
RUNX2 and Cancer

Abstract
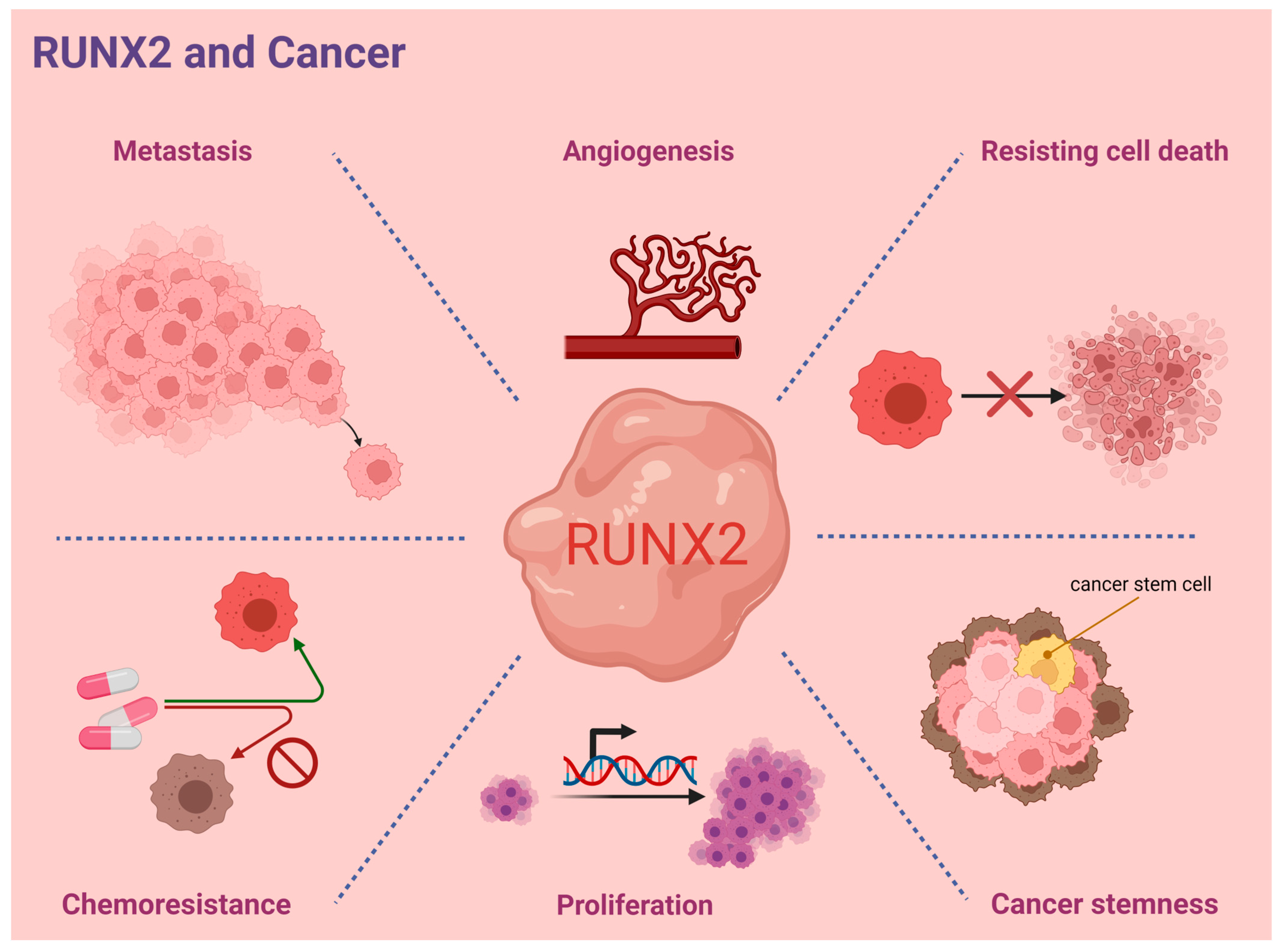
:1. Introduction
2. RUNX2 and Cancer Proliferation
3. RUNX2 and Angiogenesis
4. RUNX2 and Cancer Metastasis
5. RUNX2 and Drug Resistance
6. RUNX2, Transdifferentiation, and Cancer Stemness
7. RUNX2 Somatic Mutation and Cancer
8. RUNX2 Distribution and Expression in Normal Cell Types
9. RUNX2 Expression in Cancers
10. Correlation with Clinical Outcome
11. Summary
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kagoshima, H.; Shigesada, K.; Satake, M.; Ito, Y.; Miyoshi, H.; Ohki, M.; Pepling, M.; Gergen, P. The Runt domain identifies a new family of heteromeric transcriptional regulators. Trends Genet. 1993, 9, 338–341. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.C.; Yamaguchi-Iwai, Y.; Ogawa, E.; Maruyama, M.; Inuzuka, M.; Kagoshima, H.; Shigesada, K.; Satake, M.; Ito, Y. Isolation of PEBP2 alpha B cDNA representing the mouse homolog of human acute myeloid leukemia gene, AML1. Oncogene 1993, 8, 809–814. [Google Scholar] [PubMed]
- Komori, T.; Yagi, H.; Nomura, S.; Yamaguchi, A.; Sasaki, K.; Deguchi, K.; Shimizu, Y.; Bronson, R.T.; Gao, Y.H.; Inada, M.; et al. Targeted disruption of Cbfa1 results in a complete lack of bone formation owing to maturational arrest of osteoblasts. Cell 1997, 89, 755–764. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- van Wijnen, A.J.; Stein, G.S.; Gergen, J.P.; Groner, Y.; Hiebert, S.W.; Ito, Y.; Liu, P.; Neil, J.C.; Ohki, M.; Speck, N. Nomenclature for Runt-related (RUNX) proteins. Oncogene 2004, 23, 4209–4210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korinfskaya, S.; Parameswaran, S.; Weirauch, M.T.; Barski, A. Runx Transcription Factors in T Cells-What Is Beyond Thymic Development? Front. Immunol. 2021, 12, 701924. [Google Scholar] [CrossRef] [PubMed]
- Lian, J.B.; Javed, A.; Zaidi, S.K.; Lengner, C.; Montecino, M.; van Wijnen, A.J.; Stein, J.L.; Stein, G.S. Regulatory controls for osteoblast growth and differentiation: Role of Runx/Cbfa/AML factors. Crit. Rev. Eukaryot. Gene Expr. 2004, 14, 1–41. [Google Scholar] [CrossRef]
- Lian, J.B.; Stein, G.S. Runx2/Cbfa1: A multifunctional regulator of bone formation. Curr. Pharm. Des. 2003, 9, 2677–2685. [Google Scholar] [CrossRef]
- Komori, T. Requisite roles of Runx2 and Cbfb in skeletal development. J. Bone Miner. Metab. 2003, 21, 193–197. [Google Scholar] [CrossRef]
- Zhang, Y.W.; Yasui, N.; Ito, K.; Huang, G.; Fujii, M.; Hanai, J.; Nogami, H.; Ochi, T.; Miyazono, K.; Ito, Y. A RUNX2/PEBP2alpha A/CBFA1 mutation displaying impaired transactivation and Smad interaction in cleidocranial dysplasia. Proc. Natl. Acad. Sci. USA 2000, 97, 10549–10554. [Google Scholar] [CrossRef] [Green Version]
- Asghar, M.Y.; Knuutinen, T.; Holm, E.; Nordstrom, T.; Nguyen, V.D.; Zhou, Y.; Tornquist, K. Suppression of Calcium Entry Modulates the Expression of TRbeta1 and Runx2 in Thyroid Cancer Cells, Two Transcription Factors That Regulate Invasion, Proliferation and Thyroid-Specific Protein Levels. Cancers 2022, 14, 5838. [Google Scholar] [CrossRef]
