
The Control of the Crossover Localization in Allium
 , , , , and
, , , , and
Abstract
:1. Introduction
2. Results
2.1. GISH Analysis of Genomic Structure of F Hybrids (A. cepa × A. fistulosum)
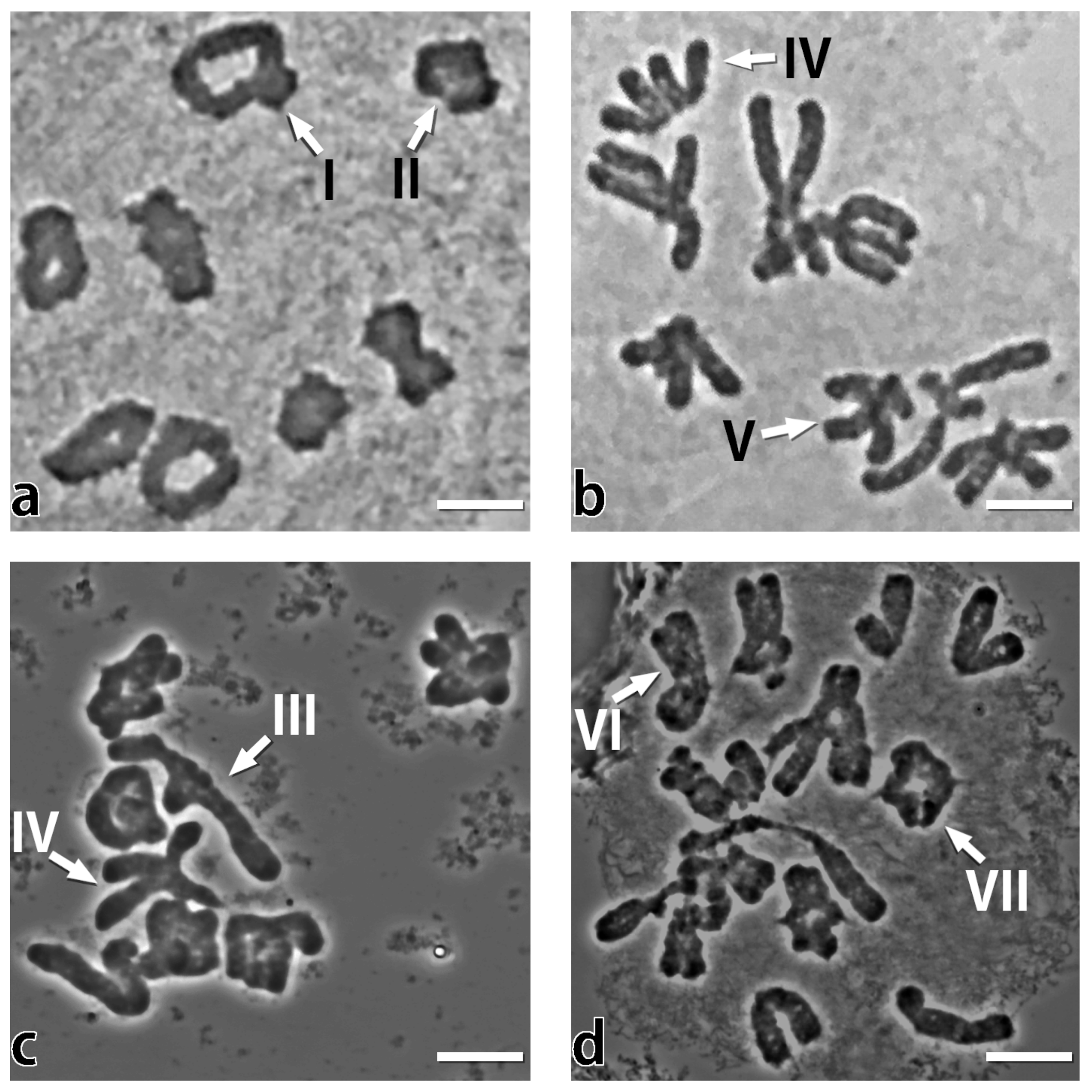
2.2. The Chiasmata Distribution in PMCs at Metaphase I
2.3. Behavior of ASY1 and ZYP1 during SC Assembly and Disassembly in A. cepa, A. fistulosum, and their F Diploid and Triploid Hybrids
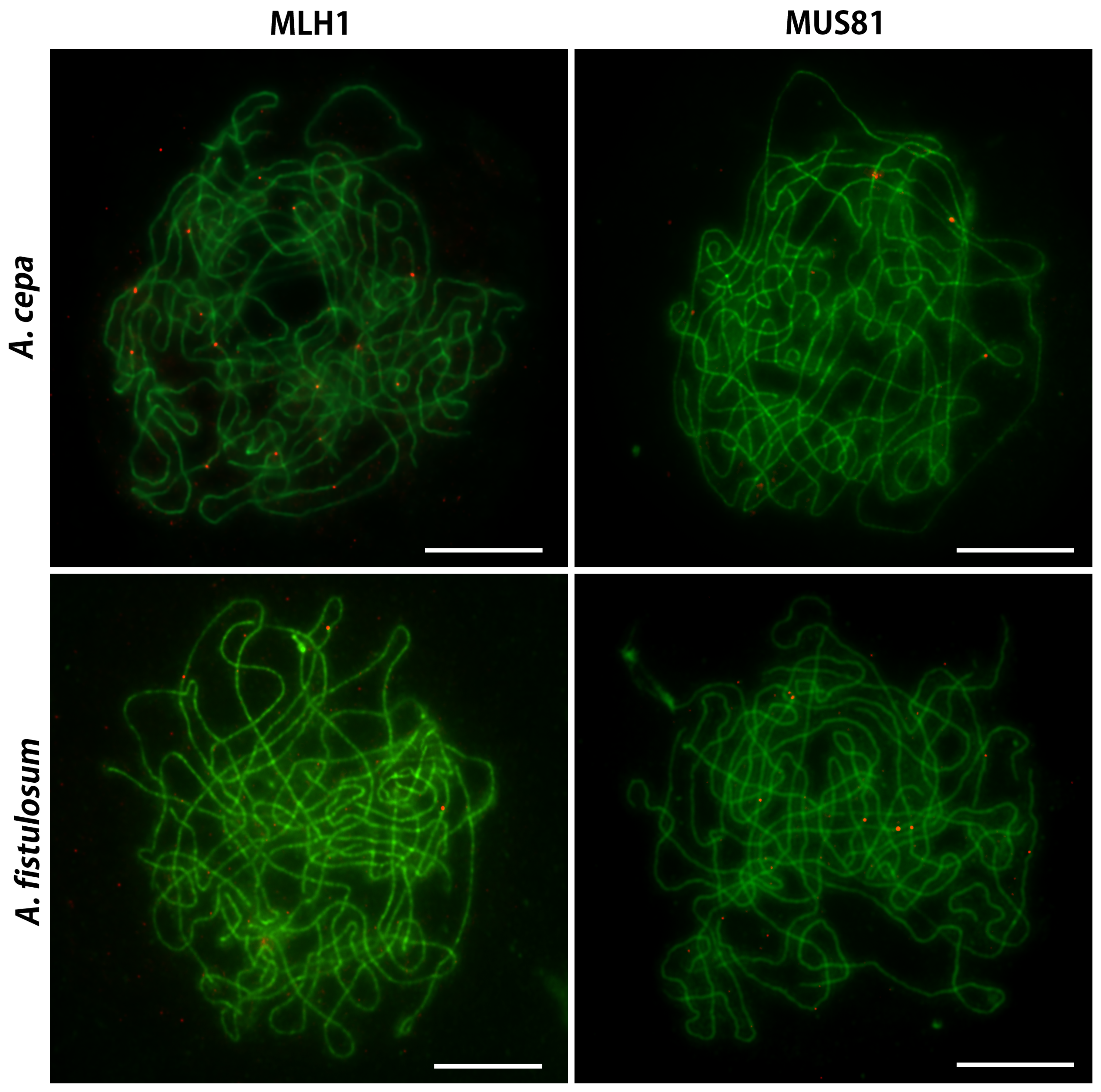
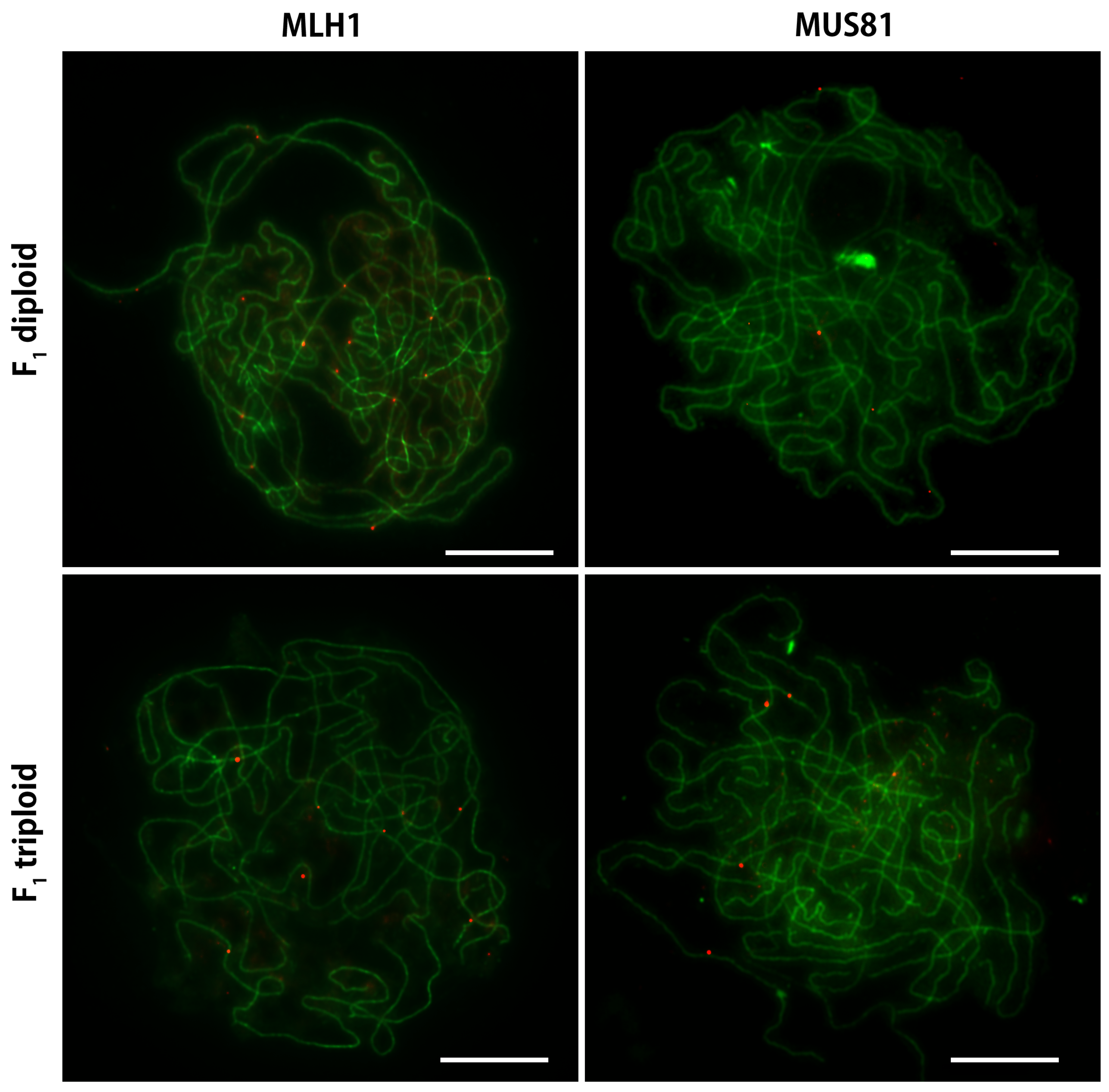
2.4. Immunocytological Detection of MLH1 and MUS81 on Pachytene Chromosomes of A. cepa and A. fistulosum and their F Diploid and Triploid Hybrids
3. Discussion
3.1. Chromatin, Genome and the Highly Contrasting Chiasmata Localization in A. cepa and A. fistulosum
3.2. The A. cepa Type of Chiasmata Distribution on Homeologous Bivalents in F Diploid Hybrid
3.3. A Significant Shift in Chiasmata Localization and MLH1/MUS81 Ratio in F Triploid Hybrid (2n = 3x = 16F + 8C) Is Evidence of Genetic Control of COs Distribution
4. Materials and Methods
4.1. Plant Materials
4.2. Genomic In Situ Hybridization (GISH)
4.3. Analysis of the Chiasmata Distribution in PMCs
4.4. Immunochemical Analysis
4.4.1. Antibody Production
4.4.2. Preparation of Meiotic Chromosome for Protein Immunolocalization
4.4.3. Immunochemical Detection of ASY1 and ZYP1
4.4.4. Immunochemical Detection of MLH1/ZYP1 and MUS81/ZYP1
4.4.5. Statistical Analysis
4.4.6. Microscopy and Imaging
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CO | Crossing Over |
| DSB | Double-Strand Breaks |
| PMC | Pollen Mother Cells |
| GISH | Genomic in situ hybridization |
| SC | Synaptonemal Complex |
| LN | Late recombination Nodules |
| ORF | Open Reading Frame |
| HJ | Holliday Junction |
References
- Darlington, C. Meiosis in diploid and tetraploid Primula sinensis. J. Genet. 1931, 24, 65–96. [Google Scholar] [CrossRef]
- Higgins, J.; Osman, K.; Jones, G.; Franklin, F. Factors underlying restricted crossover localization in barley meiosis. Annu. Rev. Genet. 2014, 48, 29–47. [Google Scholar] [CrossRef] [Green Version]
- Lam, I.; Keeney, S. Mechanism and regulation of meiotic recombination initiation. Cold Spring Harb. Perspect. Biol. 2015, 7, a016634. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercier, R.; Mézard, C.; Jenczewski, E.; Macaisne, N.; Grelon, M. The molecular biology of meiosis in plants. Annu. Rev. Plant Biol. 2015, 66, 297–327. [Google Scholar] [CrossRef]
- Gray, S.; Cohen, P.E. Control of meiotic crossovers: From double-strand break formation to designation. Annu. Rev. Genet. 2016, 50, 175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haenel, Q.; Laurentino, T.G.; Roesti, M.; Berner, D. Meta-analysis of chromosome-scale crossover rate variation in eukaryotes and its significance to evolutionary genomics. Mol. Ecol. 2018, 27, 2477–2497. [Google Scholar] [CrossRef] [Green Version]
