version]
- Chevalier, M.; Bernard, C.; Tellier, M.; Lespinasse, Y.; Filmond, R.; Le Lezec, M. Variability in the reaction of several pear (Pyrus communis) cultivars to different inocula of Venturia pyrina. Acta Hortic. 2004, 663, 177–181. [Google Scholar] [CrossRef]
- Lācis, G.; Lāce, B.; Blukmanis, M. Evaluation of the susceptibility of pear cultivars to scab (Venturia pirina Aderh.). Acta Hortic. 2015, 1099, 741–747. [Google Scholar] [CrossRef]
- Andries, N. Achievements and prospectives in pear breeding at the Fruit Research Station Voinesti, Romania. Acta Hortic. 2002, 596, 261–264. [Google Scholar] [CrossRef]
- Braniste, N.; Andries, N.; Ghidra, V. Pear genetic breeding to improve the Romanian varieties. Acta Hortic. 2008, 800, 491–496. [Google Scholar] [CrossRef]
- Pierantoni, L.; Dondini, L.; Cho, K.H.; Shin, I.S.; Gennari, F.; Chiodini, R.; Tartarini, S.; Kang, S.J.; Sansavini, S. Pear scab resistance QTLs via a European pear (Pyrus communis) linkage map. Tree Genet. Genomes 2007, 3, 311–317. [Google Scholar] [CrossRef]
- Won, K.; Bastiaanse, H.; Kim, Y.K.; Song, J.H.; Kang, S.S.; Lee, H.C.; Cho, K.H.; Brewer, L.; Singla, G.; Gardiner, S.E.; et al. Genetic mapping of polygenic scab (Venturia pirina) resistance in an interspecific pear family. Mol. Breed. 2014, 34, 2179–2189. [Google Scholar] [CrossRef]
- Abe, K.; Kotobuki, K.; Saito, T.; Terai, O. Inheritance of resistance to pear scab from European pears to Asian pears. J. Jpn. Soc. Hort. Sci. 2000, 69, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Chatzidimopoulos, M.; Pappas, A.C. Epidemiology and control of Septoria pyricola in pear leaf and fruit. J. Plant Pathol. 2016, 98, 447–452. [Google Scholar]
- Sestras, A.; Titirica Ancu, I.; Dan, C.; Andrecan, A.; Mitre, V.; Sestras, R.; Militaru, M. Yield and response to the main diseases attack of pear cultivars and heritability of the traits. Lucr. St. Seria Hort. USAMV Iaşi 2018, 61, 91–96. [Google Scholar]
- Sestras, A.; Sestras, R.; Pamfil, D.; Barbos, A. Combining ability effects of several pear cultivars used as genitors for Mycosphaerella sentina resistance. Bull. UASVM Hort. 2008, 65, 48–52. [Google Scholar]
- Thibault, B.; Lecomte, P.; Hermann, L.; Belouin, A. Assessment of the susceptibility to Erwinia amylovora of 90 varieties or selections of pear. Acta Hortic. 1987, 217, 305–310. [Google Scholar] [CrossRef]
- Pimentel, D. Fire blight Erwinia amylovora. InnovPlantProtect 2021, 1–24. [Google Scholar]
- Saygili, H.; Türküsay, H.; Hepaksoy, S.; Ünal, A.; Can, H.Z. Investigation on determining some pear varieties resistant to fire blight (Erwinia amylovora (Burrill) Winslow et al.). Acta Hortic. 1999, 225–228. [Google Scholar] [CrossRef]
- Korba, J.; Šillerová, J.; Paprštein, F.; Sedlák, J.; Prokinová, E.; Hošková, P. Evaluation of susceptibility level of pear cultivars to fire blight (Erwinia amylovora) in the Czech Republic. Hortic. Sci. 2013, 40, 58–64. [Google Scholar] [CrossRef] [Green Version]