- Song, X.; Liu, J.; Liu, B.; Piao, C.; Kong, C.; Li, Z. RUNX2 interacts with SCD1 and activates Wnt/beta-catenin signaling pathway to promote the progression of clear cell renal cell carcinoma. Cancer Med. 2023, 12, 5764–5780. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.Y.; Li, L.; Chen, S.L.; Yang, X.; Zhang, C.Z.; Cao, Y. A Zic2/Runx2/NOLC1 signaling axis mediates tumor growth and metastasis in clear cell renal cell carcinoma. Cell Death Dis. 2021, 12, 319. [Google Scholar] [CrossRef] [PubMed]
- Si, W.; Xu, X.; Wan, L.; Lv, F.; Wei, W.; Xu, X.; Li, W.; Huang, D.; Zhang, L.; Li, F. RUNX2 facilitates aggressiveness and chemoresistance of triple negative breast cancer cells via activating MMP1. Front. Oncol. 2022, 12, 996080. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Teng, X.; Ma, T.; Yang, T.; Zhang, J.; Huo, M.; Liu, W.; Yang, Y.; Yuan, B.; Yu, H.; et al. RUNX2 recruits the NuRD(MTA1)/CRL4B complex to promote breast cancer progression and bone metastasis. Cell Death Differ. 2022, 29, 2203–2217. [Google Scholar] [CrossRef]
- Ma, Y.; Gao, J.; Guo, H. miR-23a-3p Regulates Runx2 to Inhibit the Proliferation and Metastasis of Oral Squamous Cell Carcinoma. J. Oncol. 2022, 2022, 8719542. [Google Scholar] [CrossRef] [PubMed]
- Jin, G.; Ruan, Q.; Shangguan, F.; Lan, L. RUNX2 and LAMC2: Promising pancreatic cancer biomarkers identified by an integrative data mining of pancreatic adenocarcinoma tissues. Aging 2021, 13, 22963–22984. [Google Scholar] [CrossRef]
- Wang, C.; Shi, Z.; Zhang, Y.; Li, M.; Zhu, J.; Huang, Z.; Zhang, J.; Chen, J. CBFbeta promotes colorectal cancer progression through transcriptionally activating OPN, FAM129A, and UPP1 in a RUNX2-dependent manner. Cell Death Differ. 2021, 28, 3176–3192. [Google Scholar] [CrossRef]
- Liu, B.; Pan, S.; Liu, J.; Kong, C. Cancer-associated fibroblasts and the related Runt-related transcription factor 2 (RUNX2) promote bladder cancer progression. Gene 2021, 775, 145451. [Google Scholar] [CrossRef]
- Chen, Y.P.; Zhang, D.X.; Cao, Q.; He, C.K. LncRNA HCG18 Promotes Osteosarcoma Cells Proliferation, Migration, and Invasion in by Regulating miR-34a/RUNX2 Pathway. Biochem. Genet. 2022. [Google Scholar] [CrossRef]
- Guo, Q.; Ma, J.; Wu, J. MiRNA-218 inhibits cell proliferation, migration and invasion by targeting Runt-related transcription factor 2 (Runx2) in human osteosarcoma cells. Regen. Ther. 2021, 18, 508–515. [Google Scholar] [CrossRef]
- Zhou, M.; Zhang, P.; Zhao, Y.; Liu, R.; Zhang, Y. Overexpressed circRANBP17 acts as an oncogene to facilitate nasopharyngeal carcinoma via the miR-635/RUNX2 axis. J. Cancer 2021, 12, 4322–4331. [Google Scholar] [CrossRef] [PubMed]
- Bernal, C.; Otalora, A.; Canas, A.; Barreto, A.; Prieto, K.; Montecino, M.; Rojas, A. Regulatory Role of the RUNX2 Transcription Factor in Lung Cancer Apoptosis. Int. J. Cell Biol. 2022, 2022, 5198203. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Sun, B.; Zhao, X.; Zhang, Y.; Gu, Q.; Liang, X.; Dong, X.; Zhao, N. The Expression and Functional Significance of Runx2 in Hepatocellular Carcinoma: Its Role in Vasculogenic Mimicry and Epithelial-Mesenchymal Transition. Int. J. Mol. Sci. 2017, 18, 500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cecconi, D.; Brandi, J.; Manfredi, M.; Serena, M.; Dalle Carbonare, L.; Deiana, M.; Cheri, S.; Parolini, F.; Gandini, A.; Marchetto, G.; et al. Runx2 stimulates neoangiogenesis through the Runt domain in melanoma. Sci. Rep. 2019, 9, 8052. [Google Scholar] [CrossRef] [Green Version]
