

Anti-Mitochondrial and Insecticidal Effects of Artemisinin against Drosophila melanogaster

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
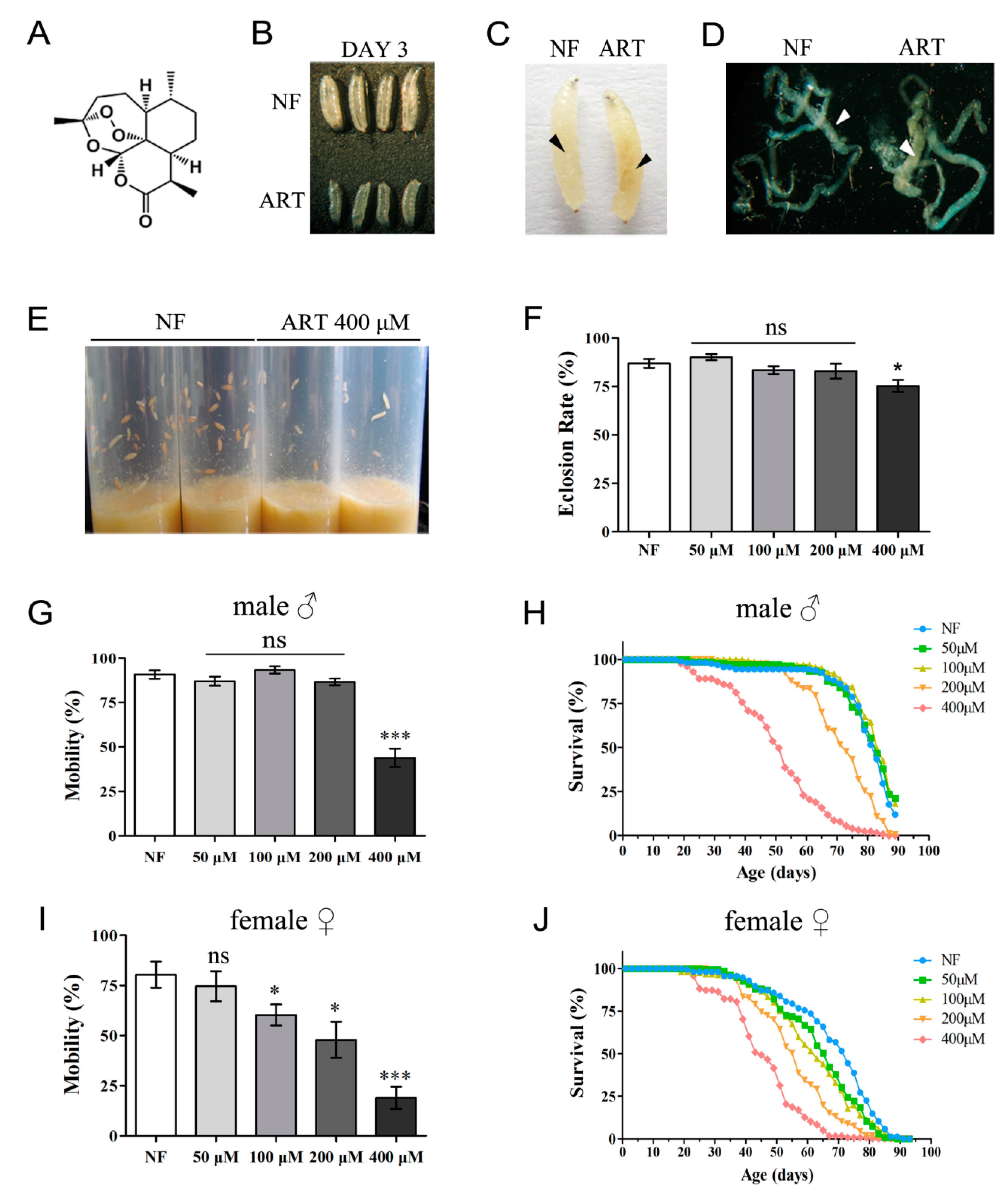
2.1. ART Negatively Affects D. melanogaster in a Concentration-Dependent Manner
2.2. Elevated Temperatures Exacerbate the Action of ART
2.3. ART Possesses Distinctly Separable Antifeeding and Insecticidal Activities
2.4. Drosophila Mitochondria Are Sensitive to the Direct Action of ART
3. Discussion
4. Materials and Methods
4.1. Fly Stocks, Culture Media Preparation
4.2. Eclosion Assay
4.3. Longevity Assay
4.4. Climbing Assay
4.5. Taste and Feeding Tendency Assay
4.6. Mitochondria Isolation
4.7. Determination of Mitochondrial Membrane Potential
4.8. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Tu, Y. Artemisinin-A Gift from Traditional Chinese Medicine to the World (Nobel Lecture). Angew. Chem. Int. Ed. Engl. 2016, 55, 10210–10226. [Google Scholar] [CrossRef] [PubMed]
- Tu, Y. The discovery of artemisinin (qinghaosu) and gifts from Chinese medicine. Nat. Med. 2011, 17, 1217–1220. [Google Scholar] [CrossRef] [PubMed]
- Zhou, B. Artemisinin (Qinghaosu): A mesmerizing drug that still puzzles. Sci. China Life Sci. 2015, 58, 1151–1153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, M.; Cao, X.; Jiang, Y.; Shi, Y.; Ma, Y.; Hu, D.; Song, X. Evaluation of the Combined Effect of Artemisinin and Ferroptosis Inducer RSL3 against Toxoplasma gondii. Int. J. Mol. Sci. 2022, 24, 229. [Google Scholar] [CrossRef] [PubMed]
- Machin, L.; Napoles, R.; Gille, L.; Monzote, L. Leishmania amazonensis response to artemisinin and derivatives. Parasitol. Int. 2021, 80, 102218. [Google Scholar] [CrossRef]
- Keiser, J.; Xiao, S.H.; Smith, T.A.; Utzinger, J. Combination chemotherapy against Clonorchis sinensis: Experiments with artemether, artesunate, OZ78, praziquantel, and tribendimidine in a rat model. Antimicrob. Agents. Chemother. 2009, 53, 3770–3776. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.X.; Wu, W.; Liang, Y.J.; Jie, Z.L.; Wang, H.; Wang, W.; Huang, Y.X. New uses for old drugs: The tale of artemisinin derivatives in the elimination of Schistosomiasis japonica in China. Molecules 2014, 19, 15058–15074. [Google Scholar] [CrossRef] [Green Version]
- Chaturvedi, D.; Goswami, A.; Saikia, P.P.; Barua, N.C.; Rao, P.G. Artemisinin and its derivatives: A novel class of anti-malarial and anti-cancer agents. Chem. Soc. Rev. 2010, 39, 435–454. [Google Scholar] [CrossRef]
