Abstract
Infectious bursal disease virus (IBDV) is an immunosuppressive pathogen causing enormous economic losses to the poultry industry across the globe. As a double-stranded RNA virus, IBDV undergoes genetic mutation or recombination in replication during circulation among flocks, leading to the generation and spread of variant or recombinant strains. In particular, the recent emergence of variant IBDV causes severe immunosuppression in chickens, affecting the efficacy of other vaccines. It seems that the genetic mutation of IBDV during the battle against host response is an effective strategy to help itself to survive. Therefore, a comprehensive understanding of the viral genome diversity will definitely help to develop effective measures for prevention and control of infectious bursal disease (IBD). In recent years, considerable progress has been made in understanding the relation of genetic mutation and genomic recombination of IBDV to its pathogenesis using the reverse genetic technique. Therefore, this review focuses on our current genetic insight into the IBDV’s genetic typing and viral genomic variation.
1. Introduction
Infectious bursal disease virus (IBDV) is a globally prevalent chicken immunosuppressive virus that causes severe immunosuppression in infected chickens, leading to increased susceptibility to other pathogens or even death [1,2,3]. Although there are two serotypes of IBDV, only serotypeI causes disease in poultry. SerotypeII was isolated from turkeys and is not pathogenic to chickens [3,4]. Under natural conditions, IBDV can infect all breeds of chickens, causing huge economic losses to the poultry industry worldwide [5,6,7,8,9,10].
Due to the unique bi-segmented double-stranded RNA genome and its high error rate of viral RNA-dependent RNA polymerase (RdRp) [3,11,12], IBDV is naturally prone to varying degrees of genomic mutation or recombination, leading to the emergence and spread of new mutant or recombinant strains in chickens [13,14,15]. Several main pathogenic types of IBDV have been identified, including classical IBDV (cIBDV), variant IBDV (varIBDV), very virulent IBDV (vvIBDV), attenuated IBDV, and novel variant IBDV (nVarIBDV), which differ in pathogenicity and antigenicity [7,16,17,18]. Nonetheless, there is no clear standard for typing the IBDV genome, and the description of the genotype of IBDV has become complex and imprecise as novel strains emerge [6,19,20]. Consequently, traditional classification methods based on pathogenicity and antigenicity need to be modified. Recently, several improved genotyping schemes have been proposed that would greatly facilitate the genetics and molecular epidemiological investigation of IBDV [21,22,23,24].
Currently, the vaccination of chickens with inactivated and live attenuated vaccines are commonly-used clinical methods to control IBD [25,26,27,28]. However, the high mutation rate of IBDV is likely to be the reason for the emergence of mutant viral strains whose antigenicity differs from that of the current commercially-available vaccines [16,29,30,31]. The battle between IBDV and the host is manifested in various ways [32,33,34,35], the most obvious being viral variation at a genomic level that is critical to genetic diversity and immune evasion [14,16,36,37]. Therefore, a comprehensive understanding of the patterns of genomic variations in IBDV is critical for the prevention and control of IBD. In recent years, reverse genetics has emerged as a useful technic to combat IBDV infection. This approach can make true the precise mutations or substitutions in the IBDV genome from a genetic perspective, and many advances have been made in this aspect to explore pathogenesis and develop novel vaccines [38,39,40,41].
This review focuses on the current epidemics and pathogenesis of infections with different IBDV strains, as well as novel approaches to virus classification based on phylogenetic evolution. In addition, we have summarized the current genetic insights into virus-host interactions at a genomic level, as well as the application of reverse genetics to IBDV vaccine development, providing new perspectives for future prevention, control, and vaccine development against IBDV.
2. Epidemics and Pathogenesis of Different IBDV Strains
2.1. Classical IBDV (cIBDV)
In 1957, the original outbreak of IBD occurred in the area of Gumboro, Delaware, USA, where researchers observed a high rate of disease occurrence in chicks [42]. Early symptoms of IBD include diarrhea, loss of appetite, weakness, and even death [42]. The main target organ of the virus is the bursa of Fabricius (BF) of 3–6-week-old chicks, which is characterized by enlarged or hemorrhagic bursa during the first four days, followed by bursal atrophy later in the course of the disease [3,25,43]. The infection eventually leads to lymphocyte failure and destruction of the bursa, which is the main feature of IBD pathogenesis [1,44]. In classical epidemic situations, mortality in diseased chickens may range between 1% and 50%, with significant effects on both broilers and laying hens [3]. In addition to mortality, IBDV also has an immunosuppressive effect, which affects the host's immune response and the efficacy of other vaccinations [3,43,45]. The virus genome consists of two segments of RNAase-resistant, double-stranded RNAs [46]. The complete terminal sequences of IBDV genomic dsRNA have been identified, indicating that different RNA structures may have an impact on the ability of the virus to replicate [47]. As more complete genomic information on IBDV has become available, the correlation between genetic and pathogenic phenotypes among different strains of IBDV could be more precisely assessed [14,17,48,49,50,51].
2.2. Variant IBDV (varIBDV)
In the late 1980s, it was first reported that IBDV variants were identified by virus neutralization tests as serotypeI IBDV with significant antigenic differences from classical strains [37,52]. These antigenically altered strains were collectively referred to as variants IBDV to distinguish them from previous classical IBDV isolates [22,42]. Initially, the variant IBDV-infected chickens were characterized by little or no mortality and no obvious clinical signs, but their bursa and spleen were damaged [29,53,54]. Subsequent studies have shown that amino acid mutations in the hypervariable region (HVR, nt 616–1050, aa 206–350) of VP2 are the major cause of IBDV variants [30,55,56]. The VP2 hypervariable region includes many amino acid residues exposed on the protein surface, and mutations in these residues can lead to a variation in the antigenicity of VP2, allowing the variant to escape from the neutralizing antibodies produced by vaccination against the cIBDV [54,56]. This phenomenon is known as antigenic drift and is the primary reason for antigenic diversity [30,57]. Several mutations in the specific residues (222 T, 249 K, 254 S, 286 I, and 318 D) in the HVR may cause antigenic drift [58,59].
2.3. Very Virulent IBDV (vvIBDV)
At the time the variant IBDV was identified in North America, very virulent IBDV (vvIBDV) emerged in Europe [60,61]. In contrast to the variant strains, vvIBDV typically causes high morbidity and mortality in SPF chickens, resulting in mortality of 50–100%, along with typical signs and lesions. Importantly, vvIBDV can establish infection in the presence of maternal antibodies to the classical strain and cause lesions in immune organs other than the bursa [2,25]. As a highly transmissible virus, vvIBDV has spread rapidly throughout the world, causing significant economic losses to the poultry industry [7,50,62,63,64,65]. However, the emergence of vvIBDV promoted scientific research on the pathogenicity of IBDV infection.
Genetic studies revealed that the emergence of the vvIBDV strains was due to the reassortment of genetic segments, specifically the reassortment of the mutated segment A with the segment B of unknown origin, resulting in a sudden increase in the pathogenicity of the virus [66]. Residues 222 A, 242 I, 256 I, 294 I, and 299 S of the vvIBDV segment A are conserved compared to other strains of IBDV and serve as markers of pathogenicity [2,67,68]. In addition, residues 253 and 284 of VP2 are proposed to be the determinants of cell tropism and major contributors to IBDV virulence [69,70,71]. Phylogenetic analysis has shown that segment B of vvIBDV is distinct and highly conserved [13,58,66]. Additional studies have confirmed that both genomic segments contribute to the high virulence of vvIBDV [72,73,74,75,76]. As the efforts in exploring vvIBDV continue, more critical residues that may be involved in the pathogenicity of the virus have been identified [77,78,79].
2.4. Novel Variant IBDV (nVarIBDV)
Since 2015, China has experienced an outbreak of the novel variant IBDV (nVarIBDV) [6]. These isolated strains have distinct subclusters, indicating genetic evolution in both segments A and B of the viral genome, which rarely occurred in China before [80,81,82,83]. While the vvIBDV strain has historically been the most prevalent with low variability in field transmission [7,17], the recent emergence of nVarIBDV outbreaks in other Asian countries such as Japan [9], South Korea [84], and Malaysia [85] are of concern. It was reported that nVarIBDV does not cause severe clinical signs but causes irreversible damage to the immune organs of chickens, including bursal lesions, spleen swelling and atrophy, and long-term immunosuppression, to a greater extent than the previous varIBDV [6,18,86]. In addition, the new strains can break through the immune protection provided by existing vvIBDV vaccines [84,87,88].
The novel variant spectrum of IBDV strains exhibits significant genetic differences from previously reported IBDV strains. The strains contain multiple amino acid substitutions, with VP2 having typical residues similar to varIBDV (222 T, 249 K, 286 I, and 318 D) [58], as well as other residues such as 221 K, 252 I, and 299 S, and VP1 containing 147 D and 508 K [6]. Although the same amino acid differences have been reported in other studies [89], the relationship between these amino acid substitutions and the antigenicity and pathogenicity of the strains has not been fully investigated. Therefore, further investigations into novel variant IBDV strains are needed to better understand the epidemics of currently circulating IBDV strains.
