

A Novel RT-LAMP for the Detection of Different Genotypes of Crimean–Congo Haemorrhagic Fever Virus in Patients from Spain

 , ,
, ,  , , , ,
, , , ,
Abstract
1. Introduction
2. Results
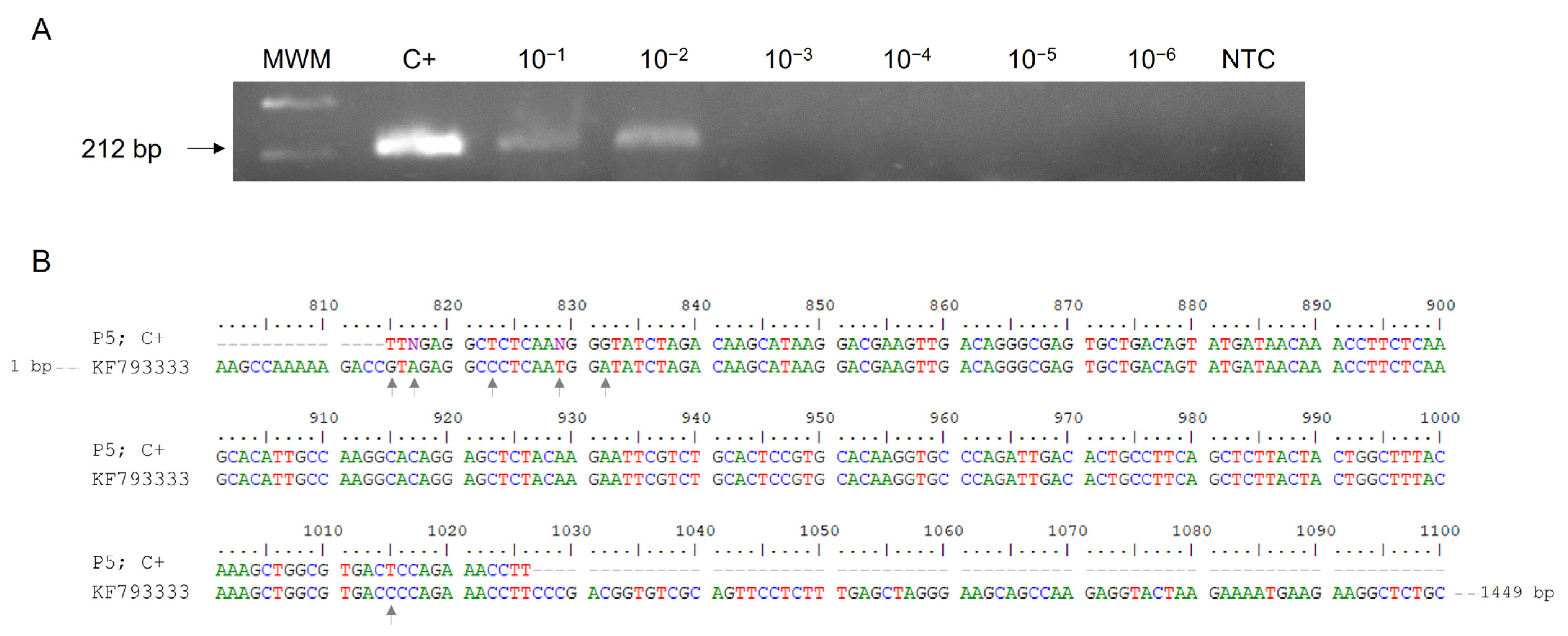
2.1. RT-PCR F3-B3 Verification of Target and Sensitivity
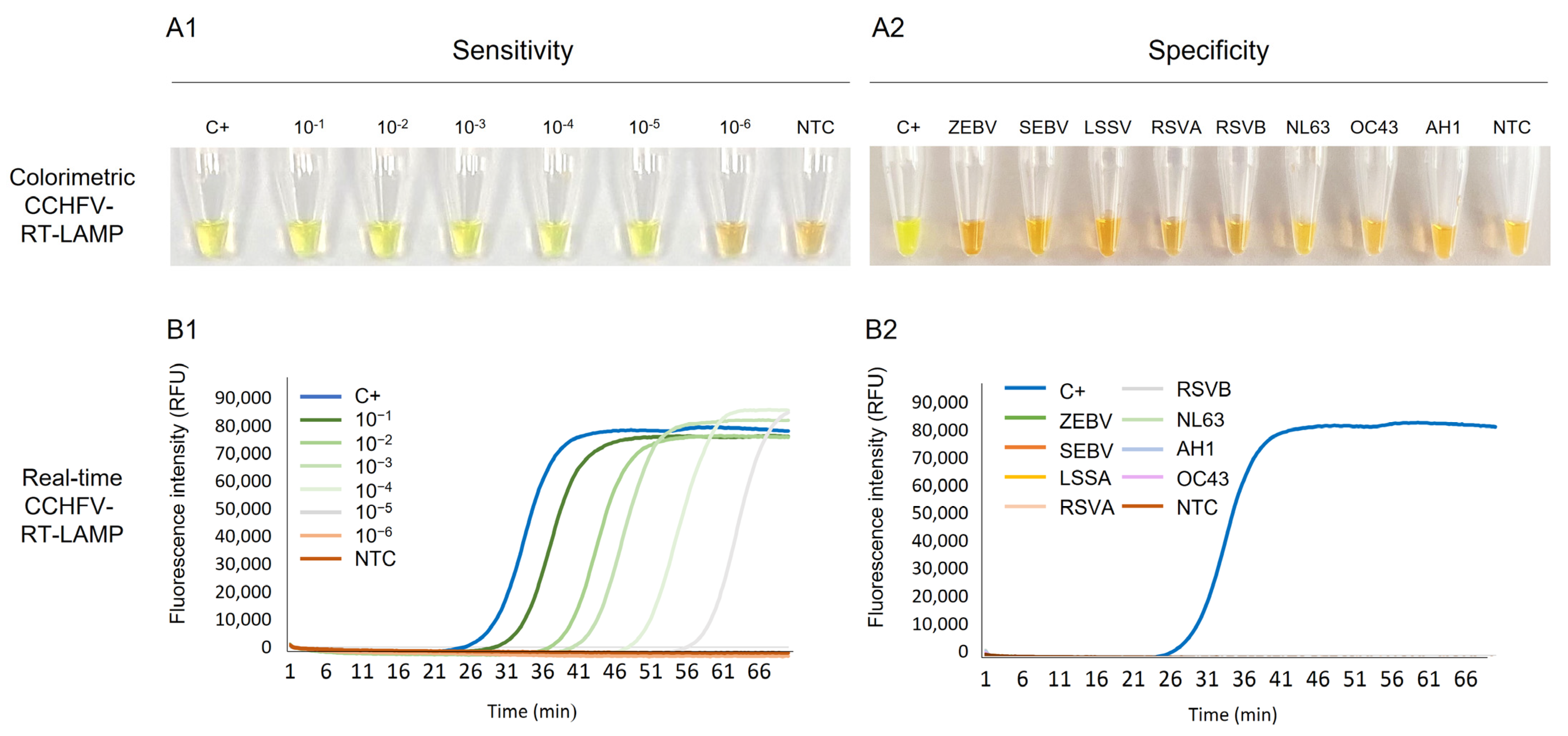
2.2. Establishing the RT-LAMP Assay: Sensitivity and Specificity
2.3. Clinical Sample Testing by CCHFV-RT-LAMP
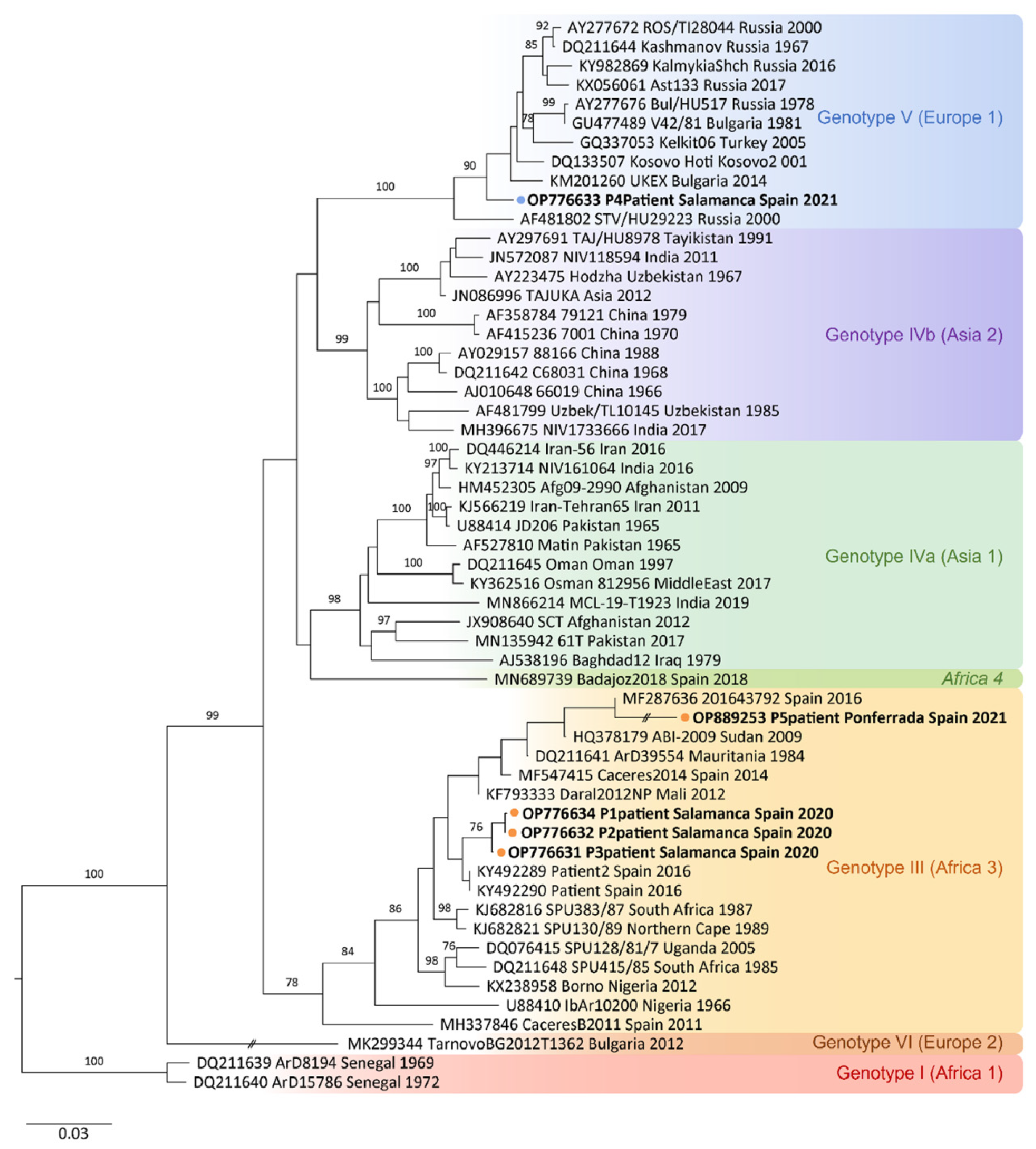
2.4. DNA Sequencing and Phylogenetic Analysis
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. CCHFV RNA-Positive Control and Patients’ RNA Samples
4.3. Design of Reverse-Transcription Loop-Mediated Isothermal Amplification Assay (RT-LAMP) for CCHFV
Sequence Selection and Primer Design for CCHFV-RT-LAMP

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin (n) | Genbank Accession | Strain/Isolate | Location, Isolation Year | Source of Isolate | Reference |
|---|---|---|---|---|---|
| African sequences (n = 11) | DQ076415 | SPU128/81/7 | Uganda (Semunya) | Tick * | [55] |
| DQ211639 | ArD8194 | Senegal, 1969 | H. truncatum | [59] | |
| DQ211640 | ArD15786 | Senegal, 1972 | Goat | [59] | |
| DQ211641 | ArD39554 | Mauritania, 1984 | H. marginatum | [59] | |
| DQ211648 | SPU415/85 | South Africa, 1985 | Human | [59] | |
| HQ378179 | AB1-2009 | Sudan (Abyei), 2009 | Human | [23] | |
| KF793333 | Daral 2012 NP | Mali (Daral), 2012 | Hyalomma | [77] | |
| KJ682816 | SPU383/87 | South Africa, 1987 | Human | [78] | |
| KJ682821 | SPU130/89 | Northern Cape, 1989 | Human | [78] | |
| KX238958 | NA | Nigeria (Borno), 2012 | Human | NA | |
| U88410 | IbAr10200 | Nigeria, 1966 | NA | NA | |
| Asian Sequences (n = 24) | AF358784 | 79121 | China, 1979 | NA | NA |
| AF415236 | 7001 | China, 1970 | NA | NA | |
| AF481799 | Uzbek/TI10145 | Uzbekistan, 1985 | Human | [79] | |
| AF527810 | Matin | Pakistan, 1965 | NA | NA | |
| AJ010648 | 66019 | China, 1966 | NA | [80] | |
| AJ538196 | Baghdad-12 | Iraq, 1979 | Human | [80] | |
| AY029157 | 88166 | China, 1988 | NA | NA | |
| AY223475 | Hodzha | Uzbekistan, 1967 | Human | [80] | |
| AY297691 | TAJ/HU8978 | Tajikistan, 1991 | Human | [81] | |
| DQ211642 | C-68031 | China, 1968 | Sheep | [59] | |
| DQ211645 | Oman | Oman, 1997 | Human | [59] | |
| DQ446214 | Iran-56 | Iran, 2017 | Human | NA | |
| GQ337053 | Turkey-Kelkit06 | Turkey, 2005 | Human | [82] | |
| HM452305 | Afg09-2990 | Afghanistan, 2009 | Human | [83] | |
| JN086996 | TAJUK | Asia, 2012 | NA | [28] | |
| JN572087 | NIV118594 | India, 2011 | H. antolicum | [84] | |
| JX908640 | SCT | Afghanistan, 2012 | Human | [85] | |
| KJ566219 | Iran-Tehran65 | Iran, 2011 | Human | [86] | |
| KY213714 | NIV161064 | India, 2016 | Human | [87] | |
| KY362516 | Oman 812956 | Middle Eastern, 2017 | Human | NA | |
| MH396675 | NIV1733666 | India | Human | NA | |
| MN135942 | 61T/Pakistan | Pakistan, 2017 | Tick | [88] | |
| U88414 | JD 206 | Pakistan, 1965 | NA | NA | |
| MN866214 | MCL-19-T-1923 | India, 2019 | Human | [89] | |
| European sequences (n = 16) | MH337846 | Cáceres/B 2011 | Spain, 2011 | H. lusitanicum | [13] |
| KX056061 | Ast133 | Russia, 2017 | Human | NA | |
| KY982869 | Kalmykia_Shch_2_2016 | Russia, 2016 | Human | [90] | |
| MK299344 | Malko Tarnovo-BG2012-T1362 | Bulgaria, 2012 | R. bursa | [91] | |
| MN689739 | Badajoz 2018 | Spain, 2018 | Human | NA | |
| MF547415 | Cáceres 2014 | Spain, 2014 | H. lusitanicum | [12] | |
| DQ133507 | Kosovo Hoti | Kosovo, 2001 | Human | [26] | |
| KM201260 | UK ex | Bulgaria, 2014 | Human | [92] | |
| GU477489 | V42/81 | Bulgaria, 1981 | Human | [93] | |
| AY277676 | NA | Russia | Human | NA | |
| AY277672 | ROS/TI28044 | Russia, 2000 | H. marginatum | NA | |
| DQ211644 | Kashmanov | Russia, 1967 | Human | [59] | |
| AF481802 | STV/HU29223 | Russia, 2000 | Human | [79] | |
| KY492290 | patient1 | Spain, 2016 | Human | [10] | |
| KY492289 | patient2 | Spain, 2016 | Human | [10] | |
| MF287636 | 201643792 | Spain, 2016 | Human | [8] |