- Ho, W.E.; Peh, H.Y.; Chan, T.K.; Wong, W.S. Artemisinins: Pharmacological actions beyond anti-malarial. Pharm. Ther. 2014, 142, 126–139. [Google Scholar] [CrossRef]
- Efferth, T.; Romero, M.R.; Wolf, D.G.; Stamminger, T.; Marin, J.J.; Marschall, M. The antiviral activities of artemisinin and artesunate. Clin. Infect. Dis. 2008, 47, 804–811. [Google Scholar] [CrossRef] [Green Version]
- Efferth, T. Beyond malaria: The inhibition of viruses by artemisinin-type compounds. Biotechnol. Adv. 2018, 36, 1730–1737. [Google Scholar] [CrossRef] [PubMed]
- Durden, K.; Sellars, S.; Cowell, B.; Brown, J.J.; Pszczolkowski, M.A. Artemisia annua extracts, artemisinin and 1,8-cineole, prevent fruit infestation by a major, cosmopolitan pest of apples. Pharm. Biol. 2011, 49, 563–568. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brisibe, E.A.; Adugbo, S.E.; Ekanem, U.; Brisibe, F.; Figueira, G.M. Controlling bruchid pests of stored cowpea seeds with dried leaves of Artemisia annua and two other common botanicals. Afr. J. Biotechnol. 2011, 10, 9593–9599. [Google Scholar] [CrossRef] [Green Version]
- Zibaee, A.; Bandani, A. A study on the toxicity of a medicinal plant, Artemisia annua L.(Asteracea) extracts to the sunn pest, Eurygaster integriceps Puton (Hemiptera: Scutelleridae). J. Plant Prot. Res. 2010, 50, 79–85. [Google Scholar] [CrossRef]
- Maggi, M.E.; Mangeaud, A.; Carpinella, M.C.; Ferrayoli, C.G.; Valladares, G.R.; Palacios, S.M. Laboratory evaluation of Artemisia annua L. extract and artemisinin activity against Epilachna paenulata and Spodoptera eridania. J. Chem. Ecol. 2005, 31, 1527–1536. [Google Scholar] [CrossRef] [PubMed]
- Hasheminia, S.M.; Sendi, J.J.; Jahromi, K.T.; Moharramipour, S. The effects of Artemisia annua L. and Achillea millefolium L. crude leaf extracts on the toxicity, development, feeding efficiency and chemical activities of small cabbage Pieris rapae L. (Lepidoptera: Pieridae). Pestic. Biochem Physiol. 2011, 99, 244–249. [Google Scholar] [CrossRef]
- Tripathi, A.K.; Prajapati, V.; Aggarwal, K.K.; Khanuja, S.P.; Kumar, S. Repellency and toxicity of oil from Artemisia annua to certain stored-product beetles. J. Econ. Entomol. 2000, 93, 43–47. [Google Scholar] [CrossRef]
- Shekari, M.; Sendi, J.J.; Etebari, K.; Zibaee, A.; Shadparvar, A. Effects of Artemisia annua L. (Asteracea) on nutritional physiology and enzyme activities of elm leaf beetle, Xanthogaleruca luteola Mull. (Coleoptera: Chrysomellidae). Pestic. Biochem. Physiol. 2008, 91, 66–74. [Google Scholar] [CrossRef]