2.5. Other Strain Types of IBDV
There are several other strain types of IBDV, including attenuated, reassortant, and recombinant strains. Attenuated strains usually refer to those attenuated live vaccine strains made especially for the control of cIBDV or vvIBDV infection and are classified as mild, intermediate, or intermediate-plus based on their attenuation [25]. The immunization of chicks with attenuated vaccine strains has become the primary defense against IBD in young chickens. Although attenuated strains are generally not lethal to chickens, intermediate and intermediate plus attenuated strains can cause varying degrees of bursal damage in vaccinated chickens [26]. However, due to the widespread use of live vaccines, the spread of different strain types among flocks in recent years has led to the increasing emergence of reassortant and recombinant strains of IBDV, such as strains with segment A of vvIBDV and segment B of attenuated strains [76,90,91], segment A of vvIBDV and segment B of serotypeII [75], and segment A of vaccine strains and segment B of vvIBDV [15,92], etc. Homologous recombination between different strains of IBDV has also been reported [58,93,94,95]. Recombination and reassortment between different strains pose new challenges to the prevention and control of IBD and require continued efforts to investigate the genetics and epidemics of IBDV.
3. Classification of IBDV Based on Genomic Phylogenetic Evolution
3.1. Recent Scheme for IBDV Genomic Classification
The transmission, mutation, and recombination of IBDV between different countries and regions over the decades have led to the emergence of different genotypic strains [21,22]. Recently, as more diverse strains are discovered, some new universal genogrouping methods have been proposed [21,23,24].
Michel and Jackwood proposed a new viral classification method based on the evolutionary analysis of IBDV segment A sequences in 2017 [21]. They sequenced 90 samples from 23 countries worldwide and selected a 579 bp (nt 743–1331) fragment of segment A containing the VP2 hypervariable region for phylogenetic analysis. They classified IBDV into seven genogroups (G1–G7). And it could partially characterize the previous classification of IBDV. For example, G1 contains most classical IBDV strains, G2 contains American variants, and G3 contains vvIBDV strains. Furthermore, strains isolated from South America represent G4, while G5 and G6 include representative strains from Mexico and Italy, respectively. The Australian and Russian strains make up G7. Jackwood et al. suggested further subdivision of the different genomes into different subgroups and proposed a more specific scientific nomenclature [22]—these proposals are encouraging.
Several studies suggested that the pathogenicity of IBDV is due to both of its genomic segments and that segmental reassortment also plays an important role in viral evolution [72,96]. The genetic data of segment A alone are insufficient to characterize the potential pathogenicity of IBDV, so it is necessary to classify IBDV based on both genomic segments. Islam et al. proposed a phylogenetic analysis based on the two segments of IBDV genome and selected a 366 bp region of segment A (nt 785–1150, aa 219–340) and a 508 bp region of segment B (nt 328–835, aa 73–241), classifying the segment A of IBDV into nine genogroups and B into five (nucleotide counting starts at the 5′ terminal of the IBDV genome) [23]. The fragment selected for segment A contains a VP2 hypervariable region, and segment B contains the “B marker region” (nt 328–756, aa 110–252), which could characterize the phylogenetic evolution of segment B [97]. Thus, genogroups of segment A were classified as A0 (serotypeII), A1 (A1a: classical virulent, A1b: classical attenuated), A2 (US antigenic variant), A3 (very virulent), A4 (early European and recent South American distinct IBDV, dIBDV), A5 (atypical or recombinant Mexican strains), A6 (atypical Italian), A7 (early Australian), and A8 (Australian variant). Genogroups of segment B were classified as B1 (classical-like), B2 (very virulent-like), B3 (early Australian-like), B4 (Polish and Tanzanian), and B5 (Nigerian).
Meanwhile, Wang et al. proposed a similar scheme to classify IBDV genogroups [24], using the HVR of segment A (nt 616–1050, aa 206–350) and the B-marker of segment B (nt 328–756, aa 110–252). Similarly, IBDV is divided into nine genogroups of “A” and five of “B” in this scheme. However, the A2 of serotypeI strains is further divided into four subgroups: A2a, A2b, A2c, and A2d. The novel variant of IBDV that has recently emerged in China is classified as subgroup A2d [6]. A8 is defined as attenuated strains with specific cell tropism and non-pathogenic characteristics. B3 consisted of HLJ0504-like strains isolated in China [98], and B4 consisted mainly of recently discovered European transitional-lineage strains [99,100]. In addition, the serotypeII strains were defined as AII and BII, respectively.
All these studies have contributed greatly to the molecular epidemiology of IBDV worldwide. We have summarized their genomic classifications in Table 1 and Table 2. Despite differences in the length of genomic fragments selected for genogrouping and the philosophy behind the classifying systems, they all have similar conclusions, indicating the reliability of this system. However, there is some controversy regarding the classification of early Australian strains, Australian variants, and attenuated strains. Besides, the scheme of Wang et al. did not include the B5 (Nigerian) genogroup for analysis [101].

Table 1.
Genogroups of IBDV segment A.
Table 2.
Genogroups of IBDV segment B.
3.2. Revised Proposal for IBDV Genomic Classification
Following their principles of sequence selection [21,23,24], we revised the classification proposal by selecting a 391 bp fragment from segment A (nt 631–1021, aa 211–340) and a 528 bp fragment from segment B (nt 217–744, aa 73–248) for phylogenetic analysis of IBDV. These fragments included as much as possible the VP2 hypervariable region of segment A and the “B marker region” of segment B, as well as sequences reported in the literature and representative sequences selected by combining Islam et al. [23] and Wang et al. [24]. Sequence information for IBDV was obtained from NCBI (www.ncbi.nlm.nih.gov, accessed on 3 March 2023). The resulting phylogenetic analyses are shown in Figure 1 and Figure 2. Notably, the genomes of nVarIBDV appear as separate subgroups for both segments A and B.
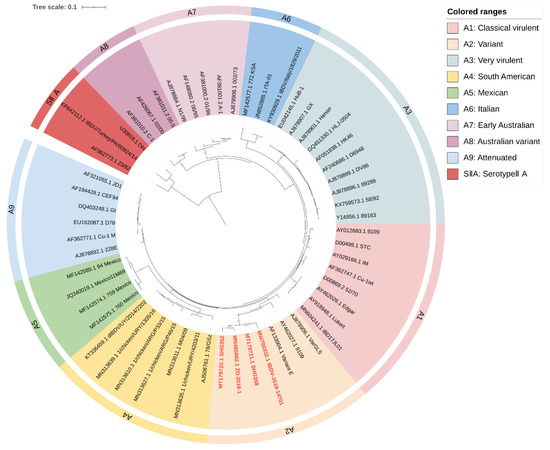
Figure 1.
Concise circular phylogenetic analysis based on the VP2 hypervariable region of IBDV segment A. The analysis of 56 IBDV strains was performed using MEGA X [102] with the maximum likelihood method, and 1000 bootstrap replications were included. The tree was annotated in iTOL (https://itol.embl.de, accessed on 3 March 2023) and drawn to scale, with genogroup information displayed in colored circular stripes on the outermost ring. The names and GenBank accession numbers of the strains are shown in the figure. The novel variant strains recently reported belonging to the A2 are shown in red bold.
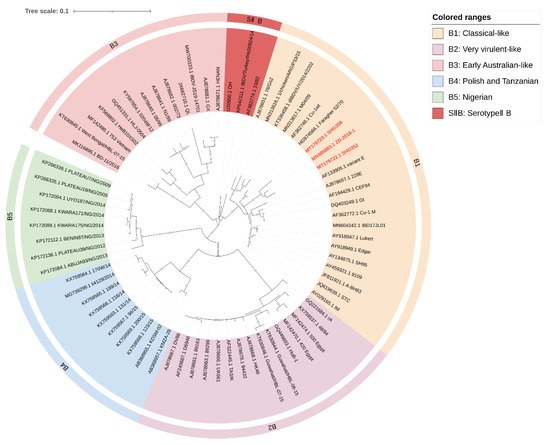
Figure 2.
Concise circular phylogenetic analysis based on the “B marker region” of IBDV segment B. The analysis of 71 IBDV strains was performed using MEGA X [102] with the maximum likelihood method, and 1000 bootstrap replications were included. The tree was annotated in iTOL (https://itol.embl.de, accessed on 3 March 2023) and drawn to scale, with genogroup information displayed in colored circular stripes on the outermost ring. The names and GenBank accession numbers of the strains are shown in the figure. The novel variant strains recently reported belonging to the B1 are shown in red bold.
Our analysis largely agrees with the classification schemes mentioned above [21,23,24], but we made some suggestions for the controversial areas. Attenuated strains are mainly derived from cIBDV or vvIBDV and could theoretically belong to the same subgroup as A1 (classical virulent) [5,26]. However, they form unique subgroups in the phylogenetic tree (Figure 1). Furthermore, the separate subgrouping of attenuated strains helps to define reassorting virulent strains. Therefore, we propose to define attenuated strains as a new subgroup called A9 (attenuated). Similar situations were observed for the early Australian and the Australian variant strains, which formed separate clades in the phylogenetic tree (Figure 1). To avoid adding further confusion to the existing genogroup classification criteria, we propose to retain the classification scheme of Islam et al. for the Australian strains as A7 (early Australian) and A8 (Australian variant). In addition, our results indicate that segment A of serotypeII has shown distinct subgroups (Figure 1), while the branches of segment B, although not forming independent subgroups (Figure 2), clearly cannot be classified as B1 (classical-like) [23]. And the existing nomenclature of either A0 [23] or AII [24] may not be clear at the level of presentation. Therefore, we propose that the genomes of serotypeII be designated as SIIA and SIIB, respectively. In addition, our phylogenetic analysis shows that the genetic lineage of segment A follows a clear stepwise pattern with distinct genogroups, where SIIA represents the most distant group (Figure 1). However, segment B exhibits more complex lineage divisions that differ from segment A (Figure 2), suggesting that the two segments may have followed different evolutionary pathways.