- Nesbitt, H.; Browne, G.; O’Donovan, K.M.; Byrne, N.M.; Worthington, J.; McKeown, S.R.; McKenna, D.J. Nitric Oxide Up-Regulates RUNX2 in LNCaP Prostate Tumours: Implications for Tumour Growth In Vitro and In Vivo. J. Cell. Physiol. 2016, 231, 473–482. [Google Scholar] [CrossRef] [PubMed]
- Colla, S.; Morandi, F.; Lazzaretti, M.; Rizzato, R.; Lunghi, P.; Bonomini, S.; Mancini, C.; Pedrazzoni, M.; Crugnola, M.; Rizzoli, V.; et al. Human myeloma cells express the bone regulating gene Runx2/Cbfa1 and produce osteopontin that is involved in angiogenesis in multiple myeloma patients. Leukemia 2005, 19, 2166–2176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bao, M.; Chen, Y.; Liu, J.T.; Bao, H.; Wang, W.B.; Qi, Y.X.; Lv, F. Extracellular matrix stiffness controls VEGF(165) secretion and neuroblastoma angiogenesis via the YAP/RUNX2/SRSF1 axis. Angiogenesis 2022, 25, 71–86. [Google Scholar] [CrossRef]
- Ma, J.; Lu, H.; Wang, S.; Chen, B.; Liu, Z.; Ke, X.; Liu, T.; Fu, J. The anthraquinone derivative Emodin inhibits angiogenesis and metastasis through downregulating Runx2 activity in breast cancer. Int. J. Oncol. 2015, 46, 1619–1628. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Cai, R.; Ma, J.; Liu, T.; Ke, X.; Lu, H.; Fu, J. The natural compound codonolactone impairs tumor induced angiogenesis by downregulating BMP signaling in endothelial cells. Phytomedicine 2015, 22, 1017–1026. [Google Scholar] [CrossRef]
- Mochin, M.T.; Underwood, K.F.; Cooper, B.; McLenithan, J.C.; Pierce, A.D.; Nalvarte, C.; Arbiser, J.; Karlsson, A.I.; Moise, A.R.; Moskovitz, J.; et al. Hyperglycemia and redox status regulate RUNX2 DNA-binding and an angiogenic phenotype in endothelial cells. Microvasc. Res. 2015, 97, 55–64. [Google Scholar] [CrossRef] [Green Version]
- Pierce, A.D.; Anglin, I.E.; Vitolo, M.I.; Mochin, M.T.; Underwood, K.F.; Goldblum, S.E.; Kommineni, S.; Passaniti, A. Glucose-activated RUNX2 phosphorylation promotes endothelial cell proliferation and an angiogenic phenotype. J. Cell. Biochem. 2012, 113, 282–292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Ren, Z.; Liu, B.; Wei, S. RUNX2 Mediates Renal Cell Carcinoma Invasion through Calpain2. Biol. Pharm. Bull. 2022, 45, 1653–1659. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Sun, R.; Zhao, X.; Sun, B. RUNX2 promotes malignant progression in gastric cancer by regulating COL1A1. Cancer Biomark 2021, 31, 227–238. [Google Scholar] [CrossRef]