- Deb, M.; Kumar, D. Bioactivity and efficacy of essential oils extracted from Artemisia annua against Tribolium casteneum (Herbst. 1797) (Coleoptera: Tenebrionidae): An eco-friendly approach. Ecotoxicol. Environ. Saf. 2020, 189, 109988. [Google Scholar] [CrossRef]
- Knudsmark Jessing, K.; Duke, S.O.; Cedergreeen, N. Potential ecological roles of artemisinin produced by Artemisia annua L. J. Chem. Ecol. 2014, 40, 100–117. [Google Scholar] [CrossRef]
- Zamani, S.; Sendi, J.J.; Ghadamyari, M. Effect of Artemisia annua L.(Asterales: Asteraceae) Essential Oil on Mortality, Development, Reproduction and Energy Reserves of Plodia Interpunctella (Hubner). (Lepidoptera: Pyralidae). J. Biofertil. Biopestici. 2011, 2, 105. [Google Scholar] [CrossRef] [Green Version]
- Allen, P.C.; Lydon, J.; Danforth, H.D. Effects of components of Artemisia annua on coccidia infections in chickens. Poult. Sci. 1997, 76, 1156–1163. [Google Scholar] [CrossRef]
- Paramanik, R.; Chikkaswamy, B.; Roy, D.; Paramanik, A.; Kumar, V. Effects of biochemicals of Artemisia annua in plants. J. Phytol. Res. 2008, 21, 11–18. [Google Scholar]
- Chattopadhyay, D.; Sen, S.; Chatterjee, R.; Roy, D.; James, J.; Thirumurugan, K. Context- and dose-dependent modulatory effects of naringenin on survival and development of Drosophila melanogaster. Biogerontology 2016, 17, 383–393. [Google Scholar] [CrossRef] [PubMed]
- Pohleven, J.; Brzin, J.; Vrabec, L.; Leonardi, A.; Cokl, A.; Strukelj, B.; Kos, J.; Sabotic, J. Basidiomycete Clitocybe nebularis is rich in lectins with insecticidal activities. Appl. Microbiol. Biotechnol. 2011, 91, 1141–1148. [Google Scholar] [CrossRef]
- Lee, S.H.; An, H.S.; Jung, Y.W.; Lee, E.J.; Lee, H.Y.; Choi, E.S.; An, S.W.; Son, H.; Lee, S.J.; Kim, J.B.; et al. Korean mistletoe (Viscum album coloratum) extract extends the lifespan of nematodes and fruit flies. Biogerontology 2014, 15, 153–164. [Google Scholar] [CrossRef]
- Sun, C.; Li, J.; Cao, Y.; Long, G.; Zhou, B. Two distinct and competitive pathways confer the cellcidal actions of artemisinins. Microb. Cell 2015, 2, 14–25. [Google Scholar] [CrossRef] [Green Version]
- Antoine, T.; Fisher, N.; Amewu, R.; O’Neill, P.M.; Ward, S.A.; Biagini, G.A. Rapid kill of malaria parasites by artemisinin and semi-synthetic endoperoxides involves ROS-dependent depolarization of the membrane potential. J. Antimicrob. Chemother. 2014, 69, 1005–1016. [Google Scholar] [CrossRef]
- Wang, J.; Huang, L.; Li, J.; Fan, Q.; Long, Y.; Li, Y.; Zhou, B. Artemisinin directly targets malarial mitochondria through its specific mitochondrial activation. PLoS ONE 2010, 5, e9582. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Zhou, B. The molecular and cellular action properties of artemisinins: What has yeast told us? Microb. Cell 2016, 3, 196–205. [Google Scholar] [CrossRef] [Green Version]