Overall, the phylogenetic evolution of IBDV is a complex process closely linked to genetic reorganization and the continuous emergence of new subtypes. The new genotyping scheme provides a more detailed and accurate classification of the genetic evolution of IBDV. With further improvement, it can help researchers better understand the epidemiological and molecular evolutionary mechanisms of IBDV and provide more reliable and effective theoretical support for the prevention and control of IBD.
4. Genetic Factors Affecting IBDV–Host Interactions
The interaction between IBDV and its host is influenced by several genetic factors, which have been well discussed in other reviews [34,103,104,105]. The variability of the IBDV genome is a critical factor affecting the interaction, involving mutations, gene reassortment and genetic recombination, and accounting for the emergence of multiple genotypes. The selection pressure associated with the widespread use of vaccines and the long-term transmission and prevalence of IBDV has resulted in the accumulation of multiple mutations and recombination in the IBDV genome sequence [7,14]. Such mutations help IBDV evade recognition and clearance by the immune system, thereby increasing its survival [30]. In this section, we focus on the amino acid mutations in IBDV that have been identified and partially studied, as well as the genome reassortment and genome recombination among the viruses.
4.1. Mutations in IBDV
Gene mutation is one of the major mechanisms of the genetic evolution of IBDV. Lines of evidence indicate that the VP2 hypervariable region is the most readily mutated region in the IBDV genome and has the most important impact on the antigenicity and pathogenicity of IBDV [36,56,106,107]. VP2 is the major structural protein of IBDV, serving as the primary protective antigen of the virus and inducing the production of neutralizing antibodies [108,109]. As shown in Figure 3, VP2 is folded into three structural domains, namely the base (B), the shell (S) and the projection (P) [107,110,111]. Among them, the B and S structural domains are relatively conserved, while the P domain is multivariate and contains the hypervariable region of VP2 [56,112]. Furthermore, the P domain contains four loops, namely PBC (aa 204–236), PDE (aa 240–265), PFG (aa 270–293), and PHI (aa 305–337) [56,107,110]. Previous studies have reported that the PDE and PFG domains mainly affect the cellular adaptability and pathogenicity of the virus [70,113,114]. In contrast, the PBC and PHI domains are responsible for the antigenic variability and immune escape of IBDV [30,56], which is also indicated in Table 3. Moreover, the PBC and PHI structural domains may also play a role in the process of virus assembly and maturation (Table 3).
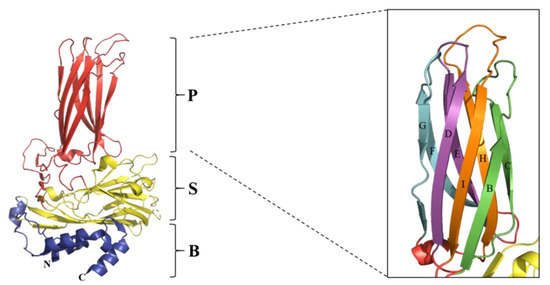
Figure 3.
Ribbon diagram of the predictive structure of IBDV VP2. The Protein Data Bank accession code is PDB-7VRP [115]. The VP2 subunit is folded into three domains: P domain (red), S domain (yellow), and B domain (blue). The right side of the figure shows the β-sheets of the P domain, with the four loops marked in different colors and the names in uppercase letters. The figure was generated using PyMOL (The PyMOL Molecular Graphics System) [116].
Earlier studies on mutation sites in the VP2 focused on antigenic variants. For example, mutations in amino acid residues 213, 222, 223, 249, and 324 have been found to be associated with loss of responsiveness to specific monoclonal antibodies [30,53,106,112]. However, in a later study, it was found that mutations in residue 222 are involved not only in immune escape but also in viral replication and virulence [56]. Other amino acid residues, such as 286 I, 318 D, and 321 A, have also been strongly associated with antigenic variants in IBDV [53,56,59]. It was found that mutations at residues 222 and 254 can cause IBDV to break through the immunity induced by the parental Del-E strain vaccine [30]. In addition, it was recently shown that mutations in residues 318 and 323 of VP2 significantly affect the antiserum neutralization of nVarIBDV with genotype A2dB1 [117]. The results from these studies suggest that amino acid mutations located in the hypervariable region of VP2 have a crucial impact on the responsiveness of the neutralizing antibodies and can lead to successful infection of chicks by the mutant strain even when maternal antibodies remain relatively high. In addition, several amino acid residues have been found to influence the cell tropism and pathogenicity of IBDV, including D279N, which has been proven to contribute to viral adaptation to cell culture and is a marker of reduced pathogenicity [113,118]. Residues at positions 253 and 284 were shown to be the determinants affecting cell tropism, with 253 H being associated with attenuation of virulence and a main contributor to IBDV virulence [69,70,113,114,119]. Mutations in residues 249 and 256 were associated with viral replication and virulence, while the additional mutation Q249R introduced on the Q253H/A284T mutation basis in VP2 can further attenuate IBDV [77]. Mutations in residue V321A have also been related to the low pathogenicity of strain 94,432 [72]. There was some controversy regarding the role of VP2 residue 279 initially, but in a later study, it was confirmed that mutations in 279 do not contribute to IBDV virulence [71].
Furthermore, the Ile-Asp-Ala (IDA) sequence (aa 234 to 236) within the VP2 P structural domain was identified to use α4β1 integrin as a possible binding receptor for invading avian cells [78], suggesting that IBDV can use the receptor-mediated pathway to enter host cells and that VP2, as a structural protein of IBDV, must play an important role in this process. A recent study deciphered the structure of IBDV virus particles using cryo-electron microscopy and found that IBDV may use two receptors to enter cells [115]. The first receptor binds to the upper region of the P structural domain, where residues 253 and 284 determine cell tropism. The second receptor interacts with the IDA motif located in a negatively charged internalization groove, which is consistent with previous findings [78]. Moreover, residues 219 Q and 324 Q were found to contribute to the interaction between adjacent VP2 trimers and play a role in virus assembly [115].
In addition to VP2, other viral proteins of IBDV also have important roles in viral infection, invasion, and replication [105]. However, these proteins are more conserved and have been less studied in regard to gene mutations. VP1 is the RdRp of IBDV and is responsible for transcription, initial translation, and replication of the viral genome [11,120]. It was found that VP1 also contributes to IBDV pathogenicity; for example, the A276T mutation in VP1 has been shown to attenuate virulence by affecting intermolecular interactions [72]. Similarly, the V4I substitution attenuates vvIBDV virulence and increases intracellular replication [79]. In addition, the amino acid triplet 145/146/147 (TDN, TEG, or NEG) in VP1 is an important virulence site affecting the activity of RdRp, and TDN is considered a conserved marker tripeptide in vvIBDV [121]. Different structural domains of VP1 have also been shown to play separate roles in viral replication and virulence, with the N-terminal domain likely playing a more prominent role, although its exact function remains unclear [122,123]. Furthermore, it was found that nVarIBDV shares an amino acid substitution A163V with vvIBDV, which may be associated with increased pathogenicity of nVarIBDV [14]. Some recent studies have focused on post-translational modifications of VP1, demonstrating that 186 R and 426 R can be methylated by protein arginine methyltransferase (PRMT), thus affecting polymerase activity [124,125]. Likewise, studies on VP3 of IBDV have revealed that residue 235 of the VP3 C-terminus (or residue 990 of the polyprotein) can affect the replication of attenuated IBDV in vitro and in vivo. Moreover, the C-terminus of VP3 is necessary for IBDV replication [126,127].
Table 3.
The roles of amino acid residues in IBDV mutation.
Table 3.
The roles of amino acid residues in IBDV mutation.
| Protein | Residues | Site | Impact | Refs. |
|---|---|---|---|---|
| VP2 | 213 D | PBC | Immune escape | [112] |
| 219 Q | PBC | Virus assembly | [115] | |
| 222 | PBC | Immune escape; Virus replication and virulence-related | [30,56,59,106] | |
| 223 | PBC | Immune escape | [106] | |
| 234–236 (IDA) | PBC | Intermolecular interactions | [78] | |
| 249 | PDE | Immune escape; virus replication and virulence-related | [53,77] | |
| 253 | PDE | Cellular adaptability; virulence-related | [69,70,114,119] | |
| 254 | PDE | Immune escape | [30] | |
| 256 | PDE | Virus replication and virulence-related | [77] | |
| D279N | PFG | Cellular adaptability | [113,118] | |
| 284 | PFG | Cellular adaptability | [69,70,113,114] | |
| 286 I | PFG | Immune escape | [112] | |
| 318 D | PHI | Immune escape | [56,59,106,117] | |
| 321 A | PHI | Immune escape; virulence-related | [56,72] | |
| 323 | PHI | Immune escape | [117] | |
| 324 | PHI | Immune escape; virus assembly | [106,115] | |
| VP1 | A276T | N/A a | Intermolecular interactions | [72] |
| V4I | N/A | Virus replication and virulence-related | [79] | |
| 145/146/147 (TDN b, TEG or NEG) | N/A | Virus replication and virulence-related | [121] | |
| A163V | N/A | virulence-related(undetermined) | [14] | |
| R186A | N/A | Polymerase activity related | [124] | |
| R426A | N/A | Virus replication and polymerase activity related | [125] | |
| VP3 | 235(C-terminal) | N/A | Virus replication | [126,127] |
a N/A—not applicable. b TDN—conserved tripeptide in the vvIBDV pathotype.