4.4. RT-PCR Using Outer Primers F3 and B3
4.5. Establishing the RT-LAMP Assay for CCHFV Detection
4.5.1. Conventional Colorimetric CCHFV-RT-LAMP
4.5.2. Real-Time CCHFV-RT-LAMP
4.5.3. Sensitivity and Specificity of CCHFV-RT-LAMP
4.5.4. Clinical Samples Testing by CCHFV-RT-LAMP
4.6. DNA Sequencing
4.7. Phylogenetic Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Portillo, A.; Palomar, A.M.; Santibáñez, P.; Oteo, J.A. Epidemiological aspects of crimean-congo hemorrhagic fever in western europe: What about the future? Microorganisms 2021, 9, 649. [Google Scholar] [CrossRef]
- Bente, D.A.; Forrester, N.L.; Watts, D.M.; McAuley, A.J.; Whitehouse, C.A.; Bray, M. Crimean-Congo hemorrhagic fever: History, epidemiology, pathogenesis, clinical syndrome and genetic diversity. Antiviral Res. 2013, 100, 159–189. [Google Scholar] [CrossRef]
- Papa, A.; Tsergouli, K.; Tsioka, K.; Mirazimi, A. Crimean-Congo hemorrhagic fever: Tick-host-virus interactions. Front. Cell. Infect. Microbiol. 2017, 7, 213. [Google Scholar] [CrossRef]
- Tsergouli, K.; Karampatakis, T.; Haidich, A.B.; Metallidis, S.; Papa, A. Nosocomial infections caused by Crimean–Congo haemorrhagic fever virus. J. Hosp. Infect. 2020, 105, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Pshenichnaya, N.Y.; Nenadskaya, S.A. Probable Crimean-Congo hemorrhagic fever virus transmission occurred after aerosol-generating medical procedures in Russia: Nosocomial cluster. Int. J. Infect. Dis. 2015, 33, 120–122. [Google Scholar] [CrossRef] [PubMed]
- Shahhosseini, N.; Wong, G.; Babuadze, G.; Camp, J.V.; Ergonul, O.; Kobinger, G.P.; Chinikar, S.; Nowotny, N. Crimean-congo hemorrhagic fever virus in asia, africa and europe. Microorganisms 2021, 9, 1907. [Google Scholar] [CrossRef] [PubMed]
- Farooq, H.; Beveridge, N.; Fletcher, T.; Ghani, E.; Jamil, B.; Hasan, Z.; Ikram, A.; Safdar, R.M.; Salman, M.; Umair, M.; et al. A systematic review on the incidence and mortality of Crimean-Congo Haemorrhagic Fever (CCHF) in Pakistan. Int. J. Infect. Dis. 2020, 101, 250. [Google Scholar] [CrossRef]
- Arellano, E.R.; Hernández, L.; Goyanes, M.J.; Arsuaga, M.; Cruz, A.F.; Negredo, A.; Sánchez-Seco, M.P. Phylogenetic Characterization of Crimean-Congo Hemorrhagic Fever Virus, Spain. Emerg. Infect. Dis. 2017, 23, 2078–2080. [Google Scholar] [CrossRef]
- Carroll, S.A.; Bird, B.H.; Rollin, P.E.; Nichol, S.T. Ancient common ancestry of Crimean-Congo hemorrhagic fever virus. Mol. Phylogenet. Evol. 2010, 55, 1103–1110. [Google Scholar] [CrossRef] [PubMed]
- Negredo, A.; de la Calle-Prieto, F.; Palencia-Herrejón, E.; Mora-Rillo, M.; Astray-Mochales, J.; Sánchez-Seco, M.P.; Bermejo Lopez, E.; Menárguez, J.; Fernández-Cruz, A.; Sánchez-Artola, B.; et al. Autochthonous Crimean–Congo Hemorrhagic Fever in Spain. N. Engl. J. Med. 2017, 377, 154–161. [Google Scholar] [CrossRef] [PubMed]
- Estrada-Peña, A.; Palomar, A.M.; Santibáñez, P.; Sánchez, N.; Habela, M.A.; Portillo, A.; Romero, L.; Oteo, J.A. Crimean-Congo Hemorrhagic Fever Virus in Ticks, Southwestern Europe, 2010. BMC Infect. Dis. 2012, 18, 179. [Google Scholar] [CrossRef] [PubMed]
- Cajimat, M.N.B.; Rodriguez, S.E.; Schuster, I.U.E.; Swetnam, D.M.; Ksiazek, T.G.; Habela, M.A.; Negredo, A.I.; Estrada-Peña, A.; Barrett, A.D.T.; Bente, D.A. Genomic Characterization of Crimean-Congo Hemorrhagic Fever Virus in Hyalomma Tick from Spain, 2014. Vector-Borne Zoonotic Dis. 2017, 17, 714–719. [Google Scholar] [CrossRef] [PubMed]
- Negredo, A.; Habela, M.Á.; Ramírez de Arellano, E.; Diez, F.; Lasala, F.; López, P.; Sarriá, A.; Labiod, N.; Calero-Bernal, R.; Arenas, M.; et al. Survey of Crimean-Congo Hemorrhagic Fever Enzootic Focus, Spain, 2011–2015. Emerg. Infect. Dis. 2019, 25, 1177–1184. [Google Scholar] [CrossRef] [PubMed]
- Moraga-Fernández, A.; Ruiz-Fons, F.; Habela, M.A.; Royo-Hernández, L.; Calero-Bernal, R.; Gortazar, C.; de la Fuente, J.; Fernández de Mera, I.G. Detection of new Crimean–Congo haemorrhagic fever virus genotypes in ticks feeding on deer and wild boar, Spain. Transbound. Emerg. Dis. 2020, 1944, 993–1000. [Google Scholar] [CrossRef]