- Dondini, L.; Pierantoni, L.; Gaiotti, F.; Chiodini, R.; Tartarini, S.; Bazzi, C.; Sansavini, S. Identifying QTLs for fire-blight resistance via a European pear (Pyrus communis L.) genetic linkage map. Mol. Breed. 2005, 14, 407–418. [Google Scholar] [CrossRef]
- Calenge, F.; Drouet, D.; Denancé, C.; van de Weg, W.E.; Brisset, M.N.; Paulin, J.P.; Durel, C.E. Identification of a major QTL together with several minor additive or epistatic QTLs for resistance to fire blight in apple in two related progenies. Theor. Appl. Genet. 2005, 111, 128–135. [Google Scholar] [CrossRef] [PubMed]
- Peil, A.; Bus, V.G.; Geider, K.; Richter, K.; Flachowsky, H.; Hanke, M.-V. Improvement of fire blight resistance in apple and pear. Int. J. Plant Breed. 2009, 3, 1–27. [Google Scholar]
- Evrenosoğlu, Y.; Mertoğlu, K.; Acarsoy Bilgin, N.; Misirli, A.; Özsoy, A.N. Inheritance pattern of fire blight resistance in pear. Sci. Hortic. 2019, 246, 887–892. [Google Scholar] [CrossRef]
- Rosati, C.; Rivalta, L.; Dradi, M.; Le Lézec, M.; Belouin, A.; Chartier, R. Fireblight evaluation of advanced Italian selections and cultivars of pear. Acta Hortic. 2002, 279–282. [Google Scholar] [CrossRef]
- Evrenosoğlu, Y.; Mertoğlu, K.; Acarsoy Bilgin, N.; Misirli, A.; Altay, Y. An analysis on some reciprocal pear hybridization combinations in terms of transferring resistance to fire blight. Erwerbs-Obstbau 2020, 62, 189–194. [Google Scholar] [CrossRef]
- Jakab-Ilyefalvi, Z.; Platon, I.; Festila, A. Fire blight susceptibility of some pear varieties (Erwinia amylovora, Burill). Fruit Grow. Res. 2012, 28, 25–31. [Google Scholar]
- Paraschivu, M.; Ciobanu, A.; Cotuna, O.; Paraschivu, M. Assessment of the bacterium Erwinia amylovora attack on several pear varieties (Pyrus communis L.) and the influence on fruits sugar content. Sci. Pap. Ser. B Hortic. 2020, 64, 163–168. [Google Scholar]
- Van Der Zwet, T.; Oitto, W.A.; Westwood, M.N. Variability in degree of fire blight resistance within and between Pyrus species, interspecific hybrids, and seedling progenies. Euphytica 1974, 23, 295–304. [Google Scholar] [CrossRef]
- Abdollahi, H.; Rugini, E.; Ruzzi, M.; Muleo, R. In Vitro system for studying the interaction between Erwinia amylovora and genotypes of pear. Plant Cell Tissue Organ Cult. 2004, 79, 203–212. [Google Scholar] [CrossRef]
- Sillerova, J.; Korba, J.; Paprstein, F.; Sedlak, J. Testing of resistance of pear cultivars after artificial inoculation with Erwinia amylovora in field conditions. Acta Hortic. 2011, 896, 353–355. [Google Scholar] [CrossRef]
- Van Der Zwet, T.; Beer, S.V. Fire Blight-Its Nature, Prevention, and Control: A Practical Guide to Integrated Disease Management; U.S. Department of Agriculture: Washington, DC, USA, 1999; p. 97.