4.2. Gene Reassortment and Recombination in IBDV
In addition to mutations, gene reassortment and recombination are important genetic factors in IBDV that cannot be ignored. Genome reassortment of different segments in IBDV has been reported in various regions across the globe and the newly emerging variant strains are becoming a major threat to the poultry industry [100,128,129,130,131]. As the IBDV genome has subgroups of segments A and B, the widespread use of live attenuated vaccines has increased the occurrence of reassortment between vvIBDV and attenuated strains. These reassortant viruses may exhibit virulence comparable to that of classical or attenuated IBDV or may still inherit the high virulence of vvIBDV [76,132,133]. However, reassortant viruses with serotypeII segment A, regardless of the genotype of segment B, do not cause clinical disease in chickens or turkeys [129]. Therefore, evaluation of the potential virulence of reassortant viruses requires genotyping of the two genomic fragments and analysis of specific amino acid site changes.
Genome recombination is infrequent in the genetic evolution of IBDV. It is possibly due to the extreme evolutionary dynamics of segmented RNA viruses, which exhibit high rates of mutation and recombination but little homologous recombination [14]. In 2008, recombination events in IBDV segment A involving attenuated vaccine strains and two wild-type strains of vvIBDV and varIBDV were first described [93]. Subsequent studies have identified several very virulent strains whose major putative parents are vaccine strains but whose hypervariable regions are from vvIBDV, with recombination breakpoints mainly at 636 nt and 1743 nt [94]. Recently, homologous recombination was also found to occur between a nVarIBDV and an intermediate vaccine strain, resulting in increased pathogenicity of the nVarIBDV strain to chicken embryos, with recombination breakpoints at 1538 nt [134]. Another study has shown that a field isolate underwent both reassortment and recombination, resulting in enhanced virulence of the intermediate vaccine strain, with recombination breakpoints at 1468 nt and 1648 nt [95]. These events suggest that genetic recombination could occur naturally between different strains and plays a role in IBDV genetic diversity. Interestingly, almost all of the recombination breakpoints identified to date have occurred at either end of the VP2 hypervariable regions, which appear to have a unique propensity for recombination. The role of these regions in the genetic evolution of IBDV requires further investigation.
As the genetic factors affecting IBDV–host interactions are complex and diverse, mutations, reassortment, and recombination are among the important factors influencing the interaction. This variability of IBDV may affect its infectivity and virulence for host cells, as well as its ability for immune escape. Further studies on the genetic evolution and genotyping of IBDV are crucial for a better understanding of the pathogenesis of IBDV infection.
5. Role of Reverse Genetics in Research on IBDV Genomic Function and Vaccine Development
Reverse genetics is a powerful tool for studying the function of viral genomes. Since the successful construction and rescue of the IBDV infectious clone [135,136], significant progress has been made in understanding the relationship between the genetic variation of IBDV and its pathogenesis [137,138]. Most of the aforementioned findings have been made using reverse genetic techniques. In addition to exploring the function of the viral genome, reverse genetic techniques are a promising tool for the development of vaccines.
Several excellent reviews have covered the development of vaccines against IBDV [5,26,105]. Although various types of vaccines have been developed for IBDV, there is still a large demand for novel effective vaccines. Emerging variant strains can overcome maternal immunity induced by commercial vaccines, indicating that current vaccines are not suitable for the control of the epidemics caused by such strains [9,31,100,129,139]. The reverse genetic technique provides a new tool for vaccine development compared to traditional inactivated and attenuated vaccines. To date, reverse genetic technique has been employed to generate different rescued strains with great potential for vaccine candidates. By inserting the VP2 sequence from circulating strains into the backbone of vaccine strains, several chimeric viruses have been generated, and vaccination of chickens with the chimeric viruses could effectively protect flocks against parental strains [39,140,141]. Knocking out VP5 of IBDV produced a VP5-deficient strain, and chickens immunized with this strain were resistant to challenges with the parental virus [138,142]. Attenuated IBDV, produced by reducing the RNA polymerase activity of VP1, can induce immune protection [143]. In addition, the recombinant viruses generated by introducing the Q253H/A284T mutation into VP2 of the endemic strain and replacing it in the backbone of the attenuated strain could confer immune protection against nVarIBDV [88]. As novel mutant strains of IBDV continue to emerge, the current strategies for the prevention and control of IBD encounter new challenges. The reverse genetic techniques provide an effective approach to the development of novel IBDV vaccines that may hold great promise for the prevention and control of IBD.
6. Conclusions
Investigation into the pathogenesis and immune control of IBDV has been ongoing for decades. Although excellent progress has been made, frequent occurrences of IBD serve as a reminder that prevention and control methods for IBD need to be further explored. As studies of the IBDV genome and protein function progress, several questions need to be addressed. For example, what are the consequences of the involvement of genomic changes in the IBDV–host interaction? Can reverse genetic techniques be used to develop an optimal live vaccine that can provide full protection against IBD without causing damage to the bursa of Fabricius? In addition, what is the exact mechanism underlying amino acid mutations that alter the virulence of IBDV? Of note, as different chicken breeds have varying susceptibility to IBDV [32], anti-defense breeding approaches are also worth exploring to combat IBDV infection. Although live attenuated vaccines have been routinely used for the clinical control of IBD, the rapid generation of attenuated vaccine strains by reverse genetic techniques is definitely a promising option to combat the variant strains. However, potential threats, such as reassortment between vaccine and endemic strains and reversion of live vaccine virulence, must be carefully considered in vaccine development. Considering that the elucidation of the genomic diversity and variability of IBDV is crucial for understanding viral evolution, antigenicity, pathogenicity, and vaccine development, more efforts will be required to delve deeper into the mechanisms of IBDV–host interactions to provide necessary clues for the manipulation of IBDV by reverse genetics, which ultimately lead to the development of novel effective vaccines for prevention and control of IBD outbreaks.
Author Contributions
Conceived and designed, S.J.Z.; writing—original draft preparation, H.G.; revised the paper, S.J.Z., L.G. and Y.W. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by grants from the National Natural Science Foundation of China (# 32130105), the National Key Research and Development Program of China (2022YFD1800300) and the earmarked fund for CARS-40, China.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data available on request from the authors.
Acknowledgments
We thank all the investigators who contributed to the IBDV study and apologize to the authors of articles on relevant research that were not cited in this manuscript due to limited space.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Becht, H. Infectious Bursal Disease Virus. In Current Topics in Microbiology and Immunology; Springer: Berlin/Heidelberg, Germany, 1980; pp. 107–121. ISBN 978-3-642-67717-5. [Google Scholar]
- Berg, T.P.V.D. Acute infectious bursal disease in poultry: A review. Avian Pathol. 2000, 29, 175–194. [Google Scholar] [CrossRef] [PubMed]