- Sánchez-Seco, M.P.; Sierra, M.J.; Estrada-Peña, A.; Valcárcel, F.; Molina, R.; De Arellano, E.R.; Olmeda, A.S.; Miguel, L.G.S.; Jiménez, M.; Romero, L.J.; et al. Widespread Detection of Multiple Strains of Crimean-Congo Hemorrhagic Fever Virus in Ticks, Spain. Emerg. Infect. Dis. 2022, 28, 394–402. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Annual Review of Diseases Prioritized under the Research and Development Blueprint. Meeting Report. 2017. Available online: https://www.who.int/blueprint/what/research-development/2017-Prioritization-Long-Report.pdf?ua=1 (accessed on 23 July 2022).
- Negredo, A.; Sánchez-Ledesma, M.; Llorente, F.; Pérez-Olmeda, M.; Belhassen-García, M.; González-Calle, D.; Sánchez-Seco, M.P.; Jiménez-Clavero, M.Á. Retrospective Identification of Early Autochthonous Case of Crimean-Congo Hemorrhagic Fever, Spain, 2013. Emerg. Infect. Dis. 2021, 27, 1754–1756. [Google Scholar] [CrossRef]
- (ECDC), E.C. for D.C. Communicable Disease Threats Report. 2022. Available online: https://www.ecdc.europa.eu/sites/default/files/documents/Communicable-disease-threats-report-13-aug-2022-all-users.pdf (accessed on 13 February 2022).
- Negredo, A.; Sánchez-Arroyo, R.; Díez-Fuertes, F.; De Ory, F.; Budiño, M.A.; Vázquez, A.; Garcinuño, Á.; Hernández, L.; de la Hoz González, C.; Gutiérrez-Arroyo, A.; et al. Fatal case of crimean-congo hemorrhagic fever caused by reassortant virus, Spain, 2018. Emerg. Infect. Dis. 2021, 27, 1211–1215. [Google Scholar] [CrossRef]
- Monsalve Arteaga, L.; Muñoz Bellido, J.L.; Negredo, A.I.; García Criado, J.; Vieira Lista, M.C.; Sánchez Serrano, J.Á.; Vicente Santiago, M.B.; López Bernús, A.; de Ory Manchón, F.; Sánchez Seco, M.P.; et al. New circulation of genotype V of Crimean-Congo haemorrhagic fever virus in humans from Spain. PLoS Negl. Trop. Dis. 2021, 15, e0009197. [Google Scholar] [CrossRef]
- Bartolini, B.; Gruber, C.E.M.; Koopmans, M.; Avšič, T.; Bino, S.; Christova, I.; Grunow, R.; Hewson, R.; Korukluoglu, G.; Lemos, C.M.; et al. Laboratory management of Crimean-Congo haemorrhagic fever virus infections: Perspectives from two European networks. Eurosurveillance 2019, 24, 1800093. [Google Scholar] [CrossRef]
- Raabe, V.N. Diagnostic Testing for Crimean-Congo Hemorrhagic Fever. J. Clin. Microbiol. 2020, 58, e01580-19. [Google Scholar] [CrossRef]
- Aradaib, I.E.; Erickson, B.R.; Karsany, M.S.; Khristova, M.L.; Elageb, R.M.; Mohamed, M.E.H.; Nichol, S.T. Multiple Crimean-Congo hemorrhagic fever virus strains are associated with disease outbreaks in Sudan, 2008–2009. PLoS Negl. Trop. Dis. 2011, 5, e1159. [Google Scholar] [CrossRef] [PubMed]
- Drosten, C.; Göttig, S.; Schilling, S.; Asper, M.; Panning, M.; Schmitz, H.; Günther, S. Rapid detection and quantification of RNA of Ebola and Marburg viruses, Lassa virus, Crimean-Congo hemorrhagic fever virus, Rift Valley fever virus, dengue virus, and yellow fever virus by real-time reverse transcription-PCR. J. Clin. Microbiol. 2002, 40, 2323–2330. [Google Scholar] [CrossRef] [PubMed]
- Yapar, M.; Aydogan, H.; Pahsa, A.; Besirbellioglu, B.A.; Bodour, H.; Basustaoglu, A.C.; Guney, C.; Avci, I.Y.; Sener, K.; Setteh, M.H.A.; et al. Rapid and quantitative detection of Crimean-Congo hemorrhagic fever virus by one-step real-time reverse transcriptase-PCR. Jpn. J. Infect. Dis. 2005, 58, 358–362. [Google Scholar] [PubMed]
- Duh, D.; Saksida, A.; Petrovec, M.; Dedushaj, I.; Avšič-ŽUpanc, T. Novel one-step real-time RT-PCR assay for rapid and specific diagnosis of Crimean-Congo hemorrhagic fever encountered in the Balkans. J. Virol. Methods 2006, 133, 175–179. [Google Scholar] [CrossRef] [PubMed]
- Wölfel, R.; Paweska, J.T.; Petersen, N.; Grobbelaar, A.A.; Leman, P.A.; Hewson, R.; Georges-Courbot, M.C.; Papa, A.; Günther, S.; Drosten, C. Virus detection and monitoring of viral load in Crimean-Congo hemorrhagic fever virus patients. Emerg. Infect. Dis. 2007, 13, 1097–1100. [Google Scholar] [CrossRef]