- Duke, S.O.; Vaughn, K.C.; Croom, E.M., Jr.; Elsohly, H.N. Artemisinin, a constituent of annual wormwood (Artemisia annua), is a selective phytotoxin. Weed Sci. 1987, 35, 499–505. [Google Scholar] [CrossRef]
- Emadi, F.; Yassa, N. Chemical composition of Iranian Artemisia annua L. essential oil and its antibacterial, antifungal and antioxidant effects. Planta. Med. 2009, 75, PJ91. [Google Scholar] [CrossRef]
- Soylu, E.; Yigitbas, H.; Tok, F.; Soylu, S.; Kurt, S.; Baysal, O.; Kaya, A. Chemical composition and antifungal activity of the essential oil of Artemisia annua L. against foliar and soil-borne fungal pathogens. Z. Pflanzenkrankh. Pflanzenschutz. J. Plant Dis. Prot. 2005, 112, 229–239. [Google Scholar]
- Palacios, S.M.; Bertoni, A.; Rossi, Y.; Santander, R.; Urzua, A. Insecticidal activity of essential oils from native medicinal plants of Central Argentina against the house fly, Musca domestica (L.). Parasitol. Res. 2009, 106, 207–212. [Google Scholar] [CrossRef]
- Lydon, J.; Teasdale, J.R.; Chen, P.K. Allelopathic activity of annual wormwood (Artemisia annua) and the role of artemisinin. Weed Sci. 1997, 45, 807–811. [Google Scholar] [CrossRef]
- Delabays, N.; Slacanin, I.; Bohren, C. Herbicidal potential of artemisinin and allelopathic properties of Artemisia annua L.: From the laboratory to the field. J. Plant Dis. Prot. 2008, 21, 317–322. [Google Scholar]
- Chen, P.K.; Leather, G.R. Plant growth regulatory activities of artemisinin and its related compounds. J. Chem. Ecol. 1990, 16, 1867–1876. [Google Scholar] [CrossRef]
- Ni, L.; Acharya, K.; Hao, X.; Li, S. Isolation and identification of an anti-algal compound from Artemisia annua and mechanisms of inhibitory effect on algae. Chemosphere 2012, 88, 1051–1057. [Google Scholar] [CrossRef]
- Laughlin, J.C. The influence of distribution of antimalarial constituents in Artemisia annua L. on time and method of harvest. Int. Soc. Hortic. Sci. 1995, 390, 67–74. [Google Scholar] [CrossRef]
- Jorge, F.S.F.; Janick, J. Floral Morphology of Artemisia annua with Special Reference to Trichomes. Int. J. Plant Sci. 1995, 156, 807–815. [Google Scholar]