- Ozrenk, K.; Balta, F.; Çelik, F. Levels of fire blight (Erwinia amylovora) susceptibility of native apple, pear and quince germplasm from Lake Van Basin, Turkey. Eur. J. Plant Pathol. 2012, 132, 229–236. [Google Scholar] [CrossRef]
- Billing, E. Weather affecting the incidence and severity of fire blight in England. Acta Hortic. 2006, 704, 163–170. [Google Scholar] [CrossRef]
- Westigard, P.; Zwick, R. The Pear Psylla in Oregon; Oregon State University: Salem, OR, USA, 1972; p. 22. [Google Scholar]
- Bell, R.L. Effect of resistant and susceptible East European pears on development and mortality of the pear psylla, Cacopsylla pyricola (Förster). HortScience 2015, 50, 661–665. [Google Scholar] [CrossRef] [Green Version]
- Bell, R.L.; Stuart, L.C. Resistance in Eastern European pyrus germplasm to pear psylla nymphal feeding. HortScience 1990, 25, 789–791. [Google Scholar] [CrossRef] [Green Version]
- Bell, R.L. Additional East European Pyrus germplasm with resistance to pear psylla nymphal feeding. HortScience 1992, 27, 412–413. [Google Scholar] [CrossRef] [Green Version]
- van der Zwet, T.; Cociu, V.; Czarnecki, B.; Nyeki, J.; Blazek, J. Collecting Pyrus Germplasm in Romania, Poland, Hungary, and Czechoslovakia. HortScience 1989, 24, 420–424. [Google Scholar] [CrossRef]
- Sehic, J.; Garkava-Gustavsson, L.; Fernández-Fernández, F.; Nybom, H. Genetic diversity in a collection of European pear (Pyrus communis) cultivars determined with SSR markers chosen by ECPGR. Sci. Hortic. 2012, 145, 39–45. [Google Scholar] [CrossRef]
- Evans, K.M.; Fernández-Fernández, F.; Govan, C. Harmonising fingerprinting protocols to allow comparisons between germplasm collections—Pyrus. Acta Hortic. 2009, 103–106. [Google Scholar] [CrossRef]
- Le Roux, P.-M.F.; Christen, D.; Duffy, B.; Tartarini, S.; Dondini, L.; Yamamoto, T.; Nishitani, C.; Terakami, S.; Lespinasse, Y.; Kellerhals, M.; et al. Redefinition of the map position and validation of a major quantitative trait locus for fire blight resistance of the pear cultivar ‘Harrow Sweet’ (Pyrus communis L.). Plant Breed. 2012, 131, 656–664. [Google Scholar] [CrossRef]
- Kocsisné, G.M.; Bolla, D.; Anhalt-Brüderl, U.C.M.; Forneck, A.; Taller, J.; Kocsis, L. Genetic diversity and similarity of pear (Pyrus communis L.) cultivars in Central Europe revealed by SSR markers. Genet. Resour. Crop Evol. 2020, 67, 1755–1763. [Google Scholar] [CrossRef]
- Brini, W.; Mars, M.; Hormaza, J.I. Genetic diversity in local Tunisian pears (Pyrus communis L.) studied with SSR markers. Sci. Hortic. 2008, 115, 337–341. [Google Scholar] [CrossRef]