- Atkinson, B.; Chamberlain, J.; Logue, C.H.; Cook, N.; Bruce, C.; Dowall, S.D.; Hewson, R. Development of a real-time RT-PCR assay for the detection of Crimean-Congo hemorrhagic fever virus. Vector-Borne Zoonotic Dis. 2012, 12, 786–793. [Google Scholar] [CrossRef]
- Zahraei, B.H.; Zadeh, M.S.H.; Asl, M.N.; Yeganeh, S.Z.; Tat, M.; Metanat, M.; Rad, N.S.; Nejad, B.K.; Zafari, E.; Sharti, M.; et al. Novel, in-house, sybr green based one-step rRT-PCR: Rapid and accurate diagnosis of crimean-congo hemorrhagic fever virus in suspected patients from Iran. Jundishapur J. Microbiol. 2015, 9, 2–6. [Google Scholar] [CrossRef]
- Koehler, J.W.; Delp, K.L.; Hall, A.T.; Olschner, S.P.; Kearney, B.J.; Garrison, A.R.; Altamura, L.A.; Rossi, C.A.; Minogue, T.D. Sequence optimized real-time reverse transcription polymerase chain reaction assay for detection of crimean-congo hemorrhagic fever virus. Am. J. Trop. Med. Hyg. 2018, 98, 211–215. [Google Scholar] [CrossRef]
- Sas, M.A.; Vina-Rodriguez, A.; Mertens, M.; Eiden, M.; Emmerich, P.; Chaintoutis, S.C.; Mirazimi, A.; Groschup, M.H. A one-step multiplex real-time RT-PCR for the universal detection of all currently known CCHFV genotypes. J. Virol. Methods 2018, 255, 38–43. [Google Scholar] [CrossRef]
- Papa, A. Diagnostic approaches for crimean-congo hemorrhagic fever virus. Expert Rev. Mol. Diagn. 2019, 19, 531–536. [Google Scholar] [CrossRef]
- Notomi, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, e63. [Google Scholar] [CrossRef] [PubMed]
- Niemz, A.; Ferguson, T.M.; Boyle, D.S. Point-of-care nucleic acid testing for infectious diseases. Trends Biotechnol. 2011, 29, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Moehling, T.J.; Choi, G.; Dugan, L.C.; Salit, M.; Meagher, R.J. LAMP Diagnostics at the Point-of-Care: Emerging Trends and Perspectives for the Developer Community. Expert Rev. Mol. Diagn. 2021, 21, 43–61. [Google Scholar] [CrossRef] [PubMed]
- Parida, M.; Posadas, G.; Inoue, S.; Hasebe, F.; Morita, K. Real-Time Reverse Transcription Loop-Mediated Isothermal Amplification for Rapid Detection of West Nile Virus. J. Clin. Microbiol. 2004, 42, 257–263. [Google Scholar] [CrossRef] [PubMed]
- Parida, M.; Shukla, J.; Sharma, S.; Santhosh, S.R.; Ravi, V.; Mani, R.; Thomas, M.; Khare, S.; Rai, A.; Ratho, R.K.; et al. Development and evaluation of reverse transcription loop-mediated isothermal amplification assay for rapid and real-time detection of the swine-origin influenza a H1N1 virus. J. Mol. Diagn. 2011, 13, 100–107. [Google Scholar] [CrossRef]
- Parida, M.M.; Santhosh, S.R.; Dash, P.K.; Tripathi, N.K.; Lakshmi, V.; Mamidi, N.; Shrivastva, A.; Gupta, N.; Saxena, P.; Pradeep Babu, J.; et al. Rapid and real-time detection of Chikungunya virus by reverse transcription loop-mediated isothermal amplification assay. J. Clin. Microbiol. 2007, 45, 351–357. [Google Scholar] [CrossRef]
- Lopez-Jimena, B.; Wehner, S.; Harold, G.; Bakheit, M.; Frischmann, S.; Bekaert, M.; Faye, O.; Sall, A.A.; Weidmann, M. Development of a single-tube one-step RT-LAMP assay to detect the Chikungunya virus genome. PLoS Negl. Trop. Dis. 2018, 12, e0006448. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Wang, H.; Wang, L.; Li, L.; Jin, H.; Xu, C.; Feng, N.; Wang, J.; Li, Q.; Zhao, Y.; et al. Visual detection of West Nile virus using reverse transcription loop-mediated isothermal amplification combined with a vertical flow visualization strip. Front. Microbiol. 2016, 7, 554. [Google Scholar] [CrossRef]
- Kumar, J.S.; Saxena, D.; Parida, M.; Rathinam, S. Evaluation of real-time reverse-transcription loop-mediated isothermal amplification assay for clinical diagnosis of west Nile virus in patients. Indian J. Med. Res. 2018, 147, 293–298. [Google Scholar] [CrossRef]
- Liu, H.; Liu, Z.J.; Jing, J.; Ren, J.Q.; Liu, Y.Y.; Guo, H.H.; Fan, M.; Lu, H.J.; Jin, N.Y. Reverse transcription loop-mediated isothermal amplification for rapid detection of Japanese encephalitis virus in swine and mosquitoes. Vector-Borne Zoonotic Dis. 2012, 12, 1042–1052. [Google Scholar] [CrossRef]