- Weathers, P.J.; Arsenault, P.R.; Covello, P.S.; McMickle, A.; Teoh, K.H.; Reed, D.W. Artemisinin production in Artemisia annua: Studies in planta and results of a novel delivery method for treating malaria and other neglected diseases. Phytochem. Rev. 2011, 10, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Brown, G.D. The biosynthesis of artemisinin (Qinghaosu) and the phytochemistry of Artemisia annua L. (Qinghao). Molecules 2010, 15, 7603–7698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira, J.F.; Janick, J. Distribution of artemisinin in Artemisia annua. In Progress in New Crops; Janick, J., Ed.; ASHS Press: Arlington, VA, USA, 1996; pp. 579–584. [Google Scholar]
- Delabays, N.; Collet, G.; Benakis, A. Selection and breeding for high artemisinin (qinghaosu) yielding strains of Artemisia annua. Int. Soc. Hortic. Sci. ISHS 1993, 303, 203–208. [Google Scholar] [CrossRef]
- Li, W.; Mo, W.; Shen, D.; Sun, L.; Wang, J.; Lu, S.; Gitschier, J.M.; Zhou, B. Yeast model uncovers dual roles of mitochondria in action of artemisinin. PLoS Genet 2005, 1, e36. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Zhou, B. Biological actions of artemisinin: Insights from medicinal chemistry studies. Molecules 2010, 15, 1378–1397. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zhang, X.; Zhou, B. Zinc Protoporphyrin-9 Potentiates the Anticancer Activity of Dihydroartemisinin. Antioxidants 2023, 12, 250. [Google Scholar] [CrossRef] [PubMed]
- El-Keredy, A.; Schleyer, M.; Konig, C.; Ekim, A.; Gerber, B. Behavioural analyses of quinine processing in choice, feeding and learning of larval Drosophila. PLoS ONE 2012, 7, e40525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miwa, S.; St-Pierre, J.; Partridge, L.; Brand, M.D. Superoxide and hydrogen peroxide production by Drosophila mi-tochondria. Free. Radic. Biol. Med. 2003, 35, 938–948. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhong, M.; Sun, C.; Zhou, B. Anti-Mitochondrial and Insecticidal Effects of Artemisinin against Drosophila melanogaster. Int. J. Mol. Sci. 2023, 24, 6912. https://doi.org/10.3390/ijms24086912
Zhong M, Sun C, Zhou B. Anti-Mitochondrial and Insecticidal Effects of Artemisinin against Drosophila melanogaster. International Journal of Molecular Sciences. 2023; 24(8):6912. https://doi.org/10.3390/ijms24086912
Chicago/Turabian StyleZhong, Mengjiao, Chen Sun, and Bing Zhou. 2023. "Anti-Mitochondrial and Insecticidal Effects of Artemisinin against Drosophila melanogaster" International Journal of Molecular Sciences 24, no. 8: 6912. https://doi.org/10.3390/ijms24086912
APA StyleZhong, M., Sun, C., & Zhou, B. (2023). Anti-Mitochondrial and Insecticidal Effects of Artemisinin against Drosophila melanogaster. International Journal of Molecular Sciences, 24(8), 6912. https://doi.org/10.3390/ijms24086912