- Queiroz, Á.; Bagoin Guimarães, J.; Sánchez, C.; Simões, F.; Maia de Sousa, R.; Viegas, W.; Veloso, M.M. Genetic diversity and structure of the Portuguese pear (Pyrus communis L.) germplasm. Sustainability 2019, 11, 5340. [Google Scholar] [CrossRef] [Green Version]
- Bielsa, F.J.; Irisarri, P.; Errea, P.; Pina, A. Genetic diversity and structure of local pear cultivars from mountainous areas from Aragon (Northeastern Spain). Agronomy 2021, 11, 1778. [Google Scholar] [CrossRef]
- Bell, R.L. Genetics, genomics, and breeding for fire blight resistance in pear. In The Pear Genome; Korban, S.S., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 243–264. [Google Scholar]
- Sharma, R.; Kumar, B.; Arora, R.; Ahlawat, S.; Mishra, A.K.; Tantia, M.S. Genetic diversity estimates point to immediate efforts for conserving the endangered Tibetan sheep of India. Meta Gene 2016, 8, 14–20. [Google Scholar] [CrossRef]
- Postman, J.D. Intergeneric hybrids in Pyrinae (=Maloideae) subtribe of Pyreae in Family Rosaceae at USDA Genebank. Acta Hortic. 2011, 918, 937–943. [Google Scholar] [CrossRef]
- Bergonzoni, L.; Alessandri, S.; Domenichini, C.; Dondini, L.; Caracciolo, G.; Pietrella, M.; Baruzzi, G.; Tartarini, S. Characterization of red-fleshed pear accessions from Emilia-Romagna region. Sci. Hortic. 2023, 312, 111857. [Google Scholar] [CrossRef]
- Draga, S.; Palumbo, F.; Miracolo Barbagiovanni, I.; Pati, F.; Barcaccia, G. Management of genetic erosion: The (successful) case study of the pear (Pyrus communis L.) germplasm of the Lazio region (Italy). Front. Plant Sci. 2023, 13. [Google Scholar] [CrossRef]
- Zurn, J.D.; Nyberg, A.; Montanari, S.; Postman, J.; Neale, D.; Bassil, N. A new SSR fingerprinting set and its comparison to existing SSR-and SNP-based genotyping platforms to manage Pyrus germplasm resources. Tree Genet. Genomes 2020, 16, 72. [Google Scholar] [CrossRef]
- Baccichet, I.; Foria, S.; Messina, R.; Peccol, E.; Losa, A.; Fabro, M.; Gori, G.; Zandigiacomo, P.; Cipriani, G.; Testolin, R. Genetic and ploidy diversity of pear (Pyrus spp.) germplasm of Friuli Venezia Giulia, Italy. Genet. Resour. Crop Evol. 2020, 67, 83–96. [Google Scholar] [CrossRef]
- Xie, W.; Yu, K.; Pauls, K.P.; Navabi, A. Application of image analysis in studies of quantitative disease resistance, exemplified using common bacterial blight–common bean pathosystem. Phytopathology 2012, 102, 434–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bock, C.H.; Poole, G.H.; Parker, P.E.; Gottwald, T.R. Plant disease severity estimated visually, by digital photography and image analysis, and by hyperspectral imaging. Crit. Rev. Plant Sci. 2010, 29, 59–107. [Google Scholar] [CrossRef]