- Parida, M.; Horioke, K.; Ishida, H.; Dash, P.K.; Saxena, P.; Jana, A.M.; Islam, M.A.; Inoue, S.; Hosaka, N.; Morita, K. Rapid detection and differentiation of dengue virus serotypes by a real-time reverse transcription-loop-mediated isothermal amplification assay. J. Clin. Microbiol. 2005, 43, 2895–2903. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.F.; Li, M.; Zhong, L.L.; Lu, S.M.; Liu, Z.X.; Pu, J.Y.; Wen, J.S.; Xi, H. Development of reverse-transcription loop-mediated isothermal amplification assay for rapid detection and differentiation of dengue virus serotypes 1-4. BMC Microbiol. 2015, 15, 265. [Google Scholar] [CrossRef] [PubMed]
- Lau, Y.L.; Lai, M.Y.; Teoh, B.T.; Abd-Jamil, J.; Johari, J.; Sam, S.S.; Tan, K.K.; AbuBakar, S. Colorimetric detection of dengue by single tube reverse-transcription-loop-mediated isothermal amplification. PLoS ONE 2015, 10, e0138694. [Google Scholar] [CrossRef]
- Oloniniyi, O.K.; Kurosaki, Y.; Miyamoto, H.; Takada, A.; Yasuda, J. Rapid detection of all known ebolavirus species by reverse transcription-loop-mediated isothermal amplification (RT-LAMP). J. Virol. Methods 2017, 246, 8–14. [Google Scholar] [CrossRef]
- Bonney, L.C.; Watson, R.J.; Slack, G.S.; Bosworth, A.; Vasileva Wand, N.I.; Hewson, R. A flexible format lamp assay for rapid detection of ebola virus. PLoS Negl. Trop. Dis. 2020, 14, e0008496. [Google Scholar] [CrossRef]
- Peyrefitte, C.N.; Boubis, L.; Coudrier, D.; Bouloy, M.; Grandadam, M.; Tolou, H.J.; Plumet, S. Real-time reverse-transcription loop-mediated isothermal amplification for rapid detection of Rift Valley fever virus. J. Clin. Microbiol. 2008, 46, 3653–3659. [Google Scholar] [CrossRef]
- Le Roux, C.A.; Kubo, T.; Grobbelaar, A.A.; Van Vuren, P.J.; Weyer, J.; Nel, L.H.; Swanepoel, R.; Morita, K.; Paweska, J.T. Development and evaluation of a real-time reverse transcription-loop- mediated isothermal amplification assay for rapid detection of rift valley fever virus in clinical specimens. J. Clin. Microbiol. 2009, 47, 645–651. [Google Scholar] [CrossRef] [PubMed]
- Han, Q.; Zhang, S.; Liu, D.; Yan, F.; Wang, H.; Huang, P.; Bi, J.; Jin, H.; Feng, N.; Cao, Z.; et al. Development of a Visible Reverse Transcription-Loop-Mediated Isothermal Amplification Assay for the Detection of Rift Valley Fever Virus. Front. Microbiol. 2020, 11, 590732. [Google Scholar] [CrossRef] [PubMed]
- ole Kwallah, A.; Inoue, S.; Muigai, A.W.T.; Kubo, T.; Sang, R.; Morita, K.; Mwau, M. A real-time reverse transcription loop-mediated isothermal amplification assay for the rapid detection of yellow fever virus. J. Virol. Methods 2013, 193, 23–27. [Google Scholar] [CrossRef]
- Osman, H.A.M.; Eltom, K.H.; Musa, N.O.; Bilal, N.M.; Elbashir, M.I.; Aradaib, I.E. Development and evaluation of loop-mediated isothermal amplification assay for detection of Crimean Congo hemorrhagic fever virus in Sudan. J. Virol. Methods 2013, 190, 4–10. [Google Scholar] [CrossRef]
- Ternovoi, V.A.; Kononova, Y.V.; Zaykovskaya, A.V.; Chub, E.V.; Volynkina, A.S.; Mikryukova, T.P.; Kotenev, E.S.; Pyankov, O.V.; Sementsova, A.O.; Loktev, V.B. Development and assessment of a reagent kit for rna detection of crimean-congo hemorrhagic fever virus with using reverse transcription loop-mediated isothermal amplification method. Russ. Clin. Lab. Diagnostics 2019, 64, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Kumar, J.S.; Parida, M.; Shete, A.M.; Majumdar, T.; Patil, S.; Yadav, P.D.; Dash, P.K. Development of a Reverse Transcription Loop-Mediated Isothermal Amplification [RT-LAMP] as a early rapid detection assay for Crimean Congo Hemorrhagic Fever virus. Acta Trop. 2022, 231, 106435. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, J.; Cook, N.; Lloyd, G.; Mioulet, V.; Tolley, H.; Hewson, R. Co-evolutionary patterns of variation in small and large RNA segments of Crimean-Congo hemorrhagic fever virus. J. Gen. Virol. 2005, 86, 3337–3341. [Google Scholar] [CrossRef] [PubMed]