- Yi, H.; Li, G.; Long, Y.; Liang, W.; Cui, H.; Zhang, B.; Tan, Y.; Li, Y.; Shen, L.; Deng, D.; et al. Integrative multi-omics analysis of a colon cancer cell line with heterogeneous Wnt activity revealed RUNX2 as an epigenetic regulator of EMT. Oncogene 2020, 39, 5152–5164. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Han, D.; Chen, Z.; Han, C.; Dong, W.; Han, L.; Zou, L.; Zhang, J.; Liu, Y.; Chai, J. RUNX2 interacts with BRG1 to target CD44 for promoting invasion and migration of colorectal cancer cells. Cancer Cell Int. 2020, 20, 505. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Y.; Zhang, C.; Chen, X.; Huang, H.; Li, W.; Zhang, J.; Liu, Y. Upregulation of OGT by Caveolin-1 promotes hepatocellular carcinoma cell migration and invasion. Cell Biol. Int. 2021, 45, 2251–2263. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Huang, L.; Liu, L.; Wang, L.; Lin, W.; Zhu, X.; Su, W.; Lv, C. Knockdown of microRNA-203 reduces cisplatin chemo-sensitivity to osteosarcoma cell lines MG63 and U2OS in vitro by targeting RUNX2. J. Chemother. 2021, 33, 328–341. [Google Scholar] [CrossRef]
- Xie, J.; Yu, F.; Li, D.; Zhu, X.; Zhang, X.; Lv, Z. MicroRNA-218 regulates cisplatin (DPP) chemosensitivity in non-small cell lung cancer by targeting RUNX2. Tumour Biol. 2016, 37, 1197–1204. [Google Scholar] [CrossRef]
- Ozaki, T.; Sugimoto, H.; Nakamura, M.; Hiraoka, K.; Yoda, H.; Sang, M.; Fujiwara, K.; Nagase, H. Runt-related transcription factor 2 attenuates the transcriptional activity as well as DNA damage-mediated induction of pro-apoptotic TAp73 to regulate chemosensitivity. FEBS J. 2015, 282, 114–128. [Google Scholar] [CrossRef] [Green Version]
- Foley, J.M.; Scholten, D.J., 2nd; Monks, N.R.; Cherba, D.; Monsma, D.J.; Davidson, P.; Dylewski, D.; Dykema, K.; Winn, M.E.; Steensma, M.R. Anoikis-resistant subpopulations of human osteosarcoma display significant chemoresistance and are sensitive to targeted epigenetic therapies predicted by expression profiling. J. Transl. Med. 2015, 13, 110. [Google Scholar] [CrossRef] [Green Version]
- Zhang, C.; Xu, X.; Trotter, T.N.; Gowda, P.S.; Lu, Y.; Suto, M.J.; Javed, A.; Murphy-Ullrich, J.E.; Li, J.; Yang, Y. Runx2 Deficiency in Osteoblasts Promotes Myeloma Resistance to Bortezomib by Increasing TSP-1-Dependent TGFbeta1 Activation and Suppressing Immunity in Bone Marrow. Mol. Cancer Ther. 2022, 21, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, L.; Xu, X.; He, X.; Wang, G.; Fan, C.; Zheng, Q.; Li, F. miR-205/RunX2 axis negatively regulates CD44(+)/CD24(-) breast cancer stem cell activity. Am. J. Cancer Res. 2020, 10, 1871–1887. [Google Scholar] [PubMed]
- Ma, F.; Xie, Y.; Lei, Y.; Kuang, Z.; Liu, X. The microRNA-130a-5p/RUNX2/STK32A network modulates tumor invasive and metastatic potential in non-small cell lung cancer. BMC Cancer 2020, 20, 580. [Google Scholar] [CrossRef] [PubMed]
- Pleasance, E.D. Pan-cancer analysis of whole genomes. Nature 2020, 578, 82–93. [Google Scholar] [CrossRef] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Gao, J.; Aksoy, B.A.; Dogrusoz, U.; Dresdner, G.; Gross, B.; Sumer, S.O.; Sun, Y.; Jacobsen, A.; Sinha, R.; Larsson, E.; et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Sci. Signal. 2013, 6, pl1. [Google Scholar] [CrossRef] [Green Version]
- Panagopoulos, I.; Gorunova, L.; Lobmaier, I.; Andersen, K.; Kostolomov, I.; Lund-Iversen, M.; Bjerkehagen, B.; Heim, S. FOS-ANKH and FOS-RUNX2 Fusion Genes in Osteoblastoma. Cancer Genom. Proteom. 2020, 17, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Gupta, S.; Ito, T.; Alex, D.; Vanderbilt, C.M.; Chang, J.C.; Islamdoust, N.; Zhang, Y.; Nafa, K.; Healey, J.; Ladanyi, M.; et al. RUNX2 (6p21.1) amplification in osteosarcoma. Hum. Pathol. 2019, 94, 23–28. [Google Scholar] [CrossRef]