- De Franceschi, P.; Dondini, L. Molecular mapping of major genes and QTLs in pear. In The Pear Genome; Korban, S.S., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 113–131. [Google Scholar]
- Matei, I.; Pacurar, I.; Rosca, S.; Bilasco, S.; Sestras, P.; Rusu, T.; Jude, E.T.; Tăut, F.D. Land use favourability assessment based on soil characteristics and anthropic pollution. Case study Somesul Mic Valley Corridor, Romania. Agronomy 2020, 10, 1245. [Google Scholar] [CrossRef]
- Van der Zwet, T.; Oitto, W.; Brooks, H. Scoring system for rating the severity of fireblight in pear. Plant Dis. Rep. 1970, 54, 835–839. [Google Scholar]
- Vilanova, S.; Alonso, D.; Gramazio, P.; Plazas, M.; García-Fortea, E.; Ferrante, P.; Schmidt, M.; Díez, M.J.; Usadel, B.; Giuliano, G.; et al. SILEX: A fast and inexpensive high-quality DNA extraction method suitable for multiple sequencing platforms and recalcitrant plant species. Plant Methods 2020, 16, 110. [Google Scholar] [CrossRef]
- Liu, K.; Muse, S.V. PowerMarker: An integrated analysis environment for genetic marker analysis. Bioinformatics 2005, 21, 2128–2129. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 4–9. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Code | Species | Venturia pyrina | Septoria pyricola | Erwinia amylovora | Psylla sp. |
|---|---|---|---|---|---|
| 1 | ×Pyronia veitkii | 9.4 ± 0.6 a | 2.8 ± 0.2 a | 0.0 ± 0.0 e | 1.7 ± 0.4 b |
| 2 | Pyrus betulaefolia | 3.8 ± 0.6 d | 2.2 ± 0.5 a,b | 20.6 ± 3.6 b,c | 0.3 ± 0.0 d |
| 4 | Pyrus caucasica | 6.7 ± 0.7 b | 2.6 ± 0.9 a | 0.0 ± 0.0 e | 1.4 ± 0.4 b,c |
| 5 | Pyrus pyraster | 5.5 ± 0.7 b,c | 2.0 ± 0.4 a–c | 0.0 ± 0.0 e | 0.1 ± 0.0 d |
| 6 | Pyrus communis | 4.8 ± 0.8 c,d | 3.4 ± 0.6 a | 0.4 ± 0.1 e | 3.9 ± 1.2 a |
| 7 | Pyrus cordata | 1.5 ± 0.4 f,g | 2.4 ± 0.8 a,b | 19.2 ± 2.7 b,c | 0.5 ± 0.1 c,d |
| 9 | Pyrus eleagrifolia | 0.5 ± 0.2 g | 1.6 ± 0.3 a–c | 24.1 ± 1.6 b | 0.4 ± 0.1 c,d |
| 12 | Pyrus lindlezi | 0.2 ± 0.1 g | 2.1 ± 0.2 a,b | 0.0 ± 0.0 e | 0.0 ± 0.0 d |
| 13 | Pyrus longipes | 0.2 ± 0.1 g | 1.5 ± 0.3 a–c | 7.3 ± 0.5 d | 0.3 ± 0.0 d |
| 14 | Pyrus malifolia | 3.6 ± 0.8 d,e | 3.4 ± 1.5 a | 0.0 ± 0.0 e | 0.2 ± 0.0 d |
| 15 | Pyrus nivalis | 0.4 ± 0.3 g | 0.4 ± 0.1 b,c | 32.4 ± 3.4 a | 0.1 ± 0.0 d |
| 16 | Pyrus persica | 0.0 ± 0.0 g | 0.0 ± 0.0 c | 0.0 ± 0.0 e | 0.0 ± 0.0 d |
| 17 | ×Sorbopyrus | 2.3 ± 0.4 e,f | 0.5 ± 0.2 b,c | 17.3 ± 1.6 c | 0.0 ± 0.0 d |
| Average value | 3.0 | 1.9 | 9.3 | 0.7 |
| Code | Cultivar | Venturia pyrina | Septoria pyricola | Erwinia amylovora | Psylla sp. |
|---|---|---|---|---|---|
| 21 | Argessis | 4.8 ± 0.5 a–c | 0.8 ± 0.2 e–h | 19.3 ± 3.1 b | 2.6 ± 0.5 c–e |
| 28 | Cântări | 5.9 ± 1.0 a | 1.9 ± 0.3 a–d | 20.7 ± 4.7 b | 1.6 ± 0.3 d,e |