- Nasirian, H. New aspects about Crimean-Congo hemorrhagic fever (CCHF) cases and associated fatality trends: A global systematic review and meta-analysis. Comp. Immunol. Microbiol. Infect. Dis. 2020, 69, 101429. [Google Scholar] [CrossRef]
- Tezer, H.; Polat, M. Diagnosis of Crimean-Congo hemorrhagic fever. Expert Rev. Anti. Infect. Ther. 2015, 13, 555–566. [Google Scholar] [CrossRef]
- Mazzola, L.T.; Kelly-Cirino, C. Diagnostic tests for Crimean-Congo haemorrhagic fever: A widespread tickborne disease. BMJ Glob. Health 2019, 4, e001114. [Google Scholar] [CrossRef]
- Deyde, V.M.; Khristova, M.L.; Rollin, P.E.; Ksiazek, T.G.; Nichol, S.T. Crimean-Congo Hemorrhagic Fever Virus Genomics and Global Diversity. J. Virol. 2006, 80, 8834–8842. [Google Scholar] [CrossRef]
- Parida, M.; Sannarangaiah, S.; Dash, P.K.; Rao, P.V.L.; Morita, K. Loop mediated isothermal amplification (LAMP): A new generation of innovative gene amplification technique; perspectives in clinical diagnosis of infectious diseases. Rev. Med. Virol. 2008, 18, 407–421. [Google Scholar] [CrossRef]
- Sanjuán, R.; Nebot, M.R.; Chirico, N.; Mansky, L.M.; Belshaw, R. Viral Mutation Rates. J. Virol. 2010, 84, 9733–9748. [Google Scholar] [CrossRef]
- Domingo, E.; Perales, C. Quasispecies and virus. Eur. Biophys. J. 2018, 47, 443–457. [Google Scholar] [CrossRef]
- Domingo, E.; Perales, C. Viral quasispecies. PLoS Genet. 2019, 15, e1008271. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Wan, Z.; Yang, S.; Li, Y.; Li, M.; Wang, B.; Hu, Y.; Xia, X.; Jin, X.; Yu, N.; et al. A mismatch-tolerant reverse transcription loop-mediated isothermal amplification method and its application on simultaneous detection of all four serotype of dengue viruses. Front. Microbiol. 2019, 10, 1056. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, Y.; Ma, Y.; Xu, R.; Jin, X.; Zhang, C. A Mismatch-tolerant RT-LAMP Method for Molecular Diagnosis of Highly Variable Viruses. Bio-Protocol. 2019, 9, e3415. [Google Scholar] [CrossRef] [PubMed]
- Waters, R.A.; Fowler, V.L.; Armson, B.; Nelson, N.; Gloster, J.; Paton, D.J.; King, D.P. Preliminary validation of direct detection of foot-and-mouth disease virus within clinical samples using reverse transcription Loop-mediated isothermal amplification coupled with a simple lateral flow device for detection. PLoS ONE 2014, 9, e105630. [Google Scholar] [CrossRef] [PubMed]
- Hasanoglu, I.; Guner, R.; Carhan, A.; Tufan, Z.K.; Caglayik, D.Y.; Yilmaz, G.R.; Tasyaran, M.A. Dynamics of viral load in Crimean Congo hemorrhagic fever. J. Med. Virol. 2018, 90, 639–643. [Google Scholar] [CrossRef]
- Saksida, A.; Duh, D.; Wraber, B.; Dedushaj, I.; Ahmeti, S.; Avšič-Županc, T. Interacting roles of immune mechanisms and viral load in the pathogenesis of Crimean-Congo hemorrhagic fever. Clin. Vaccine Immunol. 2010, 17, 1086–1093. [Google Scholar] [CrossRef]
- García-Bernalt Diego, J.G.; Fernández-Soto, P.; Domínguez-Gil, M.; Belhassen-García, M.; Bellido, J.L.M.; Muro, A. A Simple, Affordable, Rapid, Stabilized, Colorimetric, Versatile RT-LAMP Assay to Detect SARS-CoV-2. Diagnostics 2021, 11, 438. [Google Scholar] [CrossRef]
- Palomar, A.M.; Portillo, A.; Santibáñez, P.; Mazuelas, D.; Arizaga, J.; Crespo, A.; Gutiérrez, Ó.; Cuadrado, J.F.; Oteo, J.A. Crimean-congo hemorrhagic fever virus in ticks from migratory birds, Morocco. Emerg. Infect. Dis. 2013, 19, 260–263. [Google Scholar] [CrossRef]
- Thomas, S.; Thomson, G.; Dowall, S.; Bruce, C.; Cook, N.; Easterbrook, L.; O’Donoghue, L.; Summers, S.; Ajazaj, L.; Hewson, R.; et al. Review of Crimean Congo hemorrhagic fever infection in kosova in 2008 and 2009: Prolonged viremias and virus detected in urine by PCR. Vector-Borne Zoonotic Dis. 2012, 12, 800–804. [Google Scholar] [CrossRef]