- Müller, H.; Islam, M.R.; Raue, R. Research on infectious bursal disease—The past, the present and the future. Vet. Microbiol. 2003, 97, 153–165. [Google Scholar] [CrossRef]
- McFerran, J.; McNulty, M.; McKillop, E.; Connor, T.; McCracken, R.; Collins, D.; Allan, G. Isolation and serological studies with infectious bursal disease viruses from fowl, turkeys and ducks: Demonstration of a second serotype. Avian Pathol. 1980, 9, 395–404. [Google Scholar] [CrossRef]
- Jackwood, D.J. Advances in vaccine research against economically important viral diseases of food animals: Infectious bursal disease virus. Vet. Microbiol. 2017, 206, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Wu, T.; Hussain, A.; Gao, Y.; Zeng, X.; Wang, Y.; Gao, L.; Li, K.; Wang, Y.; Liu, C.; et al. Novel variant strains of infectious bursal disease virus isolated in China. Vet. Microbiol. 2019, 230, 212–220. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, X.; Gao, Y.; Qi, X. The Over-40-Years-Epidemic of Infectious Bursal Disease Virus in China. Viruses 2022, 14, 2253. [Google Scholar] [CrossRef] [PubMed]
- Zachar, T.; Popowich, S.; Goodhope, B.; Knezacek, T.; Ojkic, D.; Willson, P.; Ahmed, K.A.; Gomis, S. A 5-year study of the incidence and economic impact of variant infectious bursal disease viruses on broiler production in Saskatchewan, Canada. Can. J. Vet. Res. 2016, 80, 255–261. [Google Scholar]
- Myint, O.; Suwanruengsri, M.; Araki, K.; Izzati, U.Z.; Pornthummawat, A.; Nueangphuet, P.; Fuke, N.; Hirai, T.; Jackwood, D.J.; Yamaguchi, R. Bursa atrophy at 28 days old caused by variant infectious bursal disease virus has a negative economic impact on broiler farms in Japan. Avian Pathol. 2021, 50, 6–17. [Google Scholar] [CrossRef]
- Dey, S.; Pathak, D.; Ramamurthy, N.; Maity, H.K.; Chellappa, M.M. Infectious bursal disease virus in chickens: Prevalence, impact, and management strategies. VMRR 2019, 10, 85–97. [Google Scholar] [CrossRef]
- von Einem, U.I.; Gorbalenya, A.E.; Schirrmeier, H.; Behrens, S.-E.; Letzel, T.; Mundt, E. VP1 of infectious bursal disease virus is an RNA-dependent RNA polymerase. J. Gen. Virol. 2004, 85, 2221–2229. [Google Scholar] [CrossRef]
- Domingo, E.; Holland, J. RNA virus mutations and fitness for survival. Annu. Rev. Microbiol. 1997, 51, 151–178. [Google Scholar] [CrossRef] [PubMed]
- Islam, M.R.; Zierenberg, K.; Müller, H. The genome segment B encoding the RNA-dependent RNA polymerase protein VP1 of very virulent infectious bursal disease virus (IBDV) is phylogenetically distinct from that of all other IBDV strains. Arch. Virol. 2001, 146, 2481–2492. [Google Scholar] [CrossRef]
- Wang, W.; He, X.; Zhang, Y.; Qiao, Y.; Shi, J.; Chen, R.; Chen, J.; Xiang, Y.; Wang, Z.; Chen, G.; et al. Analysis of the global origin, evolution and transmission dynamics of the emerging novel variant IBDV (A2dB1b): The accumulation of critical aa-residue mutations and commercial trade contributes to the emergence and transmission of novel variants. Transbounding Emerg. Dis 2022, 69, e2832–e2851. [Google Scholar] [CrossRef] [PubMed]
- Wei, Y.; Li, J.; Zheng, J.; Xu, H.; Li, L.; Yu, L. Genetic reassortment of infectious bursal disease virus in nature. Biochem. Biophys. Res. Commun. 2006, 350, 277–287. [Google Scholar] [CrossRef]
- Van den Berg, T.; Morales, D.; Eterradossi, N.; Rivallan, G.; Toquin, D.; Raue, R.; Zierenberg, K.; Zhang, M.; Zhu, Y.; Wang, C. Assessment of genetic, antigenic and pathotypic criteria for the characterization of IBDV strains. Avian Pathol. 2004, 33, 470–476. [Google Scholar] [CrossRef]
- Xu, M.; Lin, S.; Zhao, Y.; Jin, J.; Tang, N.; Zhang, G. Characteristics of very virulent infectious bursal disease viruses isolated from Chinese broiler chickens (2012–2013). Acta Trop. 2015, 141, 128–134. [Google Scholar] [CrossRef]
- Xu, A.; Pei, Y.; Zhang, K.; Xue, J.; Ruan, S.; Zhang, G. Phylogenetic analyses and pathogenicity of a variant infectious bursal disease virus strain isolated in China. Virus Res. 2020, 276, 197833. [Google Scholar] [CrossRef] [PubMed]
- Tomás, G.; Marandino, A.; Courtillon, C.; Amelot, M.; Keita, A.; Pikula, A.; Hernández, M.; Hernández, D.; Vagnozzi, A.; Panzera, Y. Antigenicity, pathogenicity and immunosuppressive effect caused by a South American isolate of infectious bursal disease virus belonging to the “distinct” genetic lineage. Avian Pathol. 2019, 48, 245–254. [Google Scholar] [CrossRef] [PubMed]
- Tomás, G.; Hernández, M.; Marandino, A.; Hernández, D.; Techera, C.; Grecco, S.; Panzera, Y.; Pérez, R. Genome sequence of a distinct infectious bursal disease virus. Genome Announc. 2015, 3, e01061-15. [Google Scholar] [CrossRef]
- Michel, L.O.; Jackwood, D.J. Classification of infectious bursal disease virus into genogroups. Arch. Virol. 2017, 162, 3661–3670. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, D.J.; Schat, K.A.; Michel, L.O.; de Wit, S. A proposed nomenclature for infectious bursal disease virus isolates. Avian Pathol. 2018, 47, 576–584. [Google Scholar] [CrossRef]
- Islam, M.R.; Nooruzzaman, M.; Rahman, T.; Mumu, T.T.; Rahman, M.M.; Chowdhury, E.H.; Eterradossi, N.; Müller, H. A unified genotypic classification of infectious bursal disease virus based on both genome segments. Avian Pathol. 2021, 50, 190–206. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Fan, L.; Jiang, N.; Gao, L.; Li, K.; Gao, Y.; Liu, C.; Cui, H.; Pan, Q.; Zhang, Y.; et al. An improved scheme for infectious bursal disease virus genotype classification based on both genome-segments A and B. J. Integr. Agric. 2021, 20, 1372–1381. [Google Scholar] [CrossRef]
- Eterradossi, N.; Saif, Y. Infectious Bursal Disease (Gumboro Disease), Chapter 3.3.12. OIE Terrestrial Manual. Available online: http://www.oie.int/en/standard-setting/terrestrial-manual/access-online/ (accessed on 1 February 2023).
- Müller, H.; Mundt, E.; Eterradossi, N.; Islam, M.R. Current status of vaccines against infectious bursal disease. Avian Pathol. 2012, 41, 133–139. [Google Scholar] [CrossRef]
- Rautenschlein, S.; Kraemer, C.; Vanmarcke, J.; Montiel, E. Protective efficacy of intermediate and intermediate plus infectious bursal disease virus (IBDV) vaccines against very virulent IBDV in commercial broilers. Avian Dis. 2005, 49, 231–237. [Google Scholar] [PubMed]
- Block, H.; Meyer-Block, K.; Rebeski, D.E.; Scharr, H.; de Wit, S.; Rohn, K.; Rautenschlein, S. A field study on the significance of vaccination against infectious bursal disease virus (IBDV) at the optimal time point in broiler flocks with maternally derived IBDV antibodies. Avian Pathol. 2007, 36, 401–409. [Google Scholar] [CrossRef]
- Snyder, D.; Vakharia, V.; Savage, P. Naturally occurring-neutralizing monoclonal antibody escape variants define the epidemiology of infectious bursal disease viruses in the United States. Arch. Virol. 1992, 127, 89–101. [Google Scholar] [CrossRef]
- Jackwood, D.J.; Sommer-Wagner, S.E. Amino acids contributing to antigenic drift in the infectious bursal disease Birnavirus (IBDV). Virology 2011, 409, 33–37. [Google Scholar] [CrossRef]
- Hou, B.; Wang, C.; Luo, Z.; Shao, G. Commercial vaccines used in China do not protect against a novel infectious bursal disease virus variant isolated in Fujian. Vet. Rec. 2022, 191, e1840. [Google Scholar] [CrossRef]
- Ingrao, F.; Rauw, F.; Lambrecht, B.; van den Berg, T. Infectious Bursal Disease: A complex host–pathogen interaction. Dev. Comp. Immunol. 2013, 41, 429–438. [Google Scholar] [CrossRef]
- Trapp, J.; Rautenschlein, S. Infectious bursal disease virus’ interferences with host immune cells: What do we know? Avian Pathol. 2022, 51, 303–316. [Google Scholar] [CrossRef] [PubMed]
- Qin, Y.; Zheng, S. Infectious Bursal Disease Virus-Host Interactions: Multifunctional Viral Proteins that Perform Multiple and Differing Jobs. Int. J. Mol. Sci. 2017, 18, 161. [Google Scholar] [CrossRef]
- Li, J.; Zheng, S.J. Role of MicroRNAs in host defense against infectious bursal disease virus (IBDV) infection: A hidden front line. Viruses 2020, 12, 543. [Google Scholar] [CrossRef] [PubMed]