- Nie, J.H.; Yang, T.; Li, H.; Ye, H.S.; Zhong, G.Q.; Li, T.T.; Zhang, C.; Huang, W.H.; Xiao, J.; Li, Z.; et al. Identification of GPC3 mutation and upregulation in a multidrug resistant osteosarcoma and its spheroids as therapeutic target. J. Bone Oncol. 2021, 30, 100391. [Google Scholar] [CrossRef] [PubMed]
- Lake, S.L.; Jmor, F.; Dopierala, J.; Taktak, A.F.; Coupland, S.E.; Damato, B.E. Multiplex ligation-dependent probe amplification of conjunctival melanoma reveals common BRAF V600E gene mutation and gene copy number changes. Investig. Ophthalmol. Vis. Sci. 2011, 52, 5598–5604. [Google Scholar] [CrossRef] [Green Version]
- Gao, R.; Kim, C.; Sei, E.; Foukakis, T.; Crosetto, N.; Chan, L.K.; Srinivasan, M.; Zhang, H.; Meric-Bernstam, F.; Navin, N. Nanogrid single-nucleus RNA sequencing reveals phenotypic diversity in breast cancer. Nat. Commun. 2017, 8, 228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, C.; Gao, R.; Sei, E.; Brandt, R.; Hartman, J.; Hatschek, T.; Crosetto, N.; Foukakis, T.; Navin, N.E. Chemoresistance Evolution in Triple-Negative Breast Cancer Delineated by Single-Cell Sequencing. Cell 2018, 173, 879–893 e813. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azizi, E.; Carr, A.J.; Plitas, G.; Cornish, A.E.; Konopacki, C.; Prabhakaran, S.; Nainys, J.; Wu, K.; Kiseliovas, V.; Setty, M.; et al. Single-Cell Map of Diverse Immune Phenotypes in the Breast Tumor Microenvironment. Cell 2018, 174, 1293–1308 e1236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chung, W.; Eum, H.H.; Lee, H.O.; Lee, K.M.; Lee, H.B.; Kim, K.T.; Ryu, H.S.; Kim, S.; Lee, J.E.; Park, Y.H.; et al. Single-cell RNA-seq enables comprehensive tumour and immune cell profiling in primary breast cancer. Nat. Commun. 2017, 8, 15081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, S.; Hebert, L.L.; Eschbacher, J.M.; Kim, S. Single-Cell RNA Sequencing of a Postmenopausal Normal Breast Tissue Identifies Multiple Cell Types That Contribute to Breast Cancer. Cancers 2020, 12, 3639. [Google Scholar] [CrossRef] [PubMed]
- Uhlen, M.; Bjorling, E.; Agaton, C.; Szigyarto, C.A.; Amini, B.; Andersen, E.; Andersson, A.C.; Angelidou, P.; Asplund, A.; Asplund, C.; et al. A human protein atlas for normal and cancer tissues based on antibody proteomics. Mol. Cell. Proteom. 2005, 4, 1920–1932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.Y.; Chen, Y.K.; Lo, S.; Chi, T.C.; Chen, Y.H.; Hu, S.C.; Chen, Y.W.; Jiang, S.S.; Tsai, F.Y.; Liu, W.; et al. MRE11 promotes oral cancer progression through RUNX2/CXCR4/AKT/FOXA2 signaling in a nuclease-independent manner. Oncogene 2021, 40, 3510–3532. [Google Scholar] [CrossRef] [PubMed]
- Cruz-De la Rosa, M.I.; Jimenez-Wences, H.; Alarcon-Millan, J.; Romero-Lopez, M.J.; Castanon-Sanchez, C.A.; Salmeron-Barcenas, E.G.; Fernandez-Tilapa, G. miR-218-5p/RUNX2 Axis Positively Regulates Proliferation and Is Associated with Poor Prognosis in Cervical Cancer. Int. J. Mol. Sci. 2022, 23, 6993. [Google Scholar] [CrossRef]