| 34 | Cu miez roşu | 2.0 ± 0.3 f,g | 2.0 ± 0.2 a–c | 15.1 ± 3.3 b,c | 1.9 ± 0.3 d,e |
| 88 | Republica | 5.5 ± 0.9 a | 0.3 ± 0.2 h | 33.7 ± 4.8 a | 1.9 ± 0.2 d,e |
| 106 | Zaharoasă de vară | 3.2 ± 0.7 d–f | 1.6 ± 0.5 b–e | 0.0 ± 0.0 f | 2.5 ± 0.9 c–e |
| 107 | Adria | 2.7 ± 0.4 d–g | 1.2 ± 0.2 d–g | 0.0 ± 0.0 f | 1.0 ± 0.0 e |
| 109 | Doina | 3.5 ± 0.3 c–e | 2.1 ± 0.2 ab | 14.8 ± 2.7 b–d | 4.9 ± 1.1 a,b |
| 110 | Haydeea | 0.1 ± 0.0 h | 0.8 ± 0.1 e–h | 0.0 ± 0.0 f | 0.4 ± 0.1 e |
| 111 | Ina Estival | 1.7 ± 0.4 g | 1.1 ± 0.2 e–g | 0.0 ± 0.0 f | 2.1 ± 0.5 d,e |
| 112 | Jubileu 50 | 2.5 ± 0.6 e–g | 0.6 ± 0.1 g,h | 7.6 ± 1.1 e | 2.1 ± 0.1 d,e |
| 114 | Meda | 4.0 ± 0.3 b–d | 2.0 ± 0.2 a–c | 0.0 ± 0.0 f | 3.3 ± 0.9 b–d |
| 115 | Milenium | 2.8 ± 0.4 d–g | 2.5 ± 0.4 a | 8.1 ± 1.5 d,e | 4.1 ± 1.1 a–c |
| 116 | Napoca | 1.7 ± 0.4 g | 1.4 ± 0.3 c–f | 0.0 ± 0.0 f | 2.6 ± 0.8 c–e |
| 117 | Primadona | 3.7 ± 0.6 c–e | 0.2 ± 0.1 h | 6.6 ± 1.4 e | 2.1 ± 0.1 d,e |
| 118 | Roșioară de Cluj | 4.8 ± 0.6 a–c | 2.1 ± 0.1 a,b | 10.2 ± 2.0 c–e | 5.1 ± 1.1 a |
| 119 | Transilvania | 2.0 ± 0.3 f,g | 1.9 ± 0.2 a–d | 8.9 ± 0.8 c–e | 2.3 ± 0.6 c–e |
| 120 | Virgiliu Hibernal | 5.3 ± 0.7 a,b | 2.0 ± 0.4 a–c | 9.1 ± 2.6 c–e | 5.0 ± 0.6 a,b |
| Average value | 3.3 | 1.4 | 9.1 | 2.7 |
| Code | Cultivar | Venturia pyrina | Septoria pyricola | Erwinia amylovora | Psylla sp. |
|---|---|---|---|---|---|
| 122 | Abate Fetel | 6.7 ± 2.2 d–g | 0.4 ± 0.1 e–g | 35.3 ± 5.8 a–d | 1.3 ± 0.3 i–m |
| 127 | Arabitka | 5.3 ± 1.2 e–h | 2.0 ± 0.6 c,d | 4.3 ± 0.9 k–n | 3.7 ± 0.9 f–k |
| 136 | Bergamotte Esperen | 4.0 ± 0.6 g–i | 1.7 ± 0.3 c–e | 20.0 ± 6.0 c–k | 4.0 ± 1.0 f–j |
| 137 | Beurré Amanlis | 3.0 ± 0.6 g–i | 1.3 ± 0.3 d–g | 0.0 ± 0.0 n | 8.3 ± 1.5 c,d |
| 138 | Beurré Bachelier | 2.4 ± 0.8 h,i | 0.5 ± 0.1 e–g | 0.0 ± 0.0 n | 7.7 ± 1.5 c–e |
| 139 | Beurré Bosc | 3.3 ± 0.9 g–i | 0.9 ± 0.3 d–g | 1.8 ± 0.4 m,n | 4.0 ± 0.6 f–j |
| 142 | Beurré Diel | 3.0 ± 0.6 g–i | 1.5 ± 0.3 d–f | 21.7 ± 5.8 c–j | 3.0 ± 0.6 f–m |
| 146 | Beurré du Luçon | 1.6 ± 0.3 h,i | 0.5 ± 0.1 e–g | 0.0 ± 0.0 n | 1.0 ± 0.7 i–m |
| 152 | Blanquet Precoce | 1.5 ± 0.3 h,i | 0.8 ± 0.2 d–g | 0.0 ± 0.0 n | 4.7 ± 0.9 e–h |
| 154 | Bonne Louise d’Avranches | 1.5 ± 0.3 h,i | 1.1 ± 0.3 d–g | 36.7 ± 6.9 a–c | 8.7 ± 1.2 c,d |
| 155 | Bristol Cross | 1.6 ± 0.4 h,i | 1.5 ± 0.3 d–f | 0.0 ± 0.0 n | 2.0 ± 0.6 g–m |
| 156 | Bunte Julibirne | 1.1 ± 0.1 j | 0.6 ± 0.2 e–g | 40.7 ± 9.3 a | 2.1 ± 0.6 g–m |
| 157 | Butirra Precoce Morettini | 0.9 ± 0.1 j | 1.2 ± 0.1 d–g | 40.7 ± 6.8 a,b | 11.7 ± 4.7 b |
| 162 | Chang Pa Li | 1.3 ± 0.2 h,i | 0.6 ± 0.2 e–g | 39.7 ± 11.3 a,b | 0.8 ± 0.2 i–m |