- Kasanga, C.; Yamaguchi, T.; Wambura, P.; Maeda-Machang’u, A.; Ohya, K.; Fukushi, H. Molecular characterization of infectious bursal disease virus (IBDV): Diversity of very virulent IBDV in Tanzania. Arch. Virol. 2007, 152, 783–790. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, D.H.; Saif, Y.M. Antigenic diversity of infectious bursal disease viruses. Avian Dis. 1987, 31, 766–770. [Google Scholar] [CrossRef] [PubMed]
- Mundt, E.; de Haas, N.; van Loon, A.A. Development of a vaccine for immunization against classical as well as variant strains of infectious bursal disease virus using reverse genetics. Vaccine 2003, 21, 4616–4624. [Google Scholar] [CrossRef]
- Gao, L.; Qi, X.; Li, K.; Gao, H.; Gao, Y.; Qin, L.; Wang, Y.; Wang, X. Development of a tailored vaccine against challenge with very virulent infectious bursal disease virus of chickens using reverse genetics. Vaccine 2011, 29, 5550–5557. [Google Scholar] [CrossRef]
- Qi, X.; Wang, Y.; Gao, L.; Gao, H.; Gao, Y.; Wang, X. Development and Application of the Reverse Genetic Technologies for Infectious Bursal Disease Virus. Bing Du Xue Bao Chin. J. Virol. 2015, 31, 326–331. [Google Scholar]
- Yang, H.; Ye, C. Reverse genetics approaches for live-attenuated vaccine development of infectious bursal disease virus. Curr. Opin. Virol. 2020, 44, 139–144. [Google Scholar] [CrossRef]
- Cosgrove, A. An apparently new disease of chickens: Avian nephrosis. Avian Dis. 1962, 6, 385–389. [Google Scholar] [CrossRef]
- Sharma, J.M.; Kim, I.J.; Rautenschlein, S.; Yeh, H.Y. Infectious bursal disease virus of chickens: Pathogenesis and immunosuppression. Dev. Comp. Immunol. 2000, 24, 223–235. [Google Scholar] [CrossRef] [PubMed]
- Vasconcelos, A.C.; Lam, K.M. Apoptosis induced by infectious bursal disease virus. J. Gen. Virol. 1994, 75, 1803–1806. [Google Scholar] [CrossRef]
- Aricibasi, M.; Jung, A.; Heller, E.D.; Rautenschlein, S. Differences in genetic background influence the induction of innate and acquired immune responses in chickens depending on the virulence of the infecting infectious bursal disease virus (IBDV) strain. Vet. Immunol. Immunopathol. 2010, 135, 79–92. [Google Scholar] [CrossRef] [PubMed]
- Müller, H.; Scholtissek, C.; Becht, H. The genome of infectious bursal disease virus consists of two segments of double-stranded RNA. J. Virol. 1979, 31, 584–589. [Google Scholar] [CrossRef] [PubMed]
- Mundt, E.; Müller, H. Complete Nucleotide Sequences of 5′- and 3′-Noncoding Regions of Both Genome Segments of Different Strains of Infectious Bursal Disease Virus. Virology 1995, 209, 10–18. [Google Scholar] [CrossRef]
- Chen, F.; Liu, J.; Yan, Z.; Liu, D.; Ji, J.; Qin, J.; Li, H.; Ma, J.; Bi, Y.; Xie, Q. Complete genome sequence analysis of a natural reassortant infectious bursal disease virus in China. J. Virol. 2012, 86, 11942. [Google Scholar] [CrossRef]
- Raja, P.; Senthilkumar, T.; Parthiban, M.; Thangavelu, A.; Gowri, A.M.; Palanisammi, A.; Kumanan, K. Complete genome sequence analysis of a naturally reassorted infectious bursal disease virus from India. Genome Announc. 2016, 4, e00709-16. [Google Scholar] [CrossRef]
- Banda, A.; Villegas, P. Genetic characterization of very virulent infectious bursal disease viruses from Latin America. Avian Dis. 2004, 48, 540–549. [Google Scholar] [CrossRef]
- Hernández, M.; Tomás, G.; Marandino, A.; Iraola, G.; Maya, L.; Mattion, N.; Hernández, D.; Villegas, P.; Banda, A.; Panzera, Y. Genetic characterization of South American infectious bursal disease virus reveals the existence of a distinct worldwide-spread genetic lineage. Avian Pathol. 2015, 44, 212–221. [Google Scholar] [CrossRef]
- Rosenberger, J.; Cloud, S. Isolation and characterization of variant infectious bursal disease viruses. J. Am. Vet. Med. Assoc. 1986, 189, 357. [Google Scholar]
- Vakharia, V.N.; He, J.; Ahamed, B.; Snyder, D.B. Molecular basis of antigenic variation in infectious bursal disease virus. Virus Res. 1994, 31, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Eterradossi, N.; Toquin, D.; Rivallan, G.; Guittet, M. Modified activity of a VP2-located neutralizing epitope on various vaccine, pathogenic and hypervirulent strains of infectious bursal disease virus. Arch. Virol. 1997, 142, 255–270. [Google Scholar] [CrossRef] [PubMed]
- Bayliss, C.D.; Spies, U.; Shaw, K.; Peters, R.W.; Papageorgiou, A.; Muller, H.; Boursnell, M.E.G. A comparison of the sequences of segment A of four infectious bursal disease virus strains and identification of a variable region in VP2. J. Gen. Virol. 1990, 71, 1303–1312. [Google Scholar] [CrossRef] [PubMed]
- Letzel, T.; Coulibaly, F.; Rey, F.A.; Delmas, B.; Jagt, E.; van Loon, A.A.M.W.; Mundt, E. Molecular and Structural Bases for the Antigenicity of VP2 of Infectious Bursal Disease Virus. J. Virol. 2007, 81, 12827–12835. [Google Scholar] [CrossRef]
- Van der Marel, P.; Snyder, D.; Lütticken, D. Antigenic characterization of IBDV field isolates by their reactivity with a panel of monoclonal antibodies. DTW. Dtsch. Tierarztl. Wochenschr. 1990, 97, 81–83. [Google Scholar]
- Jackwood, D.J. Molecular epidemiologic evidence of homologous recombination in infectious bursal disease viruses. Avian Dis. 2012, 56, 574–577. [Google Scholar] [CrossRef]
- Jackwood, D.; Cookson, K.; Sommer-Wagner, S.; Galludec, H.L.; De Wit, J. Molecular characteristics of infectious bursal disease viruses from asymptomatic broiler flocks in Europe. Avian Dis. 2006, 50, 532–536. [Google Scholar] [CrossRef]
- Chettle, N.; Stuart, J.; Wyeth, P. Outbreak of virulent infectious bursal disease in East Anglia. Vet. Rec. 1989, 125, 271. [Google Scholar] [CrossRef]
- Van den Berg, T.; Gonze, M.; Meulemans, G. Acute infectious bursal disease in poultry: Isolation and characterisation of a highly virulent strain. Avian Pathol. 1991, 20, 133–143. [Google Scholar] [CrossRef]
- Di Fabio, J.; Rossini, L.; Eterradossi, N.; Toquin, M.; Gardin, Y. European-like pathogenic infectious bursal disease viruses in Brazil. Vet. Rec. 1999, 145, 203–204. [Google Scholar]
- Mawgod, S.A.; Arafa, A.S.; Hussein, H.A. Molecular genotyping of the infectious bursal disease virus (IBDV) isolated from broiler flocks in Egypt. Int. J. Vet. Sci. Med. 2014, 2, 46–52. [Google Scholar] [CrossRef]
- Barathidasan, R.; Singh, S.; Kumar, M.A.; Desingu, P.; Palanivelu, M.; Singh, M.; Dhama, K. Recurrent outbreaks of infectious bursal disease (IBD) in a layer farm caused by very virulent IBD virus (vvIBDV) in India: Pathology and molecular analysis. South Asian J. Exp. Biol. 2013, 3, 200–206. [Google Scholar] [CrossRef]
- Hernández, M.; Banda, A.; Hernández, D.; Panzera, F.; Pérez, R. Detection of very virulent strains of infectious bursal disease virus (vvIBDV) in commercial broilers from Uruguay. Avian Dis. 2006, 50, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Hon, C.-C.; Lam, T.-Y.; Drummond, A.; Rambaut, A.; Lee, Y.-F.; Yip, C.-W.; Zeng, F.; Lam, P.-Y.; Ng, P.T.W.; Leung, F.C.C. Phylogenetic Analysis Reveals a Correlation between the Expansion of Very Virulent Infectious Bursal Disease Virus and Reassortment of Its Genome Segment B. J. Virol. 2006, 80, 8503–8509. [Google Scholar] [CrossRef]
- Brown, M.D.; Skinner, M.A. Coding sequences of both genome segments of a European ‘very virulent’infectious bursal disease virus. Virus Res. 1996, 40, 1–15. [Google Scholar] [CrossRef]
- Eterradossi, N.; Arnauld, C.; Tekaia, F.; Toquin, D.; Le Coq, H.; Rivallan, G.; Guittet, M.; Domenech, J.; Van den Berg, T.; Skinner, M. Antigenic and genetic relationships between European very virulent infectious bursal disease viruses and an early West African isolate. Avian Pathol. 1999, 28, 36–46. [Google Scholar] [CrossRef]
- Mundt, E. Tissue culture infectivity of different strains of infectious bursal disease virus is determined by distinct amino acids in VP2. J. Gen. Virol. 1999, 80, 2067–2076. [Google Scholar] [CrossRef]
- Qi, X.; Gao, H.; Gao, Y.; Qin, L.; Wang, Y.; Gao, L.; Wang, X. Naturally occurring mutations at residues 253 and 284 in VP2 contribute to the cell tropism and virulence of very virulent infectious bursal disease virus. Antivir. Res. 2009, 84, 225–233. [Google Scholar] [CrossRef]
- Qi, X.; Lu, Z.; Wang, N.; Chen, Y.; Zhang, L.; Gao, L.; Li, K.; Ren, X.; Wang, Y.; Gao, H.; et al. Analysis of the function of D279N mutation of VP2 of infectious bursal disease virus. J. Integr. Agric. 2015, 14, 2618–2625. [Google Scholar] [CrossRef]