- Yang, D.P.; Huang, W.Y.; Chen, G.; Chen, S.W.; Yang, J.; He, R.Q.; Huang, S.N.; Gan, T.Q.; Ma, J.; Yang, L.J.; et al. Clinical significance of transcription factor RUNX2 in lung adenocarcinoma and its latent transcriptional regulating mechanism. Comput. Biol. Chem. 2020, 89, 107383. [Google Scholar] [CrossRef]
- Zhu, Y.; Chen, Q.Y.; Jordan, A.; Sun, H.; Roy, N.; Costa, M. RUNX2/miR-31/SATB2 pathway in nickel-induced BEAS-2B cell transformation. Oncol. Rep. 2021, 46, 154. [Google Scholar] [CrossRef]
- Zhu, J.; Sanborn, J.Z.; Benz, S.; Szeto, C.; Hsu, F.; Kuhn, R.M.; Karolchik, D.; Archie, J.; Lenburg, M.E.; Esserman, L.J.; et al. The UCSC Cancer Genomics Browser. Nat. Methods 2009, 6, 239–240. [Google Scholar] [CrossRef] [Green Version]
- Shinto, E.; Yoshida, Y.; Kajiwara, Y.; Okamoto, K.; Mochizuki, S.; Yamadera, M.; Shiraishi, T.; Nagata, K.; Tsuda, H.; Hase, K.; et al. Clinical Significance of a Gene Signature Generated from Tumor Budding Grade in Colon Cancer. Ann. Surg. Oncol. 2020, 27, 4044–4054. [Google Scholar] [CrossRef]
- Liang, S.; Li, Y.; Wang, B. The cancer-related transcription factor Runx2 combined with osteopontin: A novel prognostic biomarker in resected osteosarcoma. Int. J. Clin. Oncol. 2021, 26, 2347–2354. [Google Scholar] [CrossRef]
- Wang, H.X.; Wang, X.Y.; Fei, J.W.; Li, F.H.; Han, J.; Qin, X. microRNA-23B inhibits non-small cell lung cancer proliferation, invasion and migration via downregulation of RUNX2 and inhibition of Wnt/Beta-catenin signaling pathway. J. Biol. Regul. Homeost. Agents 2020, 34, 825–835. [Google Scholar] [CrossRef]
- Zhang, P.; Liu, L.; Zhang, L.; He, X.; Xu, X.; Lu, Y.; Li, F. Runx2 is required for activity of CD44(+)/CD24(-/low) breast cancer stem cell in breast cancer development. Am. J. Transl. Res. 2020, 12, 2305–2318. [Google Scholar]
- Uhlen, M.; Fagerberg, L.; Hallstrom, B.M.; Lindskog, C.; Oksvold, P.; Mardinoglu, A.; Sivertsson, A.; Kampf, C.; Sjostedt, E.; Asplund, A.; et al. Proteomics. Tissue-based map of the human proteome. Science 2015, 347, 1260419. [Google Scholar] [CrossRef]
- Thul, P.J.; Akesson, L.; Wiking, M.; Mahdessian, D.; Geladaki, A.; Ait Blal, H.; Alm, T.; Asplund, A.; Bjork, L.; Breckels, L.M.; et al. A subcellular map of the human proteome. Science 2017, 356, eaal3321. [Google Scholar] [CrossRef]
- Uhlen, M.; Oksvold, P.; Fagerberg, L.; Lundberg, E.; Jonasson, K.; Forsberg, M.; Zwahlen, M.; Kampf, C.; Wester, K.; Hober, S.; et al. Towards a knowledge-based Human Protein Atlas. Nat. Biotechnol. 2010, 28, 1248–1250. [Google Scholar] [CrossRef]
- Uhlen, M.; Zhang, C.; Lee, S.; Sjostedt, E.; Fagerberg, L.; Bidkhori, G.; Benfeitas, R.; Arif, M.; Liu, Z.; Edfors, F.; et al. A pathology atlas of the human cancer transcriptome. Science 2017, 357, eaan2507. [Google Scholar] [CrossRef] [Green Version]
- Gyorffy, B.; Schafer, R. Meta-analysis of gene expression profiles related to relapse-free survival in 1079 breast cancer patients. Breast Cancer Res. Treat. 2009, 118, 433–441. [Google Scholar] [CrossRef] [Green Version]