- Gardiner, L.J.; Wingen, L.U.; Bailey, P.; Joynson, R.; Brabbs, T.; Wright, J.; Higgins, J.D.; Hall, N.; Griffiths, S.; Clavijo, B.J.; et al. Analysis of the recombination landscape of hexaploid bread wheat reveals genes controlling recombination and gene conversion frequency. Genome Biol. 2019, 20, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Desjardins, S.D.; Simmonds, J.; Guterman, I.; Kanyuka, K.; Burridge, A.J.; Tock, A.J.; Sanchez-Moran, E.; Franklin, F.C.H.; Henderson, I.R.; Edwards, K.J.; et al. FANCM promotes class I interfering crossovers and suppresses class II non-interfering crossovers in wheat meiosis. Nat. Commun. 2022, 13, 3644. [Google Scholar] [CrossRef]
- Emsweller, S.; Jones, H. A Gene for Control of Interstitial Localization of Chiasmata in Allium fistulosum L. Science 1935, 81, 543–544. [Google Scholar] [CrossRef]
- Emsweller, S.; Jones, H. Meiosis in Allium fistulosum, Allium cepa, and their hybrid. Hilgardia 1935, 9, 275–294. [Google Scholar] [CrossRef] [Green Version]
- Vosa, C.G. Heterochromatic patterns in Allium. Heredity 1976, 36, 383–392. [Google Scholar] [CrossRef] [Green Version]
- Tease, C. Cytological detection of crossing-over in BUdR substituted meiotic chromosomes using the fluorescent plus Giemsa technique. Nature 1978, 272, 823–824. [Google Scholar] [CrossRef]
- Tease, C.; Jones, G. Analysis of exchanges in differentially stained meiotic chromosomes of Locusta migratoria after BrdU-substitution and FPG staining. Chromosoma 1978, 69, 163–178. [Google Scholar] [CrossRef]
- Bishop, D.K.; Zickler, D. Early decision: Meiotic crossover interference prior to stable strand exchange and synapsis. Cell 2004, 117, 9–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillers, K.J. Crossover interference. Curr. Biol. 2004, 14, R1036–R1037. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kleckner, N.; Zickler, D.; Jones, G.H.; Dekker, J.; Padmore, R.; Henle, J.; Hutchinson, J. A mechanical basis for chromosome function. Proc. Natl. Acad. Sci. USA 2004, 101, 12592–12597. [Google Scholar] [CrossRef] [Green Version]
- Moens, P.B.; Kolas, N.K.; Tarsounas, M.; Marcon, E.; Cohen, P.E.; Spyropoulos, B. The time course and chromosomal localization of recombination-related proteins at meiosis in the mouse are compatible with models that can resolve the early DNA-DNA interactions without reciprocal recombination. J. Cell Sci. 2002, 115, 1611–1622. [Google Scholar] [CrossRef]
- Martini, E.; Diaz, R.L.; Hunter, N.; Keeney, S. Crossover homeostasis in yeast meiosis. Cell 2006, 126, 285–295. [Google Scholar] [CrossRef] [Green Version]
- Pazhayam, N.M.; Turcotte, C.A.; Sekelsky, J. Meiotic crossover patterning. Front. Cell Dev. Biol. 2021, 9, 681123. [Google Scholar] [CrossRef]
- Choi, K.; Zhao, X.; Kelly, K.A.; Venn, O.; Higgins, J.D.; Yelina, N.E.; Hardcastle, T.J.; Ziolkowski, P.A.; Copenhaver, G.P.; Franklin, F.C.H.; et al. Arabidopsis meiotic crossover hot spots overlap with H2A.Z nucleosomes at gene promoters. Nat. Genet. 2013, 45, 1327–1336. [Google Scholar] [CrossRef] [Green Version]
- Muller, H.J. The mechanism of crossing-over. Am. Nat. 1916, 50, 193–221. [Google Scholar] [CrossRef]
- Sturtevant, A.H. The behavior of the chromosomes as studied through linkage. Z. FÜR Indukt. Abstammungs Und Vererbungslehre 1915, 13, 234–287. [Google Scholar] [CrossRef]
- Armstrong, S.J.; Caryl, A.P.; Jones, G.H.; Franklin, F.C.H. Asy1, a protein required for meiotic chromosome synapsis, localizes to axis-associated chromatin in Arabidopsis and Brassica. J. Cell Sci. 2002, 115, 3645–3655. [Google Scholar] [CrossRef] [Green Version]
- Lambing, C.; Kuo, P.C.; Tock, A.J.; Topp, S.D.; Henderson, I.R. ASY1 acts as a dosage-dependent antagonist of telomere-led recombination and mediates crossover interference in Arabidopsis. Proc. Natl. Acad. Sci. USA 2020, 117, 13647–13658. [Google Scholar] [CrossRef] [PubMed]
- Capilla-Pérez, L.; Durand, S.; Hurel, A.; Lian, Q.; Chambon, A.; Taochy, C.; Solier, V.; Grelon, M.; Mercier, R. The synaptonemal complex imposes crossover interference and heterochiasmy in Arabidopsis. Proc. Natl. Acad. Sci. USA 2021, 118, e2023613118. [Google Scholar] [CrossRef] [PubMed]
- France, M.G.; Enderle, J.; Röhrig, S.; Puchta, H.; Franklin, F.C.H.; Higgins, J.D. ZYP1 is required for obligate cross-over formation and cross-over interference in Arabidopsis. Proc. Natl. Acad. Sci. USA 2021, 118, e2021671118. [Google Scholar] [CrossRef]
- Storlazzi, A.; Xu, L.; Schwacha, A.; Kleckner, N. Synaptonemal complex (SC) component Zip1 plays a role in meiotic recombination independent of SC polymerization along the chromosomes. Proc. Natl. Acad. Sci. USA 1996, 93, 9043–9048. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, A. Crossover patterning in plants. Plant Reprod. 2022, 36, 55–72. [Google Scholar] [CrossRef]
- Lynn, A.; Soucek, R.; Börner, G.V. ZMM proteins during meiosis: Crossover artists at work. Chromosome Res. 2007, 15, 591–605. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakharyevich, K.; Tang, S.; Ma, Y.; Hunter, N. Delineation of joint molecule resolution pathways in meiosis identifies a crossover-specific resolvase. Cell 2012, 149, 334–347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hollingsworth, N.M.; Brill, S.J. The Mus81 solution to resolution: Generating meiotic crossovers without Holliday junctions. Genes Dev. 2004, 18, 117–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Muyt, A.; Jessop, L.; Kolar, E.; Sourirajan, A.; Chen, J.; Dayani, Y.; Lichten, M. BLM helicase ortholog Sgs1 is a central regulator of meiotic recombination intermediate metabolism. Mol. Cell 2012, 46, 43–53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berchowitz, L.E.; Francis, K.E.; Bey, A.L.; Copenhaver, G.P. The role of AtMUS81 in interference-insensitive crossovers in A. thaliana. PLoS Genet. 2007, 3, e132. [Google Scholar] [CrossRef]
- Crismani, W.; Girard, C.; Froger, N.; Pradillo, M.; Santos, J.L.; Chelysheva, L.; Copenhaver, G.P.; Horlow, C.; Mercier, R. FANCM limits meiotic crossovers. Science 2012, 336, 1588–1590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, J.D.; Buckling, E.F.; Franklin, F.C.H.; Jones, G.H. Expression and functional analysis of AtMUS81 in Arabidopsis meiosis reveals a role in the second pathway of crossing-over. Plant J. 2008, 54, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Heyer, W.D.; Ehmsen, K.T.; Solinger, J.A. Holliday junctions in the eukaryotic nucleus: Resolution in sight? Trends Biochem. Sci. 2003, 28, 548–557. [Google Scholar] [CrossRef]
- Osman, F.; Dixon, J.; Doe, C.L.; Whitby, M.C. Generating crossovers by resolution of nicked Holliday junctions: A role for Mus81-Eme1 in meiosis. Mol. Cell 2003, 12, 761–774. [Google Scholar] [CrossRef]
- Kurzbauer, M.T.; Pradillo, M.; Kerzendorfer, C.; Sims, J.; Ladurner, R.; Oliver, C.; Janisiw, M.P.; Mosiolek, M.; Schweizer, D.; Copenhaver, G.P.; et al. Arabidopsis thaliana FANCD2 promotes meiotic crossover formation. Plant Cell 2018, 30, 415–428. [Google Scholar] [CrossRef] [Green Version]
- Chelysheva, L.; Grandont, L.; Vrielynck, N.; Le Guin, S.; Mercier, R.; Grelon, M. An easy protocol for studying chromatin and recombination protein dynamics during Arabidopsis thaliana meiosis: Immunodetection of cohesins, histones and MLH1. Cytogenet. Genome Res. 2010, 129, 143–153. [Google Scholar] [CrossRef]
- Martín, A.C.; Shaw, P.; Phillips, D.; Reader, S.; Moore, G. Licensing MLH1 sites for crossover during meiosis. Nat. Commun. 2014, 5, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Lhuissier, F.G.; Offenberg, H.H.; Wittich, P.E.; Vischer, N.O.; Heyting, C. The mismatch repair protein MLH1 marks a subset of strongly interfering crossovers in tomato. Plant Cell 2007, 19, 862–876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, L.K.; Lohmiller, L.D.; Tang, X.; Hammond, D.B.; Javernick, L.; Shearer, L.; Basu-Roy, S.; Martin, O.C.; Falque, M. Combined fluorescent and electron microscopic imaging unveils the specific properties of two classes of meiotic crossovers. Proc. Natl. Acad. Sci. USA 2014, 111, 13415–13420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Desjardins, S.D.; Ogle, D.E.; Ayoub, M.A.; Heckmann, S.; Henderson, I.R.; Edwards, K.J.; Higgins, J.D. MutS homologue 4 and MutS homologue 5 maintain the obligate crossover in wheat despite stepwise gene loss following polyploidization. Plant Physiol. 2020, 183, 1545–1558. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Zhang, J.; Huang, J.; Xu, J.; Chen, Z.; Copenhaver, G.P.; Wang, Y. Regulation of interference-sensitive crossover distribution ensures crossover assurance in Arabidopsis. Proc. Natl. Acad. Sci. USA 2021, 118, e2107543118. [Google Scholar] [CrossRef] [PubMed]
- Holloway, J.K.; Booth, J.; Edelmann, W.; McGowan, C.H.; Cohen, P.E. MUS81 generates a subset of MLH1-MLH3–independent crossovers in mammalian meiosis. PLoS Genet. 2008, 4, e1000186. [Google Scholar] [CrossRef] [Green Version]
- Albini, S.; Jones, G. Synaptonemal complex spreading in Allium cepa and Allium fistulosum. III. The F1 hybrid. Genome 1990, 33, 854–866. [Google Scholar] [CrossRef]
- Netzer, D. Greenhouse technique to evaluate pink root disease caused by Pyrenochaeta terrestris. Euphytica 1985, 34, 385–391. [Google Scholar] [CrossRef]
- Galván, G.A.; Wietsma, W.; Putrasemedja, S.; Permadi, A.; Kik, C. Screening for resistance to anthracnose (Colletotrichum gloeosporioides Penz.) in Allium cepa and its wild relatives. Euphytica 1997, 95, 173–178. [Google Scholar] [CrossRef]
- Rabinowitch, H.D. Breeding alliaceous crops for pest resistance. In Proceedings of the I International Symposium on Edible Alliaceae 433, Mendoza, Argentina, 14–18 March 1994; pp. 223–246. [Google Scholar]
- Finkers, R.; van Kaauwen, M.; Ament, K.; Burger-Meijer, K.; Egging, R.; Huits, H.; Kodde, L.; Kroon, L.; Shigyo, M.; Sato, S.; et al. Insights from the first genome assembly of Onion (Allium cepa). G3 2021, 11, jkab243. [Google Scholar] [CrossRef]
- Baranyu, M.; Greilhuber, J. Genome size in Allium: In quest of reproducible data. Ann. Bot. 1999, 83, 687–695. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.J.R.; Carlton, P.M.; Golubovskaya, I.N.; Cande, W.Z. Interlock formation and coiling of meiotic chromosome axes during synapsis. Genetics 2009, 183, 905–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, M.; Wang, K.; Tang, D.; Wei, C.; Li, M.; Shen, Y.; Chi, Z.; Gu, M.; Cheng, Z. The central element protein ZEP1 of the synaptonemal complex regulates the number of crossovers during meiosis in rice. Plant Cell 2010, 22, 417–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golubovskaya, I.N.; Wang, C.R.; Timofejeva, L.; Cande, W.Z. Maize meiotic mutants with improper or non-homologous synapsis due to problems in pairing or synaptonemal complex formation. J. Exp. Bot. 2011, 62, 1533–1544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khoo, K.H.; Able, A.J.; Able, J.A. The isolation and characterisation of the wheat molecular ZIPper I homologue, TaZYP1. Bmc Res. Notes 2012, 5, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Higgins, J.D.; Sanchez-Moran, E.; Armstrong, S.J.; Jones, G.H.; Franklin, F.C.H. The Arabidopsis synaptonemal complex protein ZYP1 is required for chromosome synapsis and normal fidelity of crossing over. Genes Dev. 2005, 19, 2488–2500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mikhailova, E.; Phillips, D.; Sosnikhina, S.; Lovtsyus, A.; Jones, R.; Jenkins, G. Molecular assembly of meiotic proteins Asy1 and Zyp1 and pairing promiscuity in rye (Secale cereale L.) and its synaptic mutant sy10. Genetics 2006, 174, 1247–1258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, D.; Nibau, C.; Wnetrzak, J.; Jenkins, G. High resolution analysis of meiotic chromosome structure and behaviour in barley (Hordeum vulgare L.). PLoS ONE 2012, 7, e39539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hesse, S.; Zelkowski, M.; Mikhailova, E.I.; Keijzer, C.J.; Houben, A.; Schubert, V. Ultrastructure and dynamics of synaptonemal complex components during meiotic pairing and synapsis of standard (A) and accessory (B) rye chromosomes. Front. Plant Sci. 2019, 10, 773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehmsen, K.T.; Heyer, W.D. Biochemistry of meiotic recombination: Formation, processing, and resolution of recombination intermediates. In Recombination and Meiosis; Springer: Berlin/Heidelberg, Germany, 2008; pp. 91–164. [Google Scholar]
- Cuacos, M.; Lambing, C.; Pachon-Penalba, M.; Osman, K.; Armstrong, S.J.; Henderson, I.R.; Sanchez-Moran, E.; Franklin, F.C.H.; Heckmann, S. Meiotic chromosome axis remodelling is critical for meiotic recombination in Brassica rapa. J. Exp. Bot. 2021, 72, 3012–3027. [Google Scholar] [CrossRef] [PubMed]
- Knoll, A.; Higgins, J.D.; Seeliger, K.; Reha, S.J.; Dangel, N.J.; Bauknecht, M.; Schröpfer, S.; Franklin, F.C.H.; Puchta, H. The Fanconi anemia ortholog FANCM ensures ordered homologous recombination in both somatic and meiotic cells in Arabidopsis. Plant Cell 2012, 24, 1448–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paquin, K.L.; Mamrak, N.E.; Garzon, J.L.; Cantres-Velez, J.A.; Azzinaro, P.A.; Vuono, E.A.; Lima, K.E.; Camberg, J.L.; Howlett, N.G. FANCD2 binding to H4K20me2 via a methyl-binding domain is essential for efficient DNA cross-link repair. Mol. Cell. Biol. 2019, 39, e00194-19. [Google Scholar] [CrossRef] [Green Version]
- Lukaszewski, A.J. Unexpected behavior of an inverted rye chromosome arm in wheat. Chromosoma 2008, 117, 569–578. [Google Scholar] [CrossRef]
- Giraut, L.; Falque, M.; Drouaud, J.; Pereira, L.; Martin, O.C.; Mézard, C. Genome-wide crossover distribution in Arabidopsis thaliana meiosis reveals sex-specific patterns along chromosomes. PLoS Genet. 2011, 7, e1002354. [Google Scholar] [CrossRef] [Green Version]
- Bauer, E.; Falque, M.; Walter, H.; Bauland, C.; Camisan, C.; Campo, L.; Meyer, N.; Ranc, N.; Rincent, R.; Schipprack, W.; et al. Intraspecific variation of recombination rate in maize. Genome Biol. 2013, 14, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Levan, A. The cytology of the species hybrid Allium cepa × fistulosum and its polyploid derivatives. Hereditas 1941, 27, 253–272. [Google Scholar] [CrossRef]
- Jones, G.; Khazanehdari, K.; Ford-Lloyd, B. Meiosis in the leek (Allium porrum L.) revisited. II. Metaphase I observations. Heredity 1996, 76, 186–191. [Google Scholar] [CrossRef] [Green Version]
- Inada, I.; Endo, M. C-banded karyotype analysis of Allium fistulosum and A. altaicum and their phylogenetic relationship. J. Jpn. Soc. Hortic. Sci. 1994, 63, 593–602. [Google Scholar] [CrossRef] [Green Version]
- Newton, W.; Darlington, C. Fritillaria meleagris: Chiasmaformation and distribution. J. Genet. 1930, 22, 1–14. [Google Scholar] [CrossRef]
- Smilde, W.; Van Heusden, A.; Kik, C. AFLPs in leek (Allium porrum) are not inherited in large linkage blocks. Euphytica 1999, 110, 127–132. [Google Scholar] [CrossRef]
- Fesenko, I.; Khrustaleva, L.; Karlov, G. Organization of the 378-bp satellite repeat in terminal heterochromatin of Allium fistulosum. Russ. J. Genet. 2002, 38, 745–753. [Google Scholar] [CrossRef]
- Khrustaleva, L.; Kirov, I.; Romanov, D.; Budylin, M.; Lapitskaya, I.; Kiseleva, A.; Fesenko, I.; Karlov, G. The chromosome organization of genes and some types of extragenic DNA in Allium. Acta Hortic. 2012, 969, 43–52. [Google Scholar] [CrossRef]
- Higgins, J.D.; Perry, R.M.; Barakate, A.; Ramsay, L.; Waugh, R.; Halpin, C.; Armstrong, S.J.; Franklin, F.C.H. Spatiotemporal asymmetry of the meiotic program underlies the predominantly distal distribution of meiotic crossovers in barley. Plant Cell 2012, 24, 4096–4109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirouze, M.; Lieberman-Lazarovich, M.; Aversano, R.; Bucher, E.; Nicolet, J.; Reinders, J.; Paszkowski, J. Loss of DNA methylation affects the recombination landscape in Arabidopsis. Proc. Natl. Acad. Sci. USA 2012, 109, 5880–5885. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Underwood, C.J.; Choi, K.; Lambing, C.; Zhao, X.; Serra, H.; Borges, F.; Simorowski, J.; Ernst, E.; Jacob, Y.; Henderson, I.R.; et al. Epigenetic activation of meiotic recombination near Arabidopsis thaliana centromeres via loss of H3K9me2 and non-CG DNA methylation. Genome Res. 2018, 28, 519–531. [Google Scholar] [CrossRef] [Green Version]
- Hinch, A.G.; Zhang, G.; Becker, P.W.; Moralli, D.; Hinch, R.; Davies, B.; Bowden, R.; Donnelly, P. Factors influencing meiotic recombination revealed by whole-genome sequencing of single sperm. Science 2019, 363, eaau8861. [Google Scholar] [CrossRef]
- Dreissig, S.; Fuchs, J.; Cápal, P.; Kettles, N.; Byrne, E.; Houben, A. Measuring meiotic crossovers via multi-locus genotyping of single pollen grains in barley. PLoS ONE 2015, 10, e0137677. [Google Scholar] [CrossRef] [Green Version]
- Dreissig, S.; Fuchs, J.; Himmelbach, A.; Mascher, M.; Houben, A. Sequencing of single pollen nuclei reveals meiotic recombination events at megabase resolution and circumvents segregation distortion caused by postmeiotic processes. Front. Plant Sci. 2017, 8, 1620. [Google Scholar] [CrossRef] [Green Version]
- Kirov, I.V.; Kiseleva, A.V.; Van Laere, K.; Van Roy, N.; Khrustaleva, L.I. Tandem repeats of Allium fistulosum associated with major chromosomal landmarks. Mol. Genet. Genom. 2017, 292, 453–464. [Google Scholar] [CrossRef]
- Jenkins, G.; Rees, H. Synaptonemal complex formation in a Festuca hybrid. In Proceedings of the Kew Chromosome Conference II, Kew, UK, 1–4 September 1982; Alen & Unwin: London, UK, 1983; pp. 233–242. [Google Scholar]
- Jenkins, G.; White, J. Elimination of synaptonemal complex irregularities in a Lolium hybrid. Heredity 1990, 64, 45–53. [Google Scholar] [CrossRef]
- Renkawitz, J.; Lademann, C.A.; Jentsch, S. Mechanisms and principles of homology search during recombination. Nat. Rev. Mol. Cell Biol. 2014, 15, 369–383. [Google Scholar] [CrossRef]
- Zhang, L.; He, J.; He, H.; Wu, J.; Li, M. Genome-wide unbalanced expression bias and expression level dominance toward Brassica oleracea in artificially synthesized intergeneric hybrids of Raphanobrassica. Hortic. Res. 2021, 8, 246. [Google Scholar] [CrossRef] [PubMed]
- Cole, F.; Kauppi, L.; Lange, J.; Roig, I.; Wang, R.; Keeney, S.; Jasin, M. Homeostatic control of recombination is implemented progressively in mouse meiosis. Nat. Cell Biol. 2012, 14, 424–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kudryavtseva, N.; Havey, M.J.; Black, L.; Hanson, P.; Sokolov, P.; Odintsov, S.; Divashuk, M.; Khrustaleva, L. Cytological evaluations of advanced generations of interspecific hybrids between Allium cepa and Allium fistulosum showing resistance to Stemphylium vesicarium. Genes 2019, 10, 195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khrustaleva, L.; Kik, C. Cytogenetical studies in the bridge cross Allium cepa × (A. fistulosum × A. roylei). Theor. Appl. Genet. 1998, 96, 8–14. [Google Scholar] [CrossRef]
- Kirov, I.; Khrustaleva, L.; Van Laere, K.; Soloviev, A.; Meeus, S.; Romanov, D.; Fesenko, I. DRAWID: User-friendly java software for chromosome measurements and idiogram drawing. Comp. Cytogenet. 2017, 11, 747. [Google Scholar] [CrossRef]
- De Vries, J. Onion chromosome nomenclature and homoeology relationships workshop report. Euphytica 1990, 49, 1–3. [Google Scholar] [CrossRef]
- De Vries, J.; Jongerius, M. Interstitial C-bands on the chromosomes of Allium-species from the section cepa. In Proceedings of the Allium symposium: EUCARPIA-Section Vegetables, Coventry, UK, 6–9 September 1988; Institute of Horticultural Research: Bengaluru, India, 1988. [Google Scholar]
- Mannuss, A.; Dukowic-Schulze, S.; Suer, S.; Hartung, F.; Pacher, M.; Puchta, H. RAD5A, RECQ4A, and MUS81 have specific functions in homologous recombination and define different pathways of DNA repair in Arabidopsis thaliana. Plant Cell 2010, 22, 3318–3330. [Google Scholar] [CrossRef] [Green Version]
- Team, R.C. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. 2013. Available online: http://www.R-project.org/ (accessed on 10 January 2022).
- Mangiafico, S. Rcompanion: Functions to Support Extension Education Program Evaluation, R package version 2.4.21. 2023.



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Cells Scored | Mean PMC Chiasma Frequency | Number of Bivalents Analyzed | Chiasmata Location | |||
|---|---|---|---|---|---|---|
| Proximal % (Total) | Interstitial % (Total) | Distal % (Total) | ||||
| A. cepa | 54 | 19.1 | 435 | 1.8 (18) | 20.0 (207) | 78.2 (808) |
| A. fistulosum | 43 | 15.4 | 340 | 97.1 (643) | 1.2 (8) | 1.7 (11) |
| F (A. cepa × A. fistulosum) | ||||||
| (2n = 2x = 8F + 8C) | 35 | 12.0 | 248 | 11.4 (48) | 43.0 (180) | 45.6 (191) |
| F (A. cepa × A. fistulosum) | ||||||
| (2n = 3x = 16F + 8C) | 29 | 14.6 | 231 | 39.5 (167) | 26.0 (110) | 34.5 (146) |
| Pairwise Comparison | Cramer’s V | p Value * |
|---|---|---|
| A. cepa vs. A. fistulosum | 0.95 | <0.001 |
| A. fistulosum vs. F diploid hybrid | 0.87 | <0.001 |
| A. fistulosum vs. F triploid hybrid | 0.72 | <0.001 |
| A. cepa vs. F triploid hybrid | 0.49 | <0.001 |
| A. cepa vs. F diploid hybrid | 0.34 | <0.001 |
| F diploid hybrid vs. F triploid hybrid | 0.24 | <0.001 |
| MLH1 | MUS81 | Foci per Cell * | ||||||
|---|---|---|---|---|---|---|---|---|
| Total Cells | Total Signals | Mean Signals per Cell | Total Cells | Total Signals | Mean Signals per Cell | |||
| Allium cepa | 37 | 409 | 11.1 (73%) | 39 | 158 | 4.1 (27%) | 15.2 (100%) | |
| Allium fistulosum | 40 | 257 | 6.5 (50%) | 41 | 272 | 6.5 (50%) | 13.0 (100%) | |
| F (A. cepa × A. fistulosum) | ||||||||
| (2n = 2x = 8F + 8C) | 38 | 320 | 8.5 (70%) | 37 | 136 | 3.6 (30%) | 12.1 (100%) | |
| F (A. cepa × A. fistulosum) | ||||||||
| (2n = 3x = 16F + 8C) | 36 | 311 | 8.5 (60%) | 38 | 214 | 5.7 (40%) | 14.2 (100%) | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kudryavtseva, N.; Ermolaev, A.; Pivovarov, A.; Simanovsky, S.; Odintsov, S.; Khrustaleva, L. The Control of the Crossover Localization in Allium. Int. J. Mol. Sci. 2023, 24, 7066. https://doi.org/10.3390/ijms24087066
Kudryavtseva N, Ermolaev A, Pivovarov A, Simanovsky S, Odintsov S, Khrustaleva L. The Control of the Crossover Localization in Allium. International Journal of Molecular Sciences. 2023; 24(8):7066. https://doi.org/10.3390/ijms24087066
Chicago/Turabian StyleKudryavtseva, Natalia, Aleksey Ermolaev, Anton Pivovarov, Sergey Simanovsky, Sergey Odintsov, and Ludmila Khrustaleva. 2023. "The Control of the Crossover Localization in Allium" International Journal of Molecular Sciences 24, no. 8: 7066. https://doi.org/10.3390/ijms24087066