- Escaffre, O.; Nouën, C.L.; Amelot, M.; Ambroggio, X.; Ogden, K.M.; Guionie, O.; Toquin, D.; Müller, H.; Islam, M.R.; Eterradossi, N. Both Genome Segments Contribute to the Pathogenicity of Very Virulent Infectious Bursal Disease Virus. J. Virol. 2013, 87, 14. [Google Scholar] [CrossRef]
- Boot, H.J.; ter Huurne, A.A.H.M.; Hoekman, A.J.W.; Peeters, B.P.H.; Gielkens, A.L.J. Rescue of Very Virulent and Mosaic Infectious Bursal Disease Virus from Cloned cDNA: VP2 Is Not the Sole Determinant of the Very Virulent Phenotype. J. Virol. 2000, 74, 6701–6711. [Google Scholar] [CrossRef] [PubMed]
- Boot, H.J.; Hoekman, A.J.W.; Gielkens, A.L.J. The enhanced virulence of very virulent infectious bursal disease virus is partly determined by its B-segment. Arch. Virol. 2005, 150, 137–144. [Google Scholar] [CrossRef]
- Jackwood, D.J.; Sommer-Wagner, S.E.; Crossley, B.M.; Stoute, S.T.; Woolcock, P.R.; Charlton, B.R. Identification and pathogenicity of a natural reassortant between a very virulent serotype 1 infectious bursal disease virus (IBDV) and a serotype 2 IBDV. Virology 2011, 420, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Nouën, C.L.; Rivallan, G.; Toquin, D.; Darlu, P.; Morin, Y.; Beven, V.; de Boisseson, C.; Cazaban, C.; Comte, S.; Gardin, Y.; et al. Very virulent infectious bursal disease virus: Reduced pathogenicity in a rare natural segment-B-reassorted isolate. J. Gen. Virol. 2006, 87, 209–216. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Zhang, L.; Chen, Y.; Gao, L.; Wu, G.; Qin, L.; Wang, Y.; Ren, X.; Gao, Y.; Gao, H.; et al. Mutations of Residues 249 and 256 in VP2 Are Involved in the Replication and Virulence of Infectious Bursal Disease Virus. PLoS ONE 2013, 8, e70982. [Google Scholar] [CrossRef]
- Delgui, L.; Oña, A.; Gutiérrez, S.; Luque, D.; Navarro, A.; Castón, J.R.; Rodríguez, J.F. The capsid protein of infectious bursal disease virus contains a functional α4β1 integrin ligand motif. Virology 2009, 386, 360–372. [Google Scholar] [CrossRef]
- Yu, F.; Ren, X.; Wang, Y.; Qi, X.; Song, J.; Gao, Y.; Qin, L.; Gao, H.; Wang, X. A single amino acid V4I substitution in VP1 attenuates virulence of very virulent infectious bursal disease virus (vvIBDV) in SPF chickens and increases replication in CEF cells. Virology 2013, 440, 204–209. [Google Scholar] [CrossRef]
- Thai, T.N.; Jang, I.; Kim, H.-A.; Kim, H.-S.; Kwon, Y.-K.; Kim, H.-R. Characterization of antigenic variant infectious bursal disease virus strains identified in South Korea. Avian Pathol. 2021, 50, 174–181. [Google Scholar] [CrossRef]
- Wang, Y.; Jiang, N.; Fan, L.; Niu, X.; Zhang, W.; Huang, M.; Gao, L.; Li, K.; Gao, Y.; Liu, C. Identification and pathogenicity evaluation of a novel reassortant Infectious Bursal Disease Virus (genotype A2dB3). Viruses 2021, 13, 1682. [Google Scholar] [CrossRef]
- Wang, W.; Huang, Y.; Zhang, Y.; Qiao, Y.; Deng, Q.; Chen, R.; Chen, J.; Huang, T.; Wei, T.; Mo, M. The emerging naturally reassortant strain of IBDV (genotype A2dB3) having segment A from Chinese novel variant strain and segment B from HLJ 0504-like very virulent strain showed enhanced pathogenicity to three-yellow chickens. Transbound. Emerg. Dis. 2022, 69, e566–e579. [Google Scholar] [CrossRef]
- Jiang, N.; Wang, Y.; Zhang, W.; Niu, X.; Huang, M.; Gao, Y.; Liu, A.; Gao, L.; Li, K.; Pan, Q. Genotyping and molecular characterization of infectious bursal disease virus identified in important poultry-raising areas of China during 2019 and 2020. Front. Vet. Sci. 2021, 8, 759861. [Google Scholar] [CrossRef]
- Thai, T.N.; Yoo, D.-S.; Jang, I.; Kwon, Y.-K.; Kim, H.-R. Dynamics of the Emerging Genogroup of Infectious Bursal Disease Virus Infection in Broiler Farms in South Korea: A Nationwide Study. Viruses 2022, 14, 1604. [Google Scholar] [CrossRef] [PubMed]
- Aliyu, H.B.; Hair-Bejo, M.; Omar, A.R.; Ideris, A. Genetic diversity of recent infectious bursal disease viruses isolated from vaccinated poultry flocks in Malaysia. Front. Vet. Sci. 2021, 8, 643976. [Google Scholar] [CrossRef]
- Fan, L.; Wu, T.; Wang, Y.; Hussain, A.; Jiang, N.; Gao, L.; Li, K.; Gao, Y.; Liu, C.; Cui, H.; et al. Novel variants of infectious bursal disease virus can severely damage the bursa of fabricius of immunized chickens. Vet. Microbiol. 2020, 240, 108507. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Kuang, H.; Guo, H.; Cai, L.; Chu, D.; Wang, X.; Hu, J.; Rong, J. Development of a recombinant VP2 vaccine for the prevention of novel variant strains of infectious bursal disease virus. Avian Pathol. 2020, 49, 557–571. [Google Scholar] [CrossRef] [PubMed]
- Fan, L.; Wang, Y.; Jiang, N.; Gao, L.; Li, K.; Gao, Y.; Cui, H.; Pan, Q.; Liu, C.; Zhang, Y.; et al. A reassortment vaccine candidate of the novel variant infectious bursal disease virus. Vet. Microbiol. 2020, 251, 108905. [Google Scholar] [CrossRef] [PubMed]
- Lian, J.; Wang, Z.; Xu, Z.; Pang, Y.; Leng, M.; Tang, S.; Zhang, X.; Qin, J.; Chen, F.; Lin, W. Pathogenicity and molecular characterization of infectious bursal disease virus in China. Poult. Sci. 2022, 101, 101502. [Google Scholar] [CrossRef]
- Sun, J.; Lu, P.; Yan, Y.; Hua, X.; Jiang, J.; Zhao, Y. Sequence and analysis of genomic segment A and B of very virulent infectious bursal disease virus isolated from China. J. Vet. Med. Ser. B 2003, 50, 148–154. [Google Scholar] [CrossRef]
- Pikuła, A.; Lisowska, A.; Jasik, A.; Śmietanka, K. Identification and assessment of virulence of a natural reassortant of infectious bursal disease virus. Vet. Res. 2018, 49, 89. [Google Scholar] [CrossRef]
- Cui, P.; Ma, S.-J.; Zhang, Y.-G.; Li, X.-S.; Gao, X.-Y.; Cui, B.-A.; Chen, H.-Y. Genomic sequence analysis of a new reassortant infectious bursal disease virus from commercial broiler flocks in Central China. Arch. Virol. 2013, 158, 1973–1978. [Google Scholar] [CrossRef]
- Hon, C.-C.; Lam, T.T.-Y.; Yip, C.-W.; Wong, R.T.-Y.; Shi, M.; Jiang, J.; Zeng, F.; Leung, F.C.-C. Phylogenetic evidence for homologous recombination within the family Birnaviridae. J. Gen. Virol. 2008, 89, 3156–3164. [Google Scholar] [CrossRef] [PubMed]
- He, C.-Q.; Ma, L.-Y.; Wang, D.; Li, G.-R.; Ding, N.-Z. Homologous recombination is apparent in infectious bursal disease virus. Virology 2009, 384, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Zhu, N.; Cui, Y.; Hou, L.; Zhou, J.; Qiu, Y.; Yang, X.; Liu, C.; Wang, D.; Guo, J.; et al. Characterization and pathogenicity of a naturally reassortant and recombinant infectious bursal disease virus in China. Transbounding Emerg. Dis. 2022, 69, e746–e758. [Google Scholar] [CrossRef]
- He, X.; Xiong, Z.; Yang, L.; Guan, D.; Yang, X.; Wei, P. Molecular epidemiology studies on partial sequences of both genome segments reveal that reassortant infectious bursal disease viruses were dominantly prevalent in southern China during 2000–2012. Arch. Virol. 2014, 159, 3279–3292. [Google Scholar] [CrossRef]
- Alfonso-Morales, A.; Rios, L.; Martínez-Pérez, O.; Dolz, R.; Valle, R.; Perera, C.L.; Bertran, K.; Frías, M.T.; Ganges, L. Evaluation of a Phylogenetic Marker Based on Genomic Segment B of Infectious Bursal Disease Virus: Facilitating a Feasible Incorporation of this Segment to the Molecular Epidemiology Studies for this Viral Agent. PLoS ONE 2015, 10, e0125853. [Google Scholar] [CrossRef] [PubMed]
- Qi, X.; Gao, L.; Qin, L.; Deng, X.; Wu, G.; Zhang, L.; Yu, F.; Ren, X.; Gao, Y.; Gao, H. Genomic sequencing and molecular characteristics of a very virulent strain of infectious bursal disease virus isolated in China. Agric. Sci. Technol.-Hunan 2011, 12, 1946–1949. [Google Scholar]
- Tammiranta, N.; Ek-Kommonen, C.; Rossow, L.; Huovilainen, A. Circulation of very virulent avian infectious bursal disease virus in Finland. Avian Pathol. 2018, 47, 520–525. [Google Scholar] [CrossRef]
- Pikuła, A.; Śmietanka, K.; Perez, L.J. Emergence and expansion of novel pathogenic reassortant strains of infectious bursal disease virus causing acute outbreaks of the disease in Europe. Transbound. Emerg. Dis. 2020, 67, 1739–1744. [Google Scholar] [CrossRef]
- Nwagbo, I.O.; Shittu, I.; Nwosuh, C.I.; Ezeifeka, G.O.; Odibo, F.J.; Michel, L.O.; Jackwood, D.J. Molecular characterization of field infectious bursal disease virus isolates from Nigeria. Vet. World 2016, 9, 1420. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Zhang, S.; Zheng, S. Host Combats IBDV Infection at Both Protein and RNA Levels. Viruses 2022, 14, 2309. [Google Scholar] [CrossRef]
