
The Role of Intercellular Signaling in the Regulation of Bacterial Adaptive Proliferation
 , and
, and 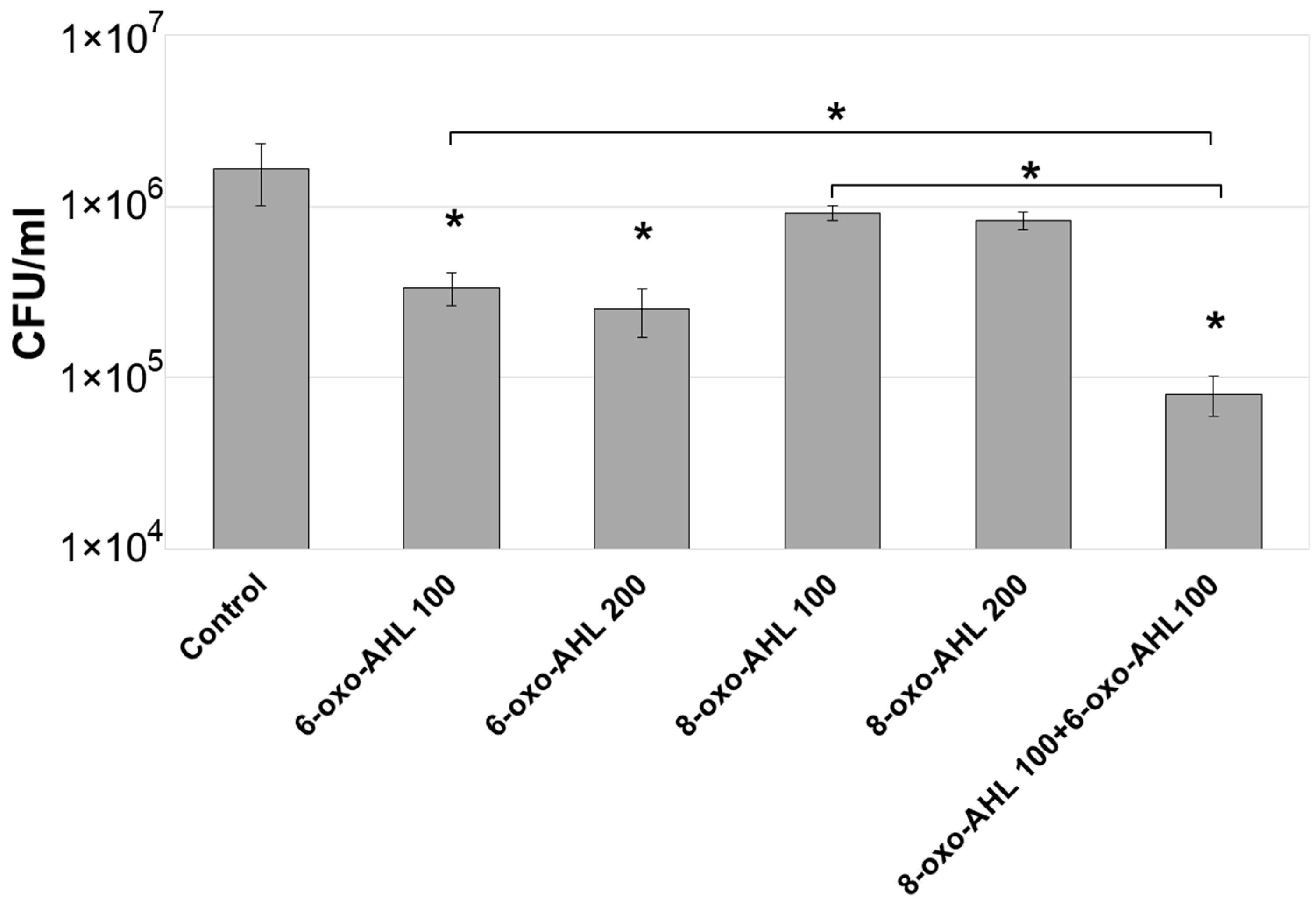{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of Exogenous AHL on the Adaptive Proliferation of Pba
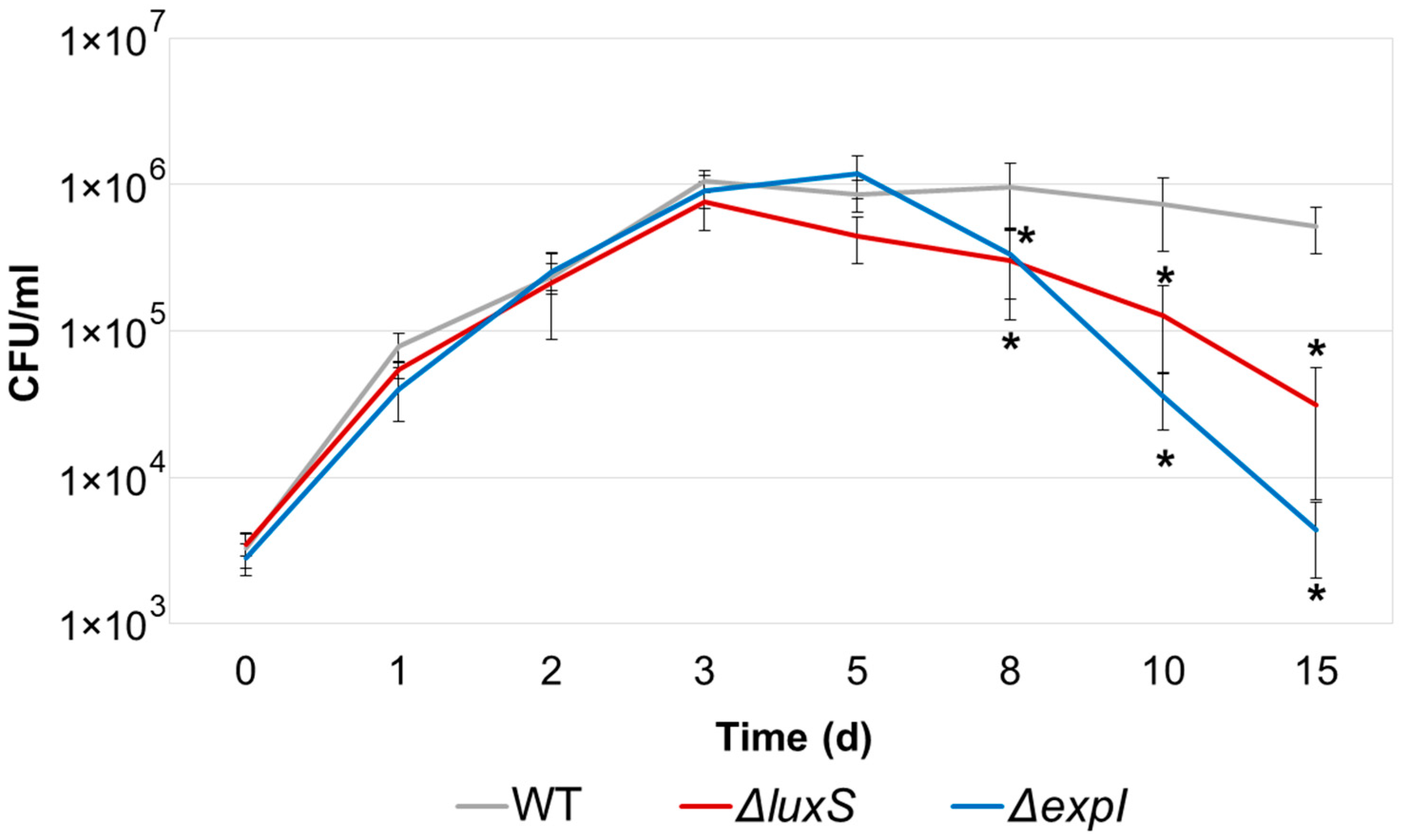
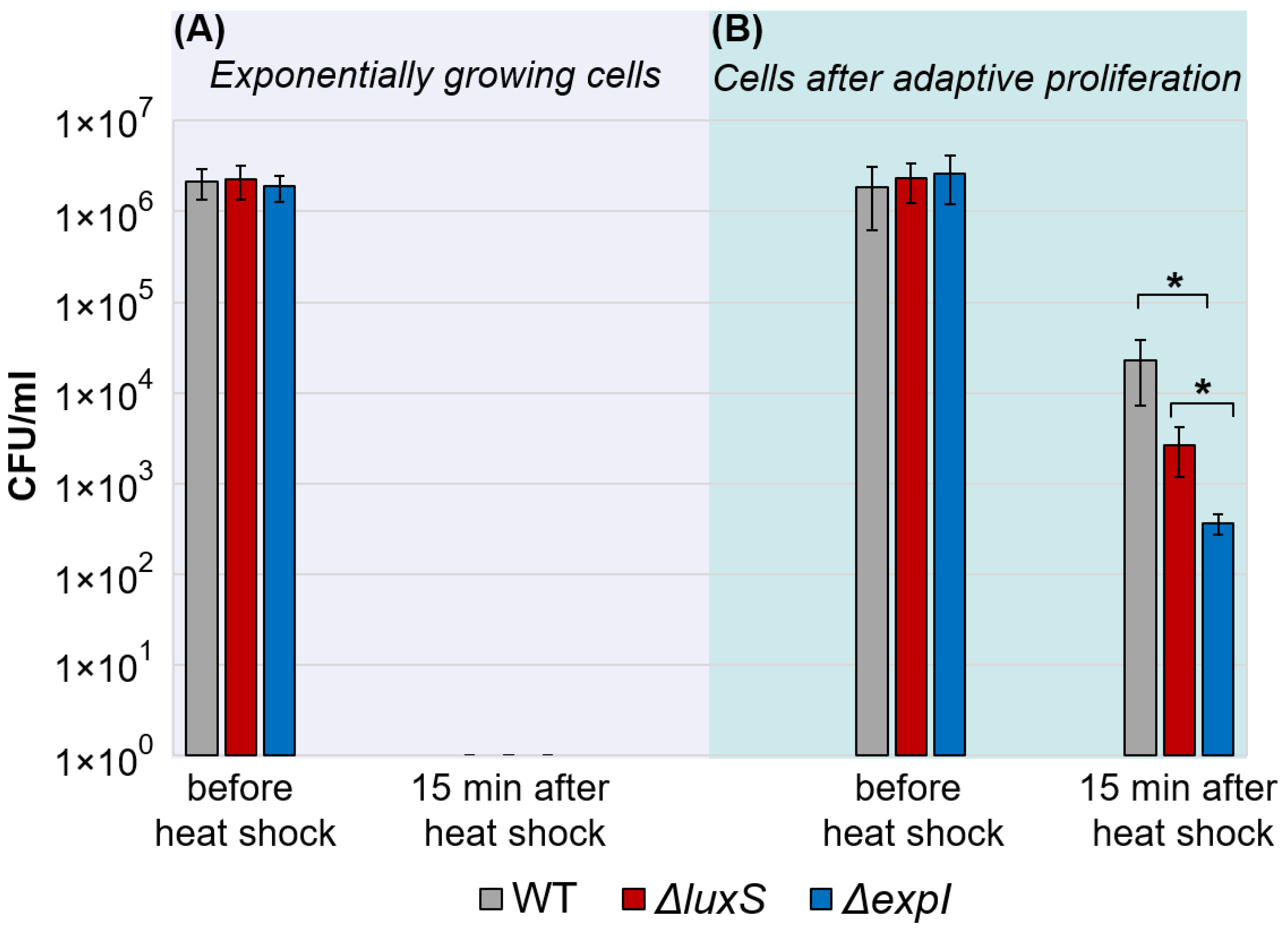
2.2. Adaptive Proliferation and Its Consequences in ΔexpI and ΔluxS Pba Mutants
2.3. Effect of the Depletion of AHL or AI-2 on the Potential of Culture Supernatants to Cause the Premature Termination of Adaptive Proliferation
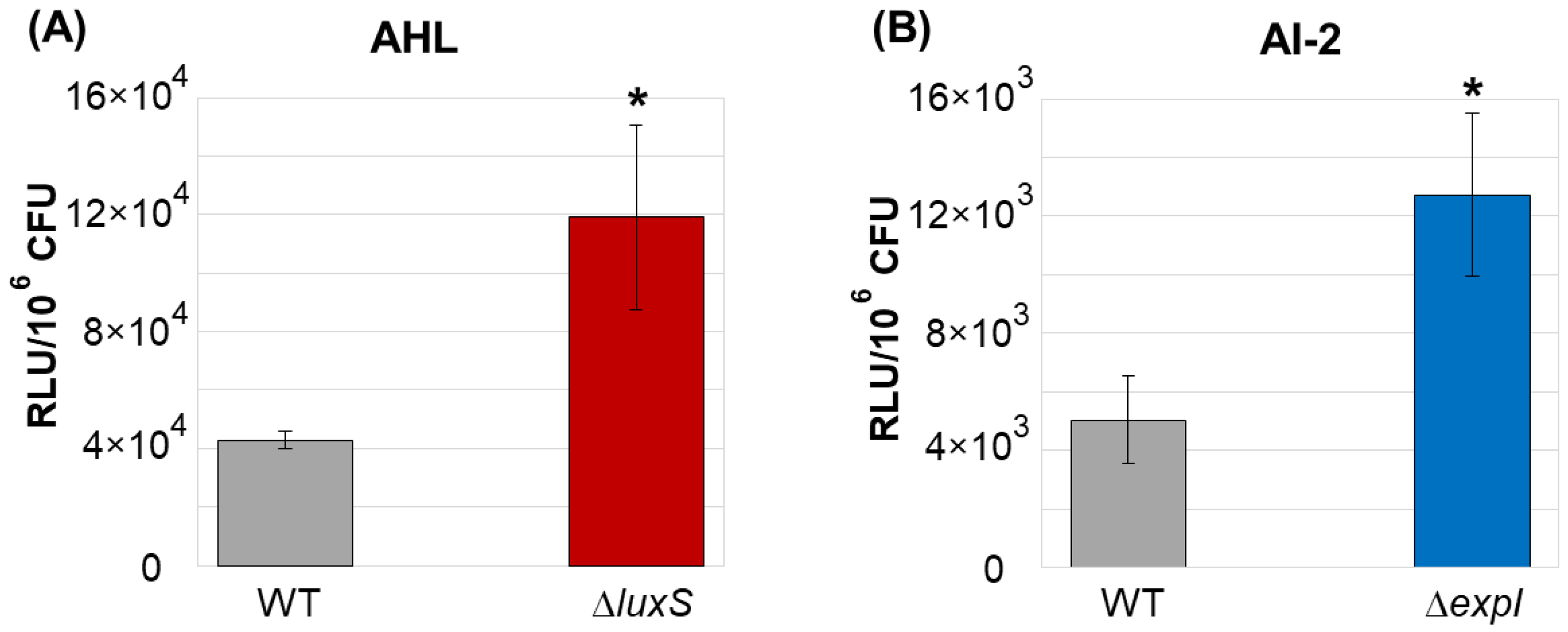
2.4. The Relative Levels of AHL and AI-2 in the Cultural Supernatants of the Wild Type Strain, ΔluxS and ΔexpI Mutants following Adaptive Proliferation
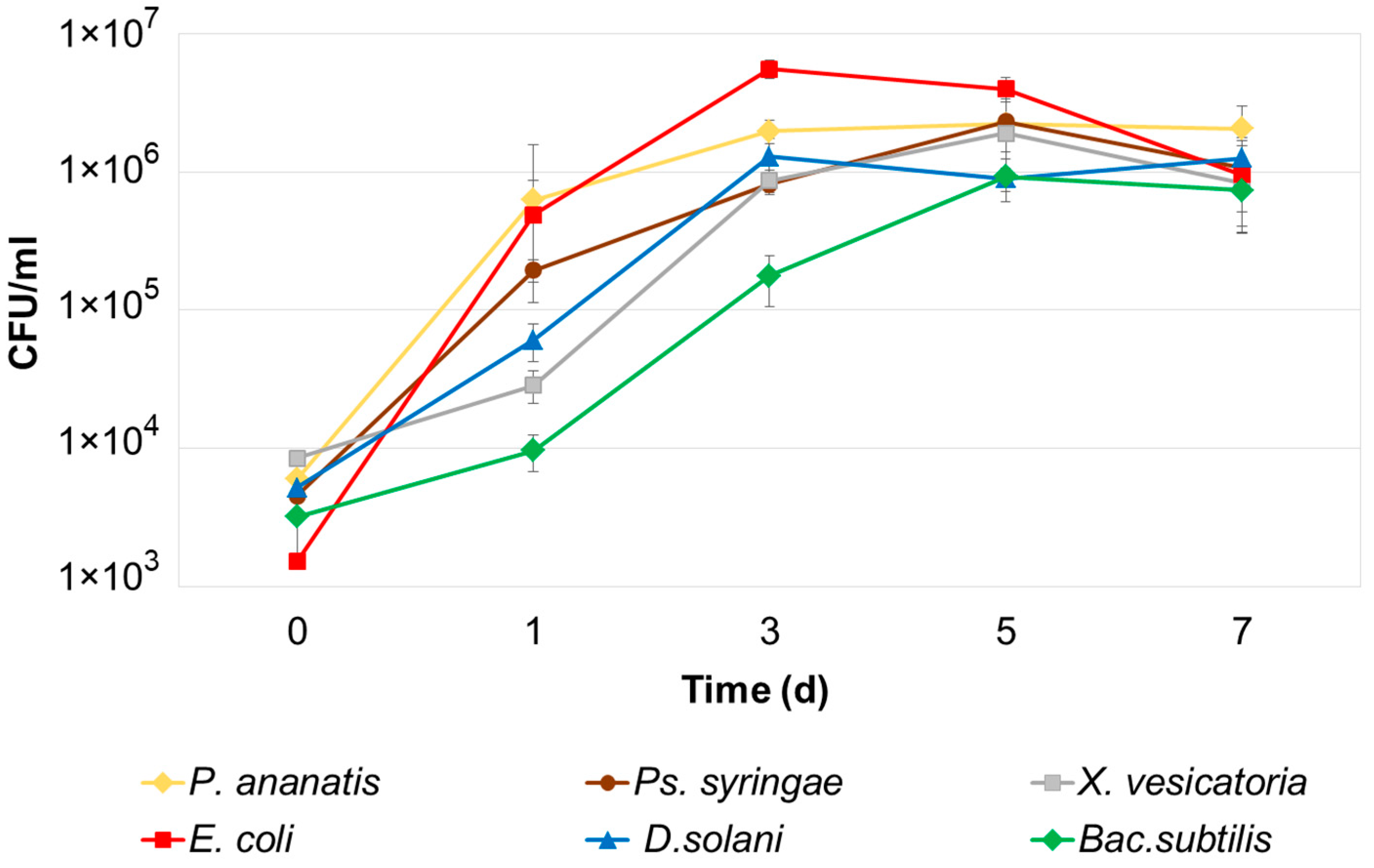
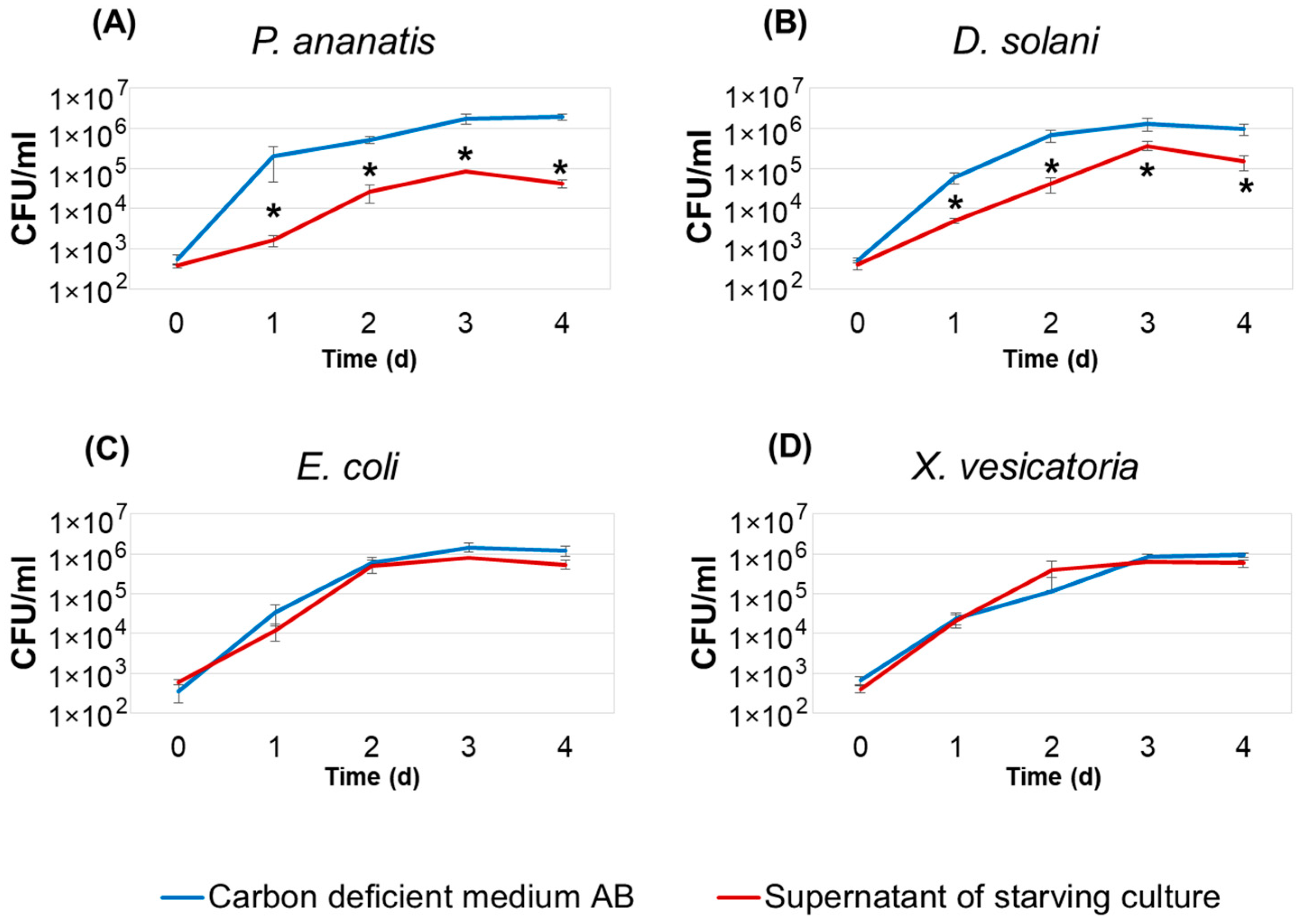
2.5. Effect of Supernatants of Pba Starving Cultures on the Adaptive Proliferation of Bacteria of Different Species
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains, Media and Culture Conditions
4.2. Construction of expI and luxS Deletion Mutants
4.3. Stress Tolerance Assay
4.4. Virulence Assay
4.5. Extraction and Detection of AHL and AI-2
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thorne, S.; Williams, H. Cell density-dependent starvation survival of Rhizobium leguminosarum bv. phaseoli: Identification of the role of an N-acyl homoserine lactone in adaptation to stationary-phase survival. J. Bacteriol. 1999, 181, 981–990. [Google Scholar] [CrossRef] [Green Version]
- Williams, P.; Cámara, M. Quorum sensing and environmental adaptation in Pseudomonas aeruginosa: A tale of regulatory networks and multifunctional signal molecules. Curr. Opin. Microbiol. 2009, 12, 182–191. [Google Scholar] [CrossRef] [PubMed]
- Patterson, A.; Jackson, S.; Taylor, C.; Evans, G.; Salmond, G.; Przybilski, R.; Staals, R.; Fineran, P. Quorum sensing controls adaptive immunity through the regulation of multiple CRISPR-Cas systems. Mol. Cell 2016, 64, 1102–1108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Qin, Q.; Defoirdt, T. Does quorum sensing interference affect the fitness of bacterial pathogens in the real world? Environ. Microbiol. 2018, 20, 3918–3926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Striednig, B.; Hilbi, H. Bacterial quorum sensing and phenotypic heterogeneity: How the collective shapes the individual. Trends Microbiol. 2022, 30, 379–389. [Google Scholar] [CrossRef]
- Gorshkov, V.; Petrova, O.; Gogoleva, N.; Gogolev, Y. Cell-to-cell communication in the populations of enterobacterium Erwinia carotovora ssp. atroseptica SCRI1043 during adaptation to stress conditions. FEMS Microbiol. Immunol. 2010, 59, 378–385. [Google Scholar] [CrossRef] [Green Version]
- Petrova, O.; Gorshkov, V.; Daminova, A.; Ageeva, M.; Moleleki, L.; Gogolev, Y. Stress response in Pectobacterium atrosepticum SCRI1043 under starvation conditions: Adaptive reactions at a low population density. Res. Microbiol. 2014, 165, 119–127. [Google Scholar] [CrossRef]
- Põllumaa, L.; Alamäe, T.; Mäe, A. Quorum sensing and expression of virulence in pectobacteria. Sensors 2012, 12, 3327–3349. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, A.; Cui, Y.; Chakrabarty, P.; Chatterjee, A. Regulation of motility in Erwinia carotovora subsp. carotovora: Quorum-sensing signal controls FlhDC, the global regulator of flagellar and exoprotein genes, by modulating the production of RsmA, an RNA-binding protein. MPMI 2010, 23, 1316–1323. [Google Scholar] [CrossRef] [Green Version]
- Moleleki, L.; Pretorius, R.; Tanui, C.; Mosina, G.; Theron, J. A quorum sensing-defective mutant of Pectobacterium carotovorum ssp. brasiliense 1692 is attenuated in virulence and unable to occlude xylem tissue of susceptible potato plant stems. Mol. Plant Pathol. 2017, 18, 32–44. [Google Scholar] [CrossRef] [Green Version]
- Baltenneck, J.; Reverchon, S.; Hommais, F. Quorum sensing regulation in phytopathogenic bacteria. Microorganisms 2021, 9, 239. [Google Scholar] [CrossRef]
- Rezzonico, F.; Smits, T.; Duffy, B. Detection of AI-2 receptors in genomes of Enterobacteriaceae suggests a role of type-2 quorum sensing in closed ecosystems. Sensors 2012, 12, 6645–6665. [Google Scholar] [CrossRef] [Green Version]
- Pereira, C.; Thompson, J.; Xavier, K. AI-2-mediated signalling in bacteria. FEMS Microbiol. Rev. 2013, 37, 156–181. [Google Scholar] [CrossRef] [Green Version]
- Long, T.; Tu, K.; Wang, Y.; Mehta, P.; Ong, N.; Bassler, B.; Wingreen, N. Quantifying the integration of quorum-sensing signals with single-cell resolution. PLoS Biol. 2009, 7, e1000068. [Google Scholar] [CrossRef] [Green Version]
- Cornforth, D.; Popat, R.; McNally, L.; Gurney, J.; Scott-Phillips, T.; Ivens, A.; Diggle, S.; Brown, S. Combinatorial quorum sensing allows bacteria to resolve their social and physical environment. Proc. Natl. Acad. Sci. USA 2014, 111, 4280–4284. [Google Scholar] [CrossRef] [Green Version]
- Anetzberger, C.; Reiger, M.; Fekete, A.; Schell, U.; Stambrau, N.; Plener, L.; Kopka, J.; Schmitt-Kopplin, P.; Hilbi, H.; Jung, K. Autoinducers act as biological timers in Vibrio harveyi. PLoS ONE 2012, 7, e48310. [Google Scholar] [CrossRef] [Green Version]
- Teng, S.; Schaffer, J.; Tu, K.; Mehta, P.; Lu, W.; Ong, N.; Bassler, B.; Wingreen, N. Active regulation of receptor ratios controls integration of quorum-sensing signals in Vibrio harveyi. Mol. Syst. Biol. 2011, 7, 491. [Google Scholar] [CrossRef]
- Fan, G.; Bressloff, P. Modeling the role of feedback in the adaptive response of bacterial quorum sensing. Bull. Math. Biol. 2019, 81, 1479–1505. [Google Scholar] [CrossRef] [PubMed]
- Crépin, A.; Barbey, C.; Beury-Cirou, A.; Hélias, V.; Taupin, L.; Reverchon, S.; Nasser, W.; Faure, D.; Dufour, A.; Orange, N.; et al. Quorum sensing signaling molecules produced by reference and emerging soft-rot bacteria (Dickeya and Pectobacterium spp.). PLoS ONE 2012, 7, e35176. [Google Scholar] [CrossRef] [PubMed]
- Potrykus, M.; Hugouvieux-Cotte-Pattat, N.; Lojkowska, E. Interplay of classic Exp and specific Vfm quorum sensing systems on the phenotypic features of Dickeya solani strains exhibiting different virulence levels. Mol. Plant Pathol. 2018, 19, 1238–1251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papenfort, K.; Bassler, B.L. Quorum sensing signal–response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 2016, 14, 576–588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abisado, R.G.; Benomar, S.; Klaus, J.R.; Dandekar, A.A.; Chandler, J.R. Bacterial quorum sensing and microbial community interactions. MBio 2018, 9, e02331-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mukherjee, S.; Bassler, B.L. Bacterial quorum sensing in complex and dynamically changing environments. Nat. Rev. Microbiol. 2019, 17, 371–382. [Google Scholar] [CrossRef]
- Riedel, K.; Hentzer, M.; Geisenberger, O.; Huber, B.; Steidle, A.; Wu, H.; Høiby, N.; Givskov, M.; Molin, S.; Eberl, L. N-acylhomoserine-lactone-mediated communication between Pseudomonas aeruginosa and Burkholderia cepacia in mixed biofilms. Microbiol. 2001, 147, 3249–3262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hosni, T.; Moretti, C.; Devescovi, G.; Suarez-Moreno, Z.R.; Fatmi, M.B.; Guarnaccia, C.; Pongor, S.; Onofri, A.; Buonaurio, R.; Venturi, V. Sharing of quorum-sensing signals and role of interspecies communities in a bacterial plant disease. ISME J. 2011, 5, 1857–1870. [Google Scholar] [CrossRef] [PubMed]
- Valente, R.S.; Nadal-Jimenez, P.; Carvalho, A.F.; Vieira, F.J.; Xavier, K.B. Signal integration in quorum sensing enables cross-species induction of virulence in Pectobacterium wasabiae. MBio 2017, 8, e00398-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wellington, S.; Greenberg, E.P. Quorum sensing signal selectivity and the potential for interspecies cross talk. MBio 2019, 10, e00146-19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaniga, K.; Delor, I.; Cornelis, G.R. A wide-host-range suicide vector for improving reverse genetics in gram-negative bacteria: Inactivation of the blaA gene of Yersinia enterocolitica. Gene 1991, 109, 137–141. [Google Scholar] [CrossRef]
- Quan, J.; Tian, J. Circular polymerase extension cloning of complex gene libraries and pathways. PLoS ONE 2009, 4, e6441. [Google Scholar] [CrossRef] [Green Version]
- Pierson, E.; Wood, D.; Cannon, J.; Blachere, F.; Pierson, L., III. Interpopulation signaling via N-acyl-homoserine lactones among bacteria in the wheat rhizosphere. MPMI 1998, 11, 1078–1084. [Google Scholar] [CrossRef] [Green Version]
- Lindsay, A.; Ahmer, B. Effect of sdiA on biosensors of N-acylhomoserine lactones. J. Bacteriol. 2005, 187, 5054–5058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Surette, M.; Bassler, B. Quorum sensing in Escherichia coli and Salmonella typhimurium. Proc. Natl. Acad. Sci. USA 1998, 95, 7046–7050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bassler, B.; Wright, M.; Silverman, M. Multiple signalling systems controlling expression of luminescence in Vibrio harveyi: Sequence and function of genes encoding a second sensory pathway. Mol. Microbiol. 1994, 13, 273–286. [Google Scholar] [CrossRef] [PubMed]
- Bell, K.S.; Sebaihia, M.; Pritchard, L.; Holden, M.T.G.; Hyman, L.J.; Holeva, M.C.; Thomson, N.R.; Bentley, S.D.; Churcher, L.J.C.; Mungall, K.; et al. Genome sequence of the enterobacterial phytopathogen Erwinia carotovora subsp. atroseptica and characterization of virulence factors. Proc. Natl. Acad. Sci. USA 2004, 101, 11105–11110. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herrero, M.; de Lorenzo, V.; Timmis, K.N. Transposon vectors containing non-antibiotic resistance selection markers for cloning and stable chromosomal insertion of foreign genes in gram-negative bacteria. J. Bacteriol. 1990, 172, 6557–6567. [Google Scholar] [CrossRef] [Green Version]
- Grinter, N.J. A broad-host-range cloning vector transposable to various replicons. Gene 1983, 21, 133–143. [Google Scholar] [CrossRef]
- De Maayer, P.; Chan, W.Y.; Venter, S.N.; Toth, I.K.; Birch, P.R.J.; Joubert, F.; Coutinho, T.A. The genome sequence of Pantoea ananatis LMG20103, the causative agent of Eucalyptus blight and dieback. J. Bacteriol. 2010, 192, 2936–2937. [Google Scholar] [CrossRef] [Green Version]
- Van der Wolf, J.M.; Nijhuis, E.H.; Kowalewska, M.J.; Saddler, G.S.; Parkinson, N.; Elphinstone, J.G.; Pritchard, L.; Toth, I.K.; Lojkowska, E.; Potrykus, M.; et al. Dickeya solani sp. nov., a pectinolytic plant-pathogenic bacterium isolated from potato (Solanum tuberosum). Int. J. Syst. Evol. Microbiol. 2014, 64, 768–774. [Google Scholar] [CrossRef] [Green Version]
- Vauterin, L.; Hoste, B.; Kersters, K.; Swings, J. Reclassification of Xanthomonas. Int. J. Syst. Bacteriol. 1995, 45, 472–489. [Google Scholar] [CrossRef] [Green Version]
- Buell, C.R.; Joardar, V.; Lindeberg, M.; Selengut, J.; Paulsen, I.T.; Gwinn, M.L.; Dodson, R.J.; Deboy, R.T.; Durkin, A.S.; Kolonay, J.F.; et al. The complete genome sequence of the Arabidopsis and tomato pathogen Pseudomonas syringae pv. tomato DC3000. Proc. Natl. Acad. Sci. USA 2003, 100, 10181–10186. [Google Scholar] [CrossRef] [Green Version]
- Ansorg, R.; Zappel, H.; Thomssen, R. Bedeutung des Nachweises antibakterieller Stoffe im Urin für die bakteriologische Diagnostik und die Kontrolle der Chemotherapie von Harnwegsinfektionen. Zentralbl. Bakteriol. Parasitenkd. Orig. Reihe. 1975, 230, 492–507. [Google Scholar]
- Datsenko, K.A.; Wanner, B.L. One-step inactivation of chromosomal genes in Escherichia coli K-12 using PCR products. Proc. Natl. Acad. Sci. USA 2000, 97, 6640–6645. [Google Scholar] [CrossRef] [PubMed] [Green Version]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Petrova, O.; Parfirova, O.; Gogoleva, N.; Vorob’ev, V.; Gogolev, Y.; Gorshkov, V. The Role of Intercellular Signaling in the Regulation of Bacterial Adaptive Proliferation. Int. J. Mol. Sci. 2023, 24, 7266. https://doi.org/10.3390/ijms24087266
Petrova O, Parfirova O, Gogoleva N, Vorob’ev V, Gogolev Y, Gorshkov V. The Role of Intercellular Signaling in the Regulation of Bacterial Adaptive Proliferation. International Journal of Molecular Sciences. 2023; 24(8):7266. https://doi.org/10.3390/ijms24087266
Chicago/Turabian StylePetrova, Olga, Olga Parfirova, Natalia Gogoleva, Vladimir Vorob’ev, Yuri Gogolev, and Vladimir Gorshkov. 2023. "The Role of Intercellular Signaling in the Regulation of Bacterial Adaptive Proliferation" International Journal of Molecular Sciences 24, no. 8: 7266. https://doi.org/10.3390/ijms24087266
APA StylePetrova, O., Parfirova, O., Gogoleva, N., Vorob’ev, V., Gogolev, Y., & Gorshkov, V. (2023). The Role of Intercellular Signaling in the Regulation of Bacterial Adaptive Proliferation. International Journal of Molecular Sciences, 24(8), 7266. https://doi.org/10.3390/ijms24087266