| 167 | Conference | 8.0 ± 1.7 c–e | 4.3 ± 1.2 b | 0.0 ± 0.0 n | 4.2 ± 1.2 f–i |
| 168 | Conseiller de la Cour | 3.1 ± 0.6 g–i | 1.5 ± 0.3 d–f | 0.0 ± 0.0 n | 4.3 ± 0.7 f–i |
| 169 | Curé | 1.0 ± 0.2 j | 0.6 ± 0.2 e–g | 2.0 ± 0.6 m,n | 0.1 ± 0.0 l,m |
| 173 | Beurré Hardy | 1.0 ± 0.1 j | 0.3 ± 0.1 f,g | 3.0 ± 0.6 l–n | 2.0 ± 0.6 g–m |
| 179 | Dr. Jules Guyot | 2.8 ± 0.6 h,i | 0.7 ± 0.2 d–g | 25.7 ± 8.8 b–f | 1.7 ± 0.3 h–m |
| 187 | Beurré Giffard | 6.7 ± 0.9 d–g | 0.5 ± 0.1 e–g | 7.3 ± 2.3 i–n | 2.0 ± 0.6 g–m |
| 188 | Er Shi Shinge | 0.0 ± 0.0 k | 0.5 ± 0.1 e–g | 1.0 ± 0.5 n | 0.2 ± 0.0 l,m |
| 189 | Er Jang Li | 4.2 ± 0.4 f–i | 0.4 ± 0.1 e–g | 21.7 ± 6.2 c–j | 4.7 ± 0.3 e–h |
| 194 | Fondante des Bois | 10.0 ± 1.5 c | 1.3 ± 0.4 d–g | 25.7 ± 9.0 b–f | 3.3 ± 0.3 f–l |
| 197 | General Leclerc | 5.3 ± 1.2 e–h | 0.4 ± 0.1 e–g | 12.3 ± 4.4 f–n | 1.3 ± 0.3 i–m |
| 198 | General Osmanwill | 8.0 ± 0.6 c–e | 0.3 ± 0.1 f,g | 0.0 ± 0.0 n | 0.3 ± 0.1 l,m |
| 202 | Grand Champion | 10.7 ± 0.9 c | 4.3 ± 1.2 b | 0.0 ± 0.0 n | 4.0 ± 0.6 f–j |
| 209 | Imperial | 1.2 ± 0.1 j | 0.3 ± 0.1 f,g | 2.3 ± 0.9 m,n | 7.7 ± 1.8 c–e |
| 213 | Juliusi Selimesi | 1.4 ± 0.3 h,i | 0.4 ± 0.1 e–g | 3.7 ± 0.9 l–n | 1.7 ± 0.3 h–m |
| 221 | Kostliche Von Germen | 15.0 ± 1.5 b | 1.1 ± 0.3 d–g | 6.3 ± 3.0 j–n | 4.7 ± 1.5 e–h |
| 222 | Kristalli | 0.0 ± 0.0 k | 7.0 ± 1.2 a | 22.0 ± 4.9 c–j | 14.7 ± 3.2 a |
| 224 | Laurence | 3.0 ± 0.6 g–i | 1.2 ± 0.2 d–g | 0.0 ± 0.0 n | 3.3 ± 0.9 f–l |
| 225 | Laxton Superb | 20.3 ± 1.2 a | 0.5 ± 0.2 e–g | 4.3 ± 1.2 k–n | 0.1 ± 0.0 l,m |
| 229 | Lincoln | 8.3 ± 1.8 c–e | 1.2 ± 0.4 d–g | 17.7 ± 3.3 e–m | 2.3 ± 0.9 g–m |
| 232 | Madame Ballet | 7.7 ± 2.0 c–f | 3.3 ± 0.9 b,c | 9.0 ± 1.2 h–n | 0.2 ± 0.1 l,m |
| 233 | Magness | 4.0 ± 1.5 g–i | 0.3 ± 0.1 f,g | 0.0 ± 0.0 n | 0.3 ± 0.1 l,m |
| 240 | Moonglow | 18.3 ± 3.7 a | 0.4 ± 0.1 e–g | 32.0 ± 5.8 a–e | 0.7 ± 0.1 k–m |
| 246 | Noiabriscaia | 10.3 ± 1.9 c | 0.8 ± 0.1 d–g | 24.3 ± 6.9 c–h | 9.7 ± 1.2 b,c |
| 248 | Okusankichi | 0.0 ± 0.0 k | 0.6 ± 0.1 e–g | 25.7 ± 10.1 b–f | 0.1 ± 0.0 l,m |
| 250 | Olivier de Serres | 0.0 ± 0.0 k | 0.5 ± 0.2 e–g | 24.7 ± 8.6 c–g | 0.0 ± 0.0 m |
| 258 | Pierre Corneille | 2.3 ± 0.3 h,i | 0.2 ± 0.1 e–g | 22.7 ± 5.8 c–i | 1.0 ± 0.5 i–m |
| 259 | Pitmaston Duchess | 2.0 ± 0.6 h,i | 0.7 ± 0.1 d–g | 18.7 ± 2.3 e–l | 1.7 ± 0.3 h–m |
| 260 | Plovdivka Parva | 0.3 ± 0.3 j | 0.6 ± 0.1 e–g | 4.0 ± 1.5 l–n | 1.3 ± 0.3 i–m |
| 263 | Précoce Trottier | 0.0 ± 0.0 k | 0.5 ± 0.2 e–g | 0.0 ± 0.0 n | 0.1 ± 0.0 l,m |
| 272 | Seigneur Esperen | 2.0 ± 0.6 h,i | 0.5 ± 0.1 e–g | 0.0 ± 0.0 n | 1.3 ± 0.3 i–m |
| 286 | Triomphe de Jodoigne | 14.0 ± 3.5 b | 0.6 ± 0.1 e–g | 9.0 ± 1.2 h–n | 0.2 ± 0.1 l,m |