- Wang, H.; Li, W.; Zheng, S.J. Advances on Innate Immune Evasion by Avian Immunosuppressive Viruses. Front. Immunol. 2022, 13, 901913. [Google Scholar] [CrossRef]
- Zheng, S.J. Infectious Bursal Disease Virus, Chapter 7. In Avian Virology: Current Research and Future Trends; Caister Academic Press: Poole, UK, 2019; ISBN 978-1-912530-10-6. [Google Scholar]
- Eterradossi, N.; Arnauld, C.; Toquin, D.; Rivallan, G. Critical amino acid changes in VP2 variable domain are associated with typical and atypical antigenicity in very virulent infectious bursal disease viruses. Arch. Virol. 1998, 143, 1627–1636. [Google Scholar] [CrossRef] [PubMed]
- Coulibaly, F.; Chevalier, C.; Gutsche, I.; Pous, J.; Navaza, J.; Bressanelli, S.; Delmas, B.; Rey, F.A. The Birnavirus Crystal Structure Reveals Structural Relationships among Icosahedral Viruses. Cell 2005, 120, 761–772. [Google Scholar] [CrossRef] [PubMed]
- Becht, H.; Müller, H.; Müller, H.K. Comparative studies on structural and antigenic properties of two serotypes of infectious bursal disease virus. J. Gen. Virol. 1988, 69, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Eterradossi, N.; Saif, Y.M. Infectious Bursal Disease. In Diseases of Poultry; Wiley: Hoboken, NJ, USA, 2020; pp. 257–283. ISBN 978-1-119-37116-8. [Google Scholar]
- Lee, C.-C.; Ko, T.-P.; Chou, C.-C.; Yoshimura, M.; Doong, S.-R.; Wang, M.-Y.; Wang, A.H.-J. Crystal structure of infectious bursal disease virus VP2 subviral particle at 2.6Å resolution: Implications in virion assembly and immunogenicity. J. Struct. Biol. 2006, 155, 74–86. [Google Scholar] [CrossRef]
- Garriga, D.; Querol-Audí, J.; Abaitua, F.; Saugar, I.; Pous, J.; Verdaguer, N.; Castón, J.R.; Rodriguez, J.F. The 2.6-Angstrom Structure of Infectious Bursal Disease Virus-Derived T=1 Particles Reveals New Stabilizing Elements of the Virus Capsid. J. Virol. 2006, 80, 6895–6905. [Google Scholar] [CrossRef]
- Heine, H.-G.; Haritou, M.; Failla, P.; Fahey, K.; Azad, A. Sequence Analysis and Expression of the Host-protective Immunogen VP2 of a Variant Strain of Infectious Bursal Disease Virus Which Can Circumvent Vaccination with Standard Type I Strains. J. Gen. Virol. 1991, 72, 1835–1843. [Google Scholar] [CrossRef]
- Lim, B.-L.; Cao, Y.; Yu, T.; Mo, C.-W. Adaptation of very virulent infectious bursal disease virus to chicken embryonic fibroblasts by site-directed mutagenesis of residues 279 and 284 of viral coat protein VP2. J. Virol. 1999, 73, 2854–2862. [Google Scholar] [CrossRef]
- Brandt, M.; Yao, K.; Liu, M.; Heckert, R.A.; Vakharia, V.N. Molecular determinants of virulence, cell tropism, and pathogenic phenotype of infectious bursal disease virus. J. Virol. 2001, 75, 11974–11982. [Google Scholar] [CrossRef]
- Bao, K.; Qi, X.; Li, Y.; Gong, M.; Wang, X.; Zhu, P. Cryo-EM structures of infectious bursal disease viruses with different virulences provide insights into their assembly and invasion. Sci. Bull. 2022, 67, 646–654. [Google Scholar] [CrossRef] [PubMed]
- Schrödinger, L.L.C. The PyMOL Molecular Graphics System, Version 1.8. 2015. Available online: https://www.pymol.org (accessed on 3 March 2023).
- Fan, L.; Wang, Y.; Jiang, N.; Gao, Y.; Niu, X.; Zhang, W.; Huang, M.; Bao, K.; Liu, A.; Wang, S.; et al. Residues 318 and 323 in capsid protein are involved in immune circumvention of the atypical epizootic infection of infectious bursal disease virus. Front. Microbiol. 2022, 13, 909252. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Ogawa, M.; Inoshima, Y.; Miyoshi, M.; Fukushi, H.; Hirai, K. Identification of Sequence Changes Responsible for the Attenuation of Highly Virulent Infectious Bursal Disease Virus. Virology 1996, 223, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, D.J.; Sreedevi, B.; LeFever, L.J.; Sommer-Wagner, S.E. Studies on naturally occurring infectious bursal disease viruses suggest that a single amino acid substitution at position 253 in VP2 increases pathogenicity. Virology 2008, 377, 110–116. [Google Scholar] [CrossRef]
- Lombardo, E.; Maraver, A.; Castón, J.R.; Rivera, J.; Fernández-Arias, A.; Serrano, A.; Carrascosa, J.L.; Rodriguez, J.F. VP1, the putative RNA-dependent RNA polymerase of infectious bursal disease virus, forms complexes with the capsid protein VP3, leading to efficient encapsidation into virus-like particles. J. Virol. 1999, 73, 6973–6983. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Li, K.; Qi, X.; Gao, H.; Gao, Y.; Qin, L.; Wang, Y.; Shen, N.; Kong, X.; Wang, X. Triplet amino acids located at positions 145/146/147 of the RNA polymerase of very virulent infectious bursal disease virus contribute to viral virulence. J. Gen. Virol. 2014, 95, 888–897. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Li, K.; Qi, X.; Gao, Y.; Wang, Y.; Gao, H.; Wang, X. N-terminal domain of the RNA polymerase of very virulent infectious bursal disease virus contributes to viral replication and virulence. Sci. China Life Sci. 2018, 61, 1127–1129. [Google Scholar] [CrossRef]
- Wang, W.; Huang, Y.; Ji, Z.; Chen, G.; Zhang, Y.; Qiao, Y.; Shi, M.; Li, M.; Huang, T.; Wei, T.; et al. The Full Region of N-Terminal in Polymerase of IBDV Plays an Important Role in Viral Replication and Pathogenicity: Either Partial Region or Single Amino Acid V4I Substitution Does Not Completely Lead to the Virus Attenuation to Three-Yellow Chickens. Viruses 2021, 13, 107. [Google Scholar] [CrossRef]
- Hu, X.; Chen, Z.; Wu, X.; Ding, Z.; Zeng, Q.; Wu, H. An Improved, Dual-Direction, Promoter-Driven, Reverse Genetics System for the Infectious Bursal Disease Virus (IBDV). Viruses 2022, 14, 1396. [Google Scholar] [CrossRef]
- Hu, X.; Chen, Z.; Wu, X.; Fu, Q.; Chen, Z.; Huang, Y.; Wu, H. PRMT5 Facilitates Infectious Bursal Disease Virus Replication through Arginine Methylation of VP1. J. Virol. 2023, 97, e01637-22. [Google Scholar] [CrossRef]
- Wang, Y.; Qi, X.; Kang, Z.; Yu, F.; Qin, L.; Gao, H.; Gao, Y.; Wang, X. A single amino acid in the C-terminus of VP3 protein influences the replication of attenuated infectious bursal disease virus in vitro and in vivo. Antivir. Res. 2010, 87, 223–229. [Google Scholar] [CrossRef] [PubMed]
- Mosley, Y.-Y.C.; Wu, C.C.; Lin, T.L. A free VP3 C-terminus is essential for the replication of infectious bursal disease virus. Virus Res. 2017, 232, 77–79. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.; Hu, H.; Chen, G.; Liu, H.; Wang, S.; Xia, D.; Yu, Y.; Zhang, Y.; Jiang, J.; Ma, J.; et al. Identification and assessment of pathogenicity of a naturally reassorted infectious bursal disease virus from Henan, China. Poult. Sci. 2019, 98, 6433–6444. [Google Scholar] [CrossRef] [PubMed]
- Jackwood, D.J.; Stoute, S.T.; Beate, M. Crossley Pathogenicity of Genome Reassortant Infectious Bursal Disease Viruses in Chickens and Turkeys. Avian Dis. 2016, 60, 765–772. [Google Scholar] [CrossRef] [PubMed]
- Legnardi, M.; Franzo, G.; Tucciarone, C.M.; Koutoulis, K.; Duarte, I.; Silva, M.; Le Tallec, B.; Cecchinato, M. Detection and molecular characterization of a new genotype of infectious bursal disease virus in Portugal. Avian Pathol. 2022, 51, 97–105. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Wang, W.; Chen, G.; Jiao, P.; Ji, Z.; Yang, L.; Wei, P. Serological study reveal different antigenic IBDV strains prevalent in southern China during the years 2000–2017 and also the antigenic differences between the field strains and the commonly used vaccine strains. Vet. Microbiol. 2019, 239, 108458. [Google Scholar] [CrossRef]
- Wei, Y.; Yu, X.; Zheng, J.; Chu, W.; Xu, H.; Yu, X.; Yu, L. Reassortant infectious bursal disease virus isolated in China. Virus Res. 2008, 131, 279–282. [Google Scholar] [CrossRef]
- Lee, H.-J.; Jang, I.; Shin, S.-H.; Lee, H.-S.; Choi, K.-S. Genome Sequence of a Novel Reassortant and Very Virulent Strain of Infectious Bursal Disease Virus. Genome Announc. 2017, 5, e00730-17. [Google Scholar] [CrossRef]
- Wu, T.; Wang, Y.; Li, H.; Fan, L.; Jiang, N.; Gao, L.; Li, K.; Gao, Y.; Liu, C.; Cui, H.; et al. Naturally occurring homologous recombination between novel variant infectious bursal disease virus and intermediate vaccine strain. Vet. Microbiol. 2020, 245, 108700. [Google Scholar] [CrossