


The Molecular Epidemiology of Pneumococcal Strains Isolated from the Nasopharynx of Preschool Children 3 Years after the Introduction of the PCV Vaccination Program in Poland

 ,
,  ,
, 
Abstract
1. Introduction
2. Results
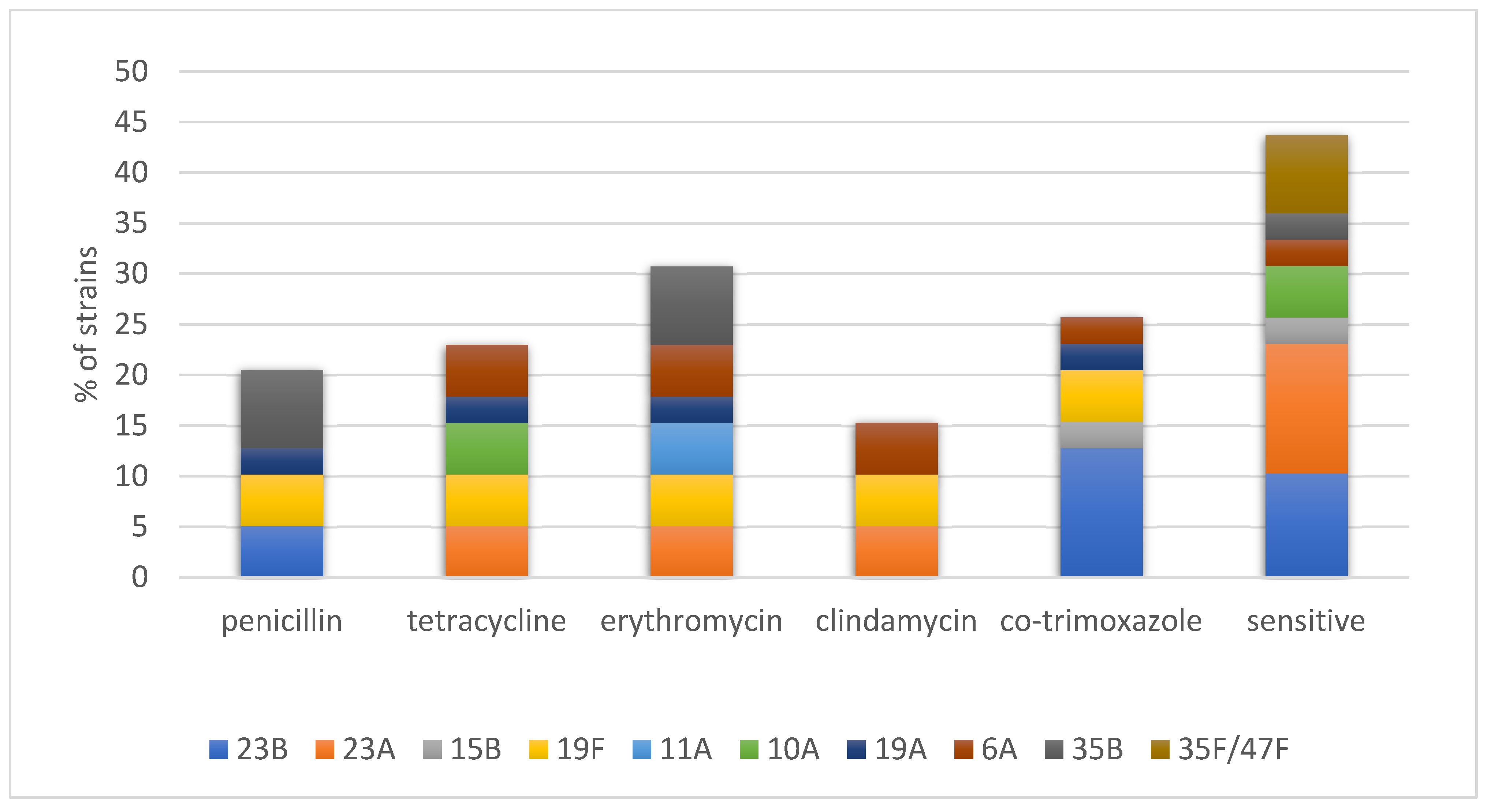
2.1. Resistance Genes and Presumptive Transposons
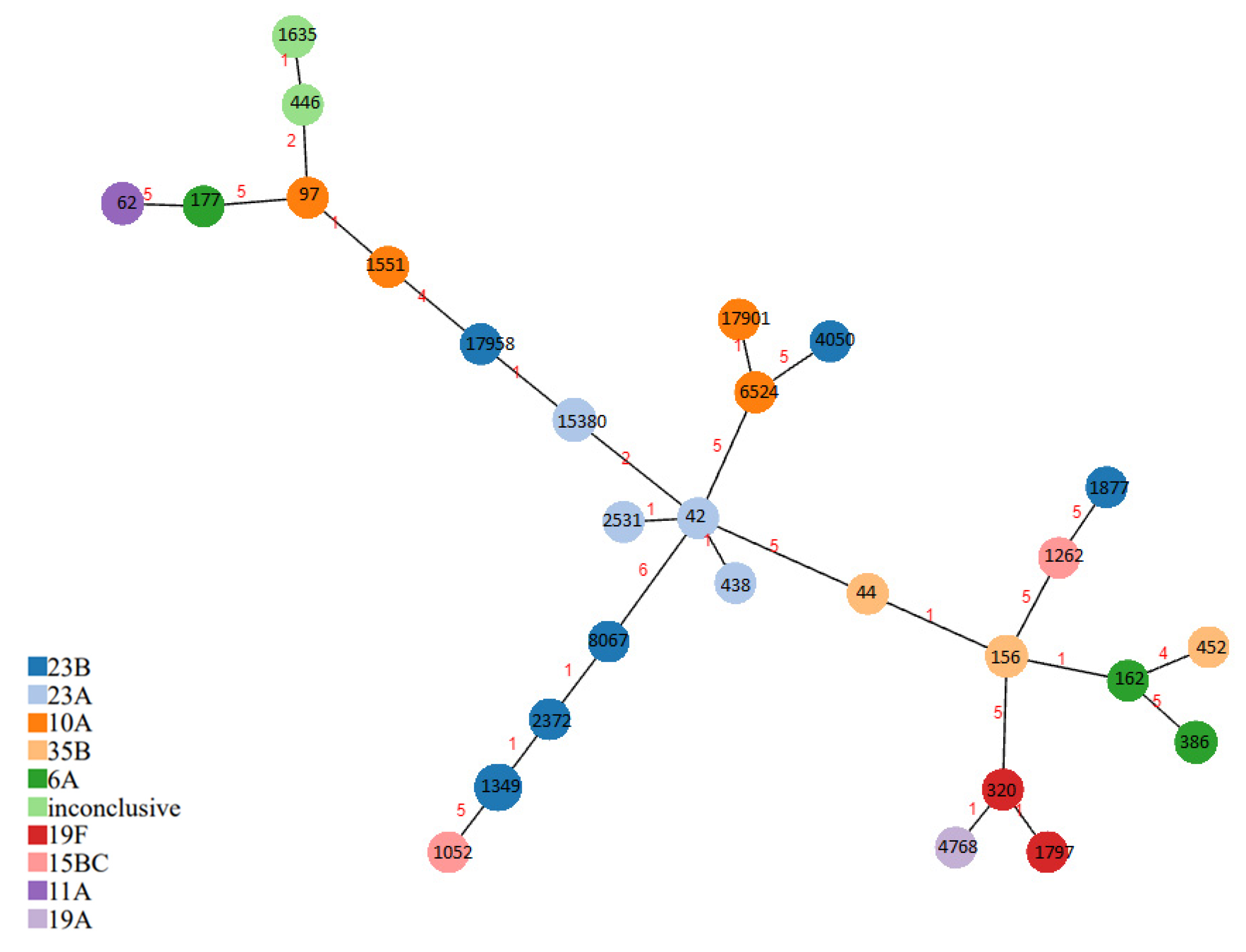
2.2. Multilocus Sequence Typing (MLST) and PMEN Clones
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Detection of Resistance Genes, Transposons, and Pili
4.3. Multilocus Sequence Typing (MLST)
4.4. Cluster Analysis and PMEN Database
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- De Lencastre, H.; Tomasz, A. From Ecological Reservoir to Disease: The Nasopharynx, Day-Care Centres and Drug-Resistant Clones of Streptococcus pneumoniae. J. Antimicrob. Chemother. 2002, 50 (Suppl. S2), 75–81. [Google Scholar] [CrossRef]
- Bogaert, D.; De Groot, R.; Hermans, P.W.M. Streptococcus pneumoniae Colonisation: The Key to Pneumococcal Disease. Lancet Infect. Dis. 2004, 4, 144–154. [Google Scholar] [CrossRef]
- Mitchell, A.M.; Mitchell, T.J. Streptococcus pneumoniae: Virulence Factors and Variation. Clin. Microbiol. Infect. 2010, 16, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Korona-Głowniak, I.; Malm, A. Streptococcus pneumoniae–colonization and pneumococcal disease. Post. Mikrobiol. 2015, 54, 309–319. Available online: https://www.pm.microbiology.pl (accessed on 20 January 2023).
- Dzaraly, N.D.; Muthanna, A.; Mohd Desa, M.N.; Taib, N.M.; Masri, S.N.; Rahman, N.I.A.; Suhaili, Z.; Tuan Soh, T.S.; Abdullah, F.H. Pilus Islets and the Clonal Spread of Piliated Streptococcus pneumoniae: A Review. Int. J. Med. Microbiol. 2020, 310, 151449. [Google Scholar] [CrossRef]
- 2017 Poland Introduced 10-Valent Pneumococcal Vaccine in the Immunization Schedule. Szczepienia. Info EN. Available online: http://szczepieniainfo.testa.com.pl/en/stories/introduction_of_pcv_2016/ (accessed on 22 August 2018).
- Korona-Glowniak, I.; Malm, A. Characteristics of Streptococcus pneumoniae Strains Colonizing Upper Respiratory Tract of Healthy Preschool Children in Poland. Sci. World J. 2012, 2012, 732901. [Google Scholar] [CrossRef] [PubMed]
- Korona-Glowniak, I.; Niedzielski, A.; Malm, A. Upper Respiratory Colonization by Streptococcus pneumoniae in Healthy Pre-School Children in South-East Poland. Int. J. Pediatr. Otorhinolaryngol. 2011, 75, 1529–1534. [Google Scholar] [CrossRef] [PubMed]
- Niedzielski, A.; Korona-Glowniak, I.; Malm, A. High Prevalence of Streptococcus pneumoniae in Adenoids and Nasopharynx in Preschool Children with Recurrent Upper Respiratory Tract Infections in Poland—Distribution of Serotypes and Drug Resistance Patterns. Med. Sci. Monit. 2013, 19, 54–60. [Google Scholar] [CrossRef]
- Korona-Glowniak, I.; Niedzielski, A.; Kosikowska, U.; Grzegorczyk, A.; Malm, A. Nasopharyngeal vs. Adenoid Cultures in Children Undergoing Adenoidectomy: Prevalence of Bacterial Pathogens, Their Interactions and Risk Factors. Epidemiol. Infect. 2015, 143, 821–830. [Google Scholar] [CrossRef] [PubMed]
- Wróbel-Pawelczyk, I.; Ronkiewicz, P.; Wanke-Rytt, M.; Rykowska, D.; Górska-Kot, A.; Włodkowska, K.; Topczewska-Cabanek, A.; Jackowska, T.; Chruszcz, J.; Marchut, W.; et al. Pneumococcal Carriage in Unvaccinated Children at the Time of Vaccine Implementation into the National Immunization Program in Poland. Sci. Rep. 2022, 12, 5858. [Google Scholar] [CrossRef] [PubMed]
- Wyres, K.L.; Lambertsen, L.M.; Croucher, N.J.; McGee, L.; von Gottberg, A.; Liñares, J.; Jacobs, M.R.; Kristinsson, K.G.; Beall, B.W.; Klugman, K.P.; et al. Pneumococcal Capsular Switching: A Historical Perspective. J. Infect. Dis. 2013, 207, 439–449. [Google Scholar] [CrossRef] [PubMed]
- Weinberger, D.M.; Malley, R.; Lipsitch, M. Serotype Replacement in Disease after Pneumococcal Vaccination. Lancet 2011, 378, 1962–1973. [Google Scholar] [CrossRef] [PubMed]
- Knutsen, E.; Johnsborg, O.; Quentin, Y.; Claverys, J.-P.; Håvarstein, L.S. BOX Elements Modulate Gene Expression in Streptococcus pneumoniae: Impact on the Fine-Tuning of Competence Development. J. Bacteriol. 2006, 188, 8307–8312. [Google Scholar] [CrossRef]
- Yahiaoui, R.Y.; Bootsma, H.J.; den Heijer, C.D.J.; Pluister, G.N.; John Paget, W.; Spreeuwenberg, P.; Trzcinski, K.; Stobberingh, E.E. Distribution of Serotypes and Patterns of Antimicrobial Resistance among Commensal Streptococcus pneumoniae in Nine European Countries. BMC Infect. Dis. 2018, 18, 440. [Google Scholar] [CrossRef]
- Harrison, O.B.; Brueggemann, A.B.; Caugant, D.A.; van der Ende, A.; Frosch, M.; Gray, S.; Heuberger, S.; Krizova, P.; Olcen, P.; Slack, M.; et al. Molecular Typing Methods for Outbreak Detection and Surveillance of Invasive Disease Caused by Neisseria Meningitidis, Haemophilus Influenzae and Streptococcus pneumoniae, a Review. Microbiology 2011, 157 Pt 8, 2181–2195. [Google Scholar] [CrossRef] [PubMed]
- Rice, L.B. Tn916 Family Conjugative Transposons and Dissemination of Antimicrobial Resistance Determinants. Antimicrob. Agents Chemother. 1998, 42, 1871–1877. [Google Scholar] [CrossRef] [PubMed]
- Del Grosso, M.; Camilli, R.; Iannelli, F.; Pozzi, G.; Pantosti, A. The Mef(E)-Carrying Genetic Element (Mega) of Streptococcus pneumoniae: Insertion Sites and Association with Other Genetic Elements. Antimicrob. Agents Chemother. 2006, 50, 3361–3366. [Google Scholar] [CrossRef]
- Del Grosso, M.; Northwood, J.G.E.; Farrell, D.J.; Pantosti, A. The Macrolide Resistance Genes Erm(B) and Mef(E) Are Carried by Tn2010 in Dual-Gene Streptococcus pneumoniae Isolates Belonging to Clonal Complex CC271. Antimicrob. Agents Chemother. 2007, 51, 4184–4186. [Google Scholar] [CrossRef]
- Kielbik, K.; Pietras, A.; Jablonska, J.; Bakiera, A.; Borek, A.; Niedzielska, G.; Grzegorczyk, M.; Grywalska, E.; Korona-Glowniak, I. Impact of Pneumococcal Vaccination on Nasopharyngeal Carriage of Streptococcus pneumoniae and Microbiota Profiles in Preschool Children in South East Poland. Vaccines 2022, 10, 791. [Google Scholar] [CrossRef]
- Korona-Glowniak, I.; Maj, M.; Siwiec, R.; Niedzielski, A.; Malm, A. Molecular Epidemiology of Streptococcus pneumoniae Isolates from Children with Recurrent Upper Respiratory Tract Infections. PLoS ONE 2016, 11, e0158909. [Google Scholar] [CrossRef]
- Korona-Glowniak, I.; Zychowski, P.; Siwiec, R.; Mazur, E.; Niedzielska, G.; Malm, A. Resistant Streptococcus pneumoniae Strains in Children with Acute Otitis Media–High Risk of Persistent Colonization after Treatment. BMC Infect. Dis. 2018, 18, 478. [Google Scholar] [CrossRef]
- Paton, J.C.; Trappetti, C. Streptococcus pneumoniae Capsular Polysaccharide. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef] [PubMed]
- Korona-Glowniak, I.; Siwiec, R.; Malm, A. Resistance Determinants and Their Association with Different Transposons in the Antibiotic-Resistant Streptococcus pneumoniae. Biomed Res. Int. 2015, 2015, 836496. [Google Scholar] [CrossRef] [PubMed]
- Izdebski, R.; Sadowy, E.; Fiett, J.; Grzesiowski, P.; Gniadkowski, M.; Hryniewicz, W. Clonal Diversity and Resistance Mechanisms in Tetracycline-Nonsusceptible Streptococcus pneumoniae Isolates in Poland. Antimicrob. Agents Chemother. 2007, 51, 1155–1163. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Akdoğan Kittana, F.N.; Mustak, I.B.; Hascelik, G.; Saricam, S.; Gurler, N.; Diker, K.S. Erythromycin-Resistant Streptococcus pneumoniae: Phenotypes, Genotypes, Transposons and Pneumococcal Vaccine Coverage Rates. J. Med. Microbiol. 2019, 68, 874–881. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Cong, Z.; Huang, W.; Li, C. Molecular Characterization of Streptococcus pneumoniae Isolated from Pediatric Patients in Shanghai, China. Pediatr. Pulmonol. 2020, 55, 2135–2141. [Google Scholar] [CrossRef]
- Chancey, S.T.; Agrawal, S.; Schroeder, M.R.; Farley, M.M.; Tettelin, H.; Stephens, D.S. Composite Mobile Genetic Elements Disseminating Macrolide Resistance in Streptococcus pneumoniae. Front. Microbiol. 2015, 6, 26. [Google Scholar] [CrossRef]
- Cochetti, I.; Tili, E.; Vecchi, M.; Manzin, A.; Mingoia, M.; Varaldo, P.E.; Montanari, M.P. New Tn916-Related Elements Causing Erm(B)-Mediated Erythromycin Resistance in Tetracycline-Susceptible Pneumococci. J. Antimicrob. Chemother. 2007, 60, 127–131. [Google Scholar] [CrossRef]
- Hjálmarsdóttir, M.; Pétursdóttir, B.; Erlendsdóttir, H.; Haraldsson, G.; Kristinsson, K. Prevalence of Pilus Genes in Pneumococci Isolated from Healthy Preschool Children in Iceland: Association with Vaccine Serotypes and Antibiotic Resistance. J. Antimicrob. Chemother. 2015, 70, 2203–2208. [Google Scholar] [CrossRef][Green Version]
- Mills, R.O.; Abdullah, M.R.; Akwetey, S.A.; Sappor, D.C.; Cole, I.; Baffuor-Asare, M.; Bolivar, J.A.; Gámez, G.; van der Linden, M.P.G.; Hammerschmidt, S. Post-Vaccination Streptococcus pneumoniae Carriage and Virulence Gene Distribution among Children Less Than Five Years of Age, Cape Coast, Ghana. Microorganisms 2020, 8, 1987. [Google Scholar] [CrossRef]
- Regev-Yochay, G.; Hanage, W.P.; Trzcinski, K.; Rifas-Shiman, S.L.; Lee, G.; Bessolo, A.; Huang, S.S.; Pelton, S.I.; McAdam, A.J.; Finkelstein, J.A.; et al. Re-Emergence of the Type 1 Pilus among Streptococcus pneumoniae Isolates in Massachusetts, USA. Vaccine 2010, 28, 4842–4846. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Aguiar, S.I.; Melo-Cristino, J.; Ramirez, M. Use of the 13-Valent Conjugate Vaccine Has the Potential to Eliminate Pilus Carrying Isolates as Causes of Invasive Pneumococcal Disease. Vaccine 2012, 30, 5487–5490. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchiya, M.; Urushibara, N.; Aung, M.S.; Shinagawa, M.; Takahashi, S.; Kobayashi, N. Serotype distribution, antimicrobial resistance and prevalence of pilus islets in pneumococci following the use of conjugate vaccines. J. Med. Microbiol. 2017, 66, 643–650. [Google Scholar] [CrossRef]
- Regev-Yochay, G.; Jaber, H.; Hamdan, A.; Daana, M.; Nammouz, H.; Thalji, A.; Jaar, F.; Abdeen, Z.; Rubin, C.; Huppert, A.; et al. Vaccine escape of piliated Streptococcus pneumoniae strains. Vaccine 2016, 34, 2787–2792. [Google Scholar] [CrossRef]
- Enright, M.C.; Spratt, B.G. A Multilocus Sequence Typing Scheme for Streptococcus pneumoniae: Identification of Clones Associated with Serious Invasive Disease. Microbiology 1998, 144 Pt 11, 3049–3060. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Clonal Group | Sequence Types (n) | Associated PMEN Clone | Serotype (n) | Resistance Pattern | Resistance Genes (n) | Transposon Genes | Presumptive Transposon | Pili |
|---|---|---|---|---|---|---|---|---|
| 1 | 42 * (1) | 23A | ECCTe | erm(B), tet(M) | Int/xis916 | Tn6002 | ||
| 438 (1) | Tennessee23F-4, DLV | 23A | S | |||||
| 2531 (1) | Tennessee23F-4, DLV | 23A | ECCTe | erm(B), tet(M) | Int/xis916, orf20-19(3.7 kb) | Tn6002 | ||
| 15380 (3) | 23A (3) | S (3) | PI-1 + PI-2 (1) | |||||
| 17958 (1) | 23A | S | PI-2 | |||||
| 2 | 44 (1) | Spain 9V-3, SVL | 35B | PE | mef(E) | mel | MEGA | PI-1 |
| 156 * (2) | Spain 9V-3 | 35B (2) | PE (2) | mef(E) (2) | mel | MEGA (2) | PI-1 (2) | |
| 162 (1) | Spain 9V-3, SVL | 6A | S | PI-1 | ||||
| 3 | 97 (1) | 10A | S | |||||
| 446 (1) | 35F/47F | S | ||||||
| 1551 (1) | 10A | S | ||||||
| 1635 (2) | 35F/47F (2) | S (2) | ||||||
| 4 | 320 * (1) | 19F | PECCTeSXT | erm(B), tet(M) | int/xis916, orf20-19(3.7 kb) | Tn6002 | PI-1 + PI-2 | |
| 4768 (1) | 19A | PETeSXT | mef(E), tet(M) | int/xis916, orf20-29(3.7 kb) | Tn2009 | PI-1 + PI-2 | ||
| 17907 (1) | Taiwan 19F-14, DLV | 19F | PECCTeSXT | erm(B), tet(M) | int/xis916, orf20-19(3.7 kb) | Tn6002 | PI-1 + PI-2 | |
| 5 | 1349 (6) | Colombia 23F-26, DLV | 23B (6) | SXT (4) S (2) | PI-1 (1), PI-2 (1) | |||
| 2372 * (1) | 23B | P | PI-1 + PI-2 | |||||
| 8067 (1) | 23B | PSXT | PI-2 | |||||
| 6 | 6524 (1) | 10A | Te | tet(M) | int/xis916, orf20-19(0.8kb) | Tn916 | ||
| 17901 (1) | 10A | Te | tet(M) | int/xis916, orf20-19(0.8kb) | Tn916 | |||
| Singletons | 62 (2) | Netherlands 8-33, DLV | 11A (2) | E (2) | mef(E) (2) | mel | MEGA(2) | PI-2 |
| 177 (1) | 6A | SXT | PI-1 + PI-2 | |||||
| 386 (2) | Poland 6B-20, DLV | 6A (2) | ECCTe (2) | erm(B), tet(M) (2) | int/xis916, orf20-19(3.7 kb) | Tn6002 (2) | ||
| 452 (1) | 35B | S | ||||||
| 1025 (1) | 15B | SXT | ||||||
| 1262 (1) | 15B | S | PI-2 | |||||
| 1877 (1) | Greece 21-30, DLV | 23B | S | |||||
| 4050 (1) | 23B | S |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kielbik, K.; Grywalska, E.; Glowniak, A.; Mielnik-Niedzielska, G.; Korona-Glowniak, I. The Molecular Epidemiology of Pneumococcal Strains Isolated from the Nasopharynx of Preschool Children 3 Years after the Introduction of the PCV Vaccination Program in Poland. Int. J. Mol. Sci. 2023, 24, 7883. https://doi.org/10.3390/ijms24097883
Kielbik K, Grywalska E, Glowniak A, Mielnik-Niedzielska G, Korona-Glowniak I. The Molecular Epidemiology of Pneumococcal Strains Isolated from the Nasopharynx of Preschool Children 3 Years after the Introduction of the PCV Vaccination Program in Poland. International Journal of Molecular Sciences. 2023; 24(9):7883. https://doi.org/10.3390/ijms24097883
Chicago/Turabian StyleKielbik, Karolina, Ewelina Grywalska, Andrzej Glowniak, Grażyna Mielnik-Niedzielska, and Izabela Korona-Glowniak. 2023. "The Molecular Epidemiology of Pneumococcal Strains Isolated from the Nasopharynx of Preschool Children 3 Years after the Introduction of the PCV Vaccination Program in Poland" International Journal of Molecular Sciences 24, no. 9: 7883. https://doi.org/10.3390/ijms24097883
APA StyleKielbik, K., Grywalska, E., Glowniak, A., Mielnik-Niedzielska, G., & Korona-Glowniak, I. (2023). The Molecular Epidemiology of Pneumococcal Strains Isolated from the Nasopharynx of Preschool Children 3 Years after the Introduction of the PCV Vaccination Program in Poland. International Journal of Molecular Sciences, 24(9), 7883. https://doi.org/10.3390/ijms24097883