| 288 | Triomphe de Vienne | 2.0 ± 0.6 h,i | 0.6 ± 0.2 e–g | 18.3 ± 3.7 e–l | 2.3 ± 0.9 g–m |
| 289 | Van Mons | 9.3 ± 0.9 c,d | 2.7 ± 1.2 c | 10.0 ± 2.1 g–n | 1.7 ± 0.3 h–m |
| 293 | Williams | 0.7 ± 0.2 j | 0.2 ± 0.1 h | 0.0 ± 0.0 n | 5.0 ± 0.6 e–g |
| 294 | Williams Bovey | 0.9 ± 0.1 j | 0.1 ± 0.1 h | 0.0 ± 0.0 n | 4.0 ±0.6 f–j |
| 296 | Williams Red | 1.0 ± 0.2 j | 1.1 ± 0.1 d–g | 35.0 ± 6.1 a–d | 6.0 ± 0.6 d–f |
| Average value | 4.4 | 1.1 | 12.6 | 3.2 |
| SSR Marker | Major Allele Frequency | NG | No. Obs. | Na | Ava | Ho | He | PIC |
|---|---|---|---|---|---|---|---|---|
| 01D08 | 0.322 | 24 | 59 | 13 | 0.738 | 0.729 | 0.796 | 0.769 |
| 04E03A | 0.620 | 6 | 79 | 4 | 0.988 | 0.608 | 0.543 | 0.485 |
| 04E03B | 0.297 | 18 | 59 | 12 | 0.738 | 0.237 | 0.823 | 0.803 |
| 03D12 | 0.278 | 32 | 72 | 18 | 0.900 | 0.611 | 0.824 | 0.805 |
| 01D09 | 0.232 | 36 | 56 | 21 | 0.700 | 0.607 | 0.880 | 0.870 |
| EMPC117 | 0.350 | 25 | 70 | 16 | 0.875 | 0.443 | 0.758 | 0.726 |
| 01F07A | 0.257 | 34 | 72 | 20 | 0.900 | 0.764 | 0.864 | 0.851 |
| 03G07 | 0.544 | 24 | 68 | 14 | 0.850 | 0.471 | 0.669 | 0.647 |
| 05C06A | 0.571 | 19 | 77 | 13 | 0.963 | 0.247 | 0.642 | 0.621 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Simionca Mărcășan, L.I.; Oltean, I.; Popa, S.; Plazas, M.; Vilanova, S.; Gramazio, P.; Sestras, A.F.; Prohens, J.; Sestras, R.E. Comparative Analysis of Phenotypic and Molecular Data on Response to Main Pear Diseases and Pest Attack in a Germplasm Collection. Int. J. Mol. Sci. 2023, 24, 6239. https://doi.org/10.3390/ijms24076239
Simionca Mărcășan LI, Oltean I, Popa S, Plazas M, Vilanova S, Gramazio P, Sestras AF, Prohens J, Sestras RE. Comparative Analysis of Phenotypic and Molecular Data on Response to Main Pear Diseases and Pest Attack in a Germplasm Collection. International Journal of Molecular Sciences. 2023; 24(7):6239. https://doi.org/10.3390/ijms24076239
Chicago/Turabian StyleSimionca Mărcășan, Leontina I., Ion Oltean, Sergiu Popa, Mariola Plazas, Santiago Vilanova, Pietro Gramazio, Adriana F. Sestras, Jaime Prohens, and Radu E. Sestras. 2023. "Comparative Analysis of Phenotypic and Molecular Data on Response to Main Pear Diseases and Pest Attack in a Germplasm Collection" International Journal of Molecular Sciences 24, no. 7: 6239. https://doi.org/10.3390/ijms24076239
APA StyleSimionca Mărcășan, L. I., Oltean, I., Popa, S., Plazas, M., Vilanova, S., Gramazio, P., Sestras, A. F., Prohens, J., & Sestras, R. E. (2023). Comparative Analysis of Phenotypic and Molecular Data on Response to Main Pear Diseases and Pest Attack in a Germplasm Collection. International Journal of Molecular Sciences, 24(7), 6239. https://doi.org/10.3390/ijms24076239