- Yagci-Caglayik, D.; Kayaaslan, B.; Yapar, D.; Kocagul-Celikbas, A.; Ozkaya-Parlakay, A.; Emek, M.; Baykam, N.; Tezer, H.; Korukluoglu, G.; Ozkul, A. Monitoring Crimean-Congo haemorrhagic fever virus RNA shedding in body secretions and serological status in hospitalised patients, Turkey, 2015. Eurosurveillance 2020, 25, 1900284. [Google Scholar] [CrossRef]
- Bodur, H.; Akinci, E.; Öngürü, P.; Carhan, A.; Uyar, Y.; Tanrici, A.; Cataloluk, O.; Kubar, A. Detection of Crimean-Congo hemorrhagic fever virus genome in saliva and urine. Int. J. Infect. Dis. 2010, 14, 2009–2011. [Google Scholar] [CrossRef] [PubMed]
- Thompson, J.D.; Higgins, D.G.; Gibson, T.J. CLUSTAL W: Improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 1994, 22, 4673–4680. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Eiken Chemical Co., Ltd. A Guide to LAMP primer designing (PrimerExplorer V5). PrimerExplorer V5 2019, 1–18. Available online: https://primerexplorer.jp/e/v5_manual/pdf/PrimerExplorerV5_Manual_1.pdf (accessed on 20 January 2022).
- Zivcec, M.; Maïga, O.; Kelly, A.; Feldmann, F.; Sogoba, N.; Schwan, T.G.; Feldmann, H.; Safronetz, D. Unique strain of crimean-congo hemorrhagic fever virus, Mali. Emerg. Infect. Dis. 2014, 20, 911–913. [Google Scholar] [CrossRef] [PubMed]
- Goedhals, D.; Bester, P.A.; Paweska, J.T.; Swanepoel, R.; Burt, F.J. Next-generation sequencing of southern African Crimean-Congo haemorrhagic fever virus isolates reveals a high frequency of M segment reassortment. Epidemiol. Infect. 2014, 142, 1952–1962. [Google Scholar] [CrossRef]
- Petrova, I.D.; Seregin, S.V.; Petrov, V.S.; Vyshemirski, O.I.; Kuzina, I.I.; L’vov, D.K.; Samokhvalov, E.I.; Tyunikov, G.I.; Gutorov, V.V.; Yashina, L.N.; et al. Genetic characteristics of the S-segment of RNA from two strains of the Crimean-Congo hemorrhagic fever virus isolated in the south of Russia and in Uzbekistan. Vopr Virusol 2003, 48, 8–11. [Google Scholar]
- Hewson, R.; Gmyl, A.; Gmyl, L.; Smirnova, S.E.; Karganova, G.; Jamil, B.; Hasan, R.; Chamberlain, J.; Clegg, C. Evidence of segment reassortment in Crimean-Congo haemorrhagic fever virus. J. Gen. Virol. 2004, 85, 3059–3070. [Google Scholar] [CrossRef]
- Seregin, S.V.; Tumanova, I.Y.; Vyshemirski, O.I.; Petrova, I.D.; Lvov, D.K.; Gromashevski, V.L.; Samokhvalov, E.I.; Tiunnikov, G.I.; Gutorov, V.V.; Tishkova, F.H.; et al. Study of the genetic variability of Crimean-Congo hemorrhagic fever virus in Central Asia. Dokl. Biochem. Biophys. 2004, 398, 313–315. [Google Scholar] [CrossRef]
- Özdarendeli, A.; Çanakoǧlu, N.; Berber, E.; Aydin, K.; Tonbak, Ş.; Ertek, M.; Buzgan, T.; Bolat, Y.; Aktaş, M.; Kalkan, A. The complete genome analysis of Crimean-Congo hemorrhagic fever virus isolated in Turkey. Virus Res. 2010, 147, 288–293. [Google Scholar] [CrossRef]
- Ölschläger, S.; Gabriel, M.; Schmidt-Chanasit, J.; Meyer, M.; Osborn, E.; Conger, N.G.; Allan, P.F.; Günther, S. Complete sequence and phylogenetic characterisation of Crimean-Congo hemorrhagic fever virus from Afghanistan. J. Clin. Virol. 2011, 50, 90–92. [Google Scholar] [CrossRef] [PubMed]
- Yadav, P.D.; Cherian, S.S.; Zawar, D.; Kokate, P.; Gunjikar, R.; Jadhav, S.; Mishra, A.C.; Mourya, D.T. Genetic characterization and molecular clock analyses of the Crimean-Congo hemorrhagic fever virus from human and ticks in India, 2010-2011. Infect. Genet. Evol. 2013, 14, 223–231. [Google Scholar] [CrossRef] [PubMed]
- Chamberlain, J.; Atkinson, B.; Logue, C.H.; Latham, J.; Newman, E.N.C.; Hewson, R. Genome sequence of ex-Afghanistan Crimean-Congo hemorrhagic fever virus SCT strain, from an imported United Kingdom case in October 2012. Genome Announc. 2013, 1, 2012–2013. [Google Scholar] [CrossRef] [PubMed]
- Chinikar, S.; Shah-Hosseini, N.; Bouzari, S.; Shokrgozar, M.A.; Mostafavi, E.; Jalali, T.; Khakifirouz, S.; Groschup, M.H.; Niedrig, M. Assessment of recombination in the s-segment genome of crimean-congo hemorrhagic fever virus in iran. J. Arthropod. Borne. Dis. 2016, 10, 12–23. [Google Scholar]
- Yadav, P.D.; Thacker, S.; Patil, D.Y.; Jain, R.; Mourya, D.T. Crimean-congo hemorrhagic fever in migrant worker returning from Oman to India, 2016. Emerg. Infect. Dis. 2017, 23, 1005–1008. [Google Scholar] [CrossRef]