- Tahirov, T.H.; Inoue-Bungo, T.; Morii, H.; Fujikawa, A.; Sasaki, M.; Kimura, K.; Shiina, M.; Sato, K.; Kumasaka, T.; Yamamoto, M.; et al. Structural analyses of DNA recognition by the AML1/Runx-1 Runt domain and its allosteric control by CBFbeta. Cell 2001, 104, 755–767. [Google Scholar] [CrossRef] [PubMed]
- Xiao, G.; Jiang, D.; Gopalakrishnan, R.; Franceschi, R.T. Fibroblast growth factor 2 induction of the osteocalcin gene requires MAPK activity and phosphorylation of the osteoblast transcription factor, Cbfa1/Runx2. J. Biol. Chem. 2002, 277, 36181–36187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, G.; Jiang, D.; Thomas, P.; Benson, M.D.; Guan, K.; Karsenty, G.; Franceschi, R.T. MAPK pathways activate and phosphorylate the osteoblast-specific transcription factor, Cbfa1. J. Biol. Chem. 2000, 275, 4453–4459. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, E.J.; Lee, K.Y.; Choi, N.S.; Lee, M.H.; Kim, H.N.; Jin, Y.H.; Ryoo, H.M.; Choi, J.Y.; Yoshida, M.; Nishino, N.; et al. Bone morphogenetic protein-2 stimulates Runx2 acetylation. J. Biol. Chem. 2006, 281, 16502–16511. [Google Scholar] [CrossRef] [Green Version]
- Yoon, W.J.; Islam, R.; Cho, Y.D.; Woo, K.M.; Baek, J.H.; Uchida, T.; Komori, T.; van Wijnen, A.; Stein, J.L.; Lian, J.B.; et al. Pin1-mediated Runx2 modification is critical for skeletal development. J. Cell. Physiol. 2013, 228, 2377–2385. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample ID | Cancer Type | Protein Change | Mutation Type | Variant Type | Copy Number | Mutations in Sample |
|---|---|---|---|---|---|---|
| #1 | Uterine Endometrioid Carcinoma | S8N | Missense | SNP | Diploid | 568 |
| #2 | Esophagogastric Cancer | F17V | Missense | SNP | Gain | 141 |
| #3 | Prostate Cancer | S31Lfs*130 | FS ins | INS | Gain | 41 |
| #4 | Prostate Cancer | S31Lfs*130 | FS ins | INS | Gain | 48 |
| #5 | Prostate Cancer | S31Lfs*130 | FS ins | INS | Gain | 55 |
| #6 | Prostate Cancer | S31Lfs*130 | FS ins | INS | Gain | 54 |
| #7 | Prostate Cancer | S31Lfs*130 | FS ins | INS | Gain | 50 |
| #8 | Ovarian Cancer | L136V | Missense | SNP | Gain | 63 |
| #9 | Colorectal Cancer | D161N | Missense | SNP | Diploid | 4888 |
| #10 | Ovarian Cancer | K233E | Missense | SNP | Amp | 102 |
| #11 | Uterine Endometrioid Carcinoma | V261A | Missense | SNP | Diploid | 1321 |
| #12 | Hepatobiliary Cancer | S293P | Missense | SNP | Diploid | 68 |
| #13 | Melanoma | P320L | Missense | SNP | Amp | 1401 |
| #14 | Lung Cancer | R337M | Missense | SNP | Gain | 80 |
| #15 | Embryonal Tumor | S371Ffs*14 | FS ins | INS | Diploid | 17 |
| #16 | Glioma | P402L | Missense | SNP | Diploid | 77 |
| #8 | Ovarian Cancer | P402T | Missense | SNP | Gain | 63 |
| #17 | Breast Cancer | C434* | Nonsense | SNP | Diploid | 82 |
| #18 | Endometrial Cancer | E458K | Missense | SNP | Gain | 256 |
| #19 | Pancreatic Cancer | G459V | Missense | SNP | Diploid | 62 |
| #20 | Melanoma | P466L | Missense | SNP | Diploid | 950 |
| #13 | Melanoma | P466S | Missense | SNP | Amp | 1401 |
| #21 | Pancreatic Cancer | G512D | Missense | SNP | Diploid | 55 |
| Symbol | Cancer Type | Prognosis | Endpoint | p Value | Case | Dataset | Method | Probe ID |
|---|---|---|---|---|---|---|---|---|
| RUNX2 | Glioma | Poor | Overall survival | 0.02 | 153 | TCGA | RNA Seq | |
| RUNX2 | Thyroid Cancer | - | Overall survival | N.S. | 501 | TCGA | RNA Seq | |
| RUNX2 | Lung Cancer | - | Overall survival | N.S. | 994 | TCGA | RNA Seq | |
| RUNX2 | Colorectal Cancer | Poor | Overall survival | 0.04 | 597 | TCGA | RNA Seq | |
| RUNX2 | Head and Neck Cancer | - | Overall survival | N.S. | 499 | TCGA | RNA Seq | |
| RUNX2 | Stomach Cancer | Poor | Overall survival | <0.001 | 354 | TCGA | RNA Seq | |
| RUNX2 | Liver Cancer | - | Overall survival | N.S. | 365 | TCGA | RNA Seq | |
| RUNX2 | Pancreatic Cancer | Poor | Overall survival | 0.037 | 176 | TCGA | RNA Seq | |
| RUNX2 | Renal Cancer | Poor | Overall survival | <0.001 | 877 | TCGA | RNA Seq | |
| RUNX2 | Urothelial Cancer | Poor | Overall survival | <0.001 | 406 | TCGA | RNA Seq | |
| RUNX2 | Prostate Cancer | - | Overall survival | N.S. | 494 | TCGA | RNA Seq | |
| RUNX2 | Testis Cancer | - | Overall survival | N.S. | 134 | TCGA | RNA Seq | |
| RUNX2 | Breast Cancer | - | Overall survival | N.S. | 1075 | TCGA | RNA Seq | |
| RUNX2 | Cervical Cancer | Poor | Overall survival | 0.0089 | 291 | TCGA | RNA Seq | |
| RUNX2 | Endometrial Cancer | - | Overall survival | N.S. | 541 | TCGA | RNA Seq | |
| RUNX2 | Ovarian Cancer | - | Overall survival | N.S. | 373 | TCGA | RNA Seq | |
| RUNX2 | Melanoma | - | Overall survival | N.S. | 102 | TCGA | RNA Seq | |
| RUNX2 | Breast Cancer | Good | Relapse-free survival | <0.001 | 4929 | E-MTAB-365, E-TABM-43, GSE: 11,121, 12,093, | Array | 216994_s_at |
| 12,276, 1456, 16,391, 16,446, 16,716, 17,705, 17,907, | ||||||||
| 18,728, 19,615, 20,194, 20,271, 2034, 20,685, 20,711, | ||||||||
| 21,653, 22,093, 25,066, 2603, 26,971, 29,044, 2990, | ||||||||
| 31,448, 31,519, 32,646, 3494, 36,771, 37,946, 41,998, | ||||||||
| 42,568, 43,358, 43,365, 45,255, 4611, 46,184, 48,390, | ||||||||
| 50,948, 5327, 58,812, 61,304, 65,194, 6532, 69,031, | ||||||||
| 7390, 76,275, 78,958, 9195 | ||||||||
| RUNX2 | Ovarian Cancer | Good | Progression-free survival | 0.0037 | 1435 | GSE: 14,764, 15,622, 18,520, 19,829, 23,554, 26,193, | Array | 216994_s_at |
| 26,712, 27,651, 30,161, 3149, 51,373, 63,885, 65,986, | RNA Seq | |||||||
| 9891, TCGA (N = 565) | ||||||||
| RUNX2 | Lung Cancer | Poor | Postprogression survival | <0.001 | 1925 | CAARRAY, GSE: 14,814, 19,188, 29,013, 30,219, | Array | 216994_s_at |
| 31,210, 3141, 31,908, 37,745, 43,580, 4573, 50,081, | RNA Seq | |||||||
| 8894, TCGA (N = 133) | ||||||||
| RUNX2 | Gastric Cancer | Poor | Postprogression survival | <0.001 | 875 | GSE: 14,210, 15,459, 22,377, 29,272, 51,105, 62,254 | Array | 216994_s_at |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, T.-C. RUNX2 and Cancer. Int. J. Mol. Sci. 2023, 24, 7001. https://doi.org/10.3390/ijms24087001
Lin T-C. RUNX2 and Cancer. International Journal of Molecular Sciences. 2023; 24(8):7001. https://doi.org/10.3390/ijms24087001
Chicago/Turabian StyleLin, Tsung-Chieh. 2023. "RUNX2 and Cancer" International Journal of Molecular Sciences 24, no. 8: 7001. https://doi.org/10.3390/ijms24087001
APA StyleLin, T.-C. (2023). RUNX2 and Cancer. International Journal of Molecular Sciences, 24(8), 7001. https://doi.org/10.3390/ijms24087001