- Zohaib, A.; Saqib, M.; Athar, M.A.; Hussain, M.H.; Sial, A.; Tayyab, M.H.; Batool, M.; Sadia, H.; Taj, Z.; Tahir, U.; et al. Crimean-Congo Hemorrhagic Fever Virus in Humans and Livestock, Pakistan, 2015–2017. Emerg. Infect. Dis. 2020, 26, 773–777. [Google Scholar] [CrossRef]
- Sahay, R.R.; Dhandore, S.; Yadav, P.D.; Chauhan, A.; Bhatt, L.; Garg, V.; Gupta, N.; Nyayanit, D.A.; Shete, A.M.; Singh, R.; et al. Detection of African genotype in Hyalomma tick pools during Crimean Congo hemorrhagic fever outbreak, Rajasthan, India, 2019. Virus Res. 2020, 286, 198046. [Google Scholar] [CrossRef]
- Dedkov, V.G.; Shchelkanov, M.Y.; Bushkieva, B.T.; Rudenko, T.A.; Kurdyukova, O.V.; Galkina, I.V.; Sapotsky, M.V.; Blinova, E.A.; Dzhambinov, S.D.; Shipulin, G.A. A neonatal death associated with Crimean-Congo hemorrhagic fever (Republic of Kalmykia, Russia, June 2016). Antiviral Res. 2017, 146, 146–148. [Google Scholar] [CrossRef]
- Hua, B.L.; Scholte, F.E.M.; Ohlendorf, V.; Kopp, A.; Marklewitz, M.; Drosten, C.; Nichol, S.T.; Spiropoulou, C.F.; Junglen, S.; Bergeron, É. A single mutation in crimean-congo hemorrhagic fever virus discovered in ticks impairs infectivity in human cells. eLife 2020, 9, e50999. [Google Scholar] [CrossRef]
- Lumley, S.; Atkinson, B.; Dowall, S.D.; Pitman, J.K.; Staplehurst, S.; Busuttil, J.; Simpson, A.J.; Aarons, E.J.; Petridou, C.; Nijjar, N.; et al. Non-fatal case of Crimean-Congo haemorrhagic fever imported into the United Kingdom (ex Bulgaria), June 2014. Euro Surveill 2014, 19, 20864. [Google Scholar] [CrossRef]
- Papa, A.; Papadimitriou, E.; Christova, I. The Bulgarian vaccine Crimean-Congo haemorrhagic fever virus strain. Scand. J. Infect. Dis. 2011, 43, 225–229. [Google Scholar] [CrossRef] [PubMed]
- Hall, T.A. Bioedit: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]






| P1 | P2 | P3 | P4 | P5 (C+) | |
|---|---|---|---|---|---|
| Sex | Male | Male | Male | Male | Female |
| Age (years) | 70 | 54 | 69 | 59 | 30 |
| Date | June 2020 | July 2020 | August 2020 | April 2021 | June 2021 |
| Tick bite | Yes | Yes | Yes | Yes | Yes |
| Habitat | Rural | Rural | Rural | Rural | Rural |
| Fever | Yes | Yes | Yes | Yes | Yes |
| Any bleeding symptomatology | Yes | No | Yes | No | Yes |
| Exitus letalis | No | No | Yes | No | No |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Febrer-Sendra, B.; Fernández-Soto, P.; García-Bernalt Diego, J.; Crego-Vicente, B.; Negredo, A.; Muñor-Bellido, J.L.; Belhassen-García, M.; Sánchez-Seco, M.P.; Muro, A. A Novel RT-LAMP for the Detection of Different Genotypes of Crimean–Congo Haemorrhagic Fever Virus in Patients from Spain. Int. J. Mol. Sci. 2023, 24, 6411. https://doi.org/10.3390/ijms24076411
Febrer-Sendra B, Fernández-Soto P, García-Bernalt Diego J, Crego-Vicente B, Negredo A, Muñor-Bellido JL, Belhassen-García M, Sánchez-Seco MP, Muro A. A Novel RT-LAMP for the Detection of Different Genotypes of Crimean–Congo Haemorrhagic Fever Virus in Patients from Spain. International Journal of Molecular Sciences. 2023; 24(7):6411. https://doi.org/10.3390/ijms24076411
Chicago/Turabian StyleFebrer-Sendra, Begoña, Pedro Fernández-Soto, Juan García-Bernalt Diego, Beatriz Crego-Vicente, Anabel Negredo, Juan Luis Muñor-Bellido, Moncef Belhassen-García, María Paz Sánchez-Seco, and Antonio Muro. 2023. "A Novel RT-LAMP for the Detection of Different Genotypes of Crimean–Congo Haemorrhagic Fever Virus in Patients from Spain" International Journal of Molecular Sciences 24, no. 7: 6411. https://doi.org/10.3390/ijms24076411
APA StyleFebrer-Sendra, B., Fernández-Soto, P., García-Bernalt Diego, J., Crego-Vicente, B., Negredo, A., Muñor-Bellido, J. L., Belhassen-García, M., Sánchez-Seco, M. P., & Muro, A. (2023). A Novel RT-LAMP for the Detection of Different Genotypes of Crimean–Congo Haemorrhagic Fever Virus in Patients from Spain. International Journal of Molecular Sciences, 24(7), 6411. https://doi.org/10.3390/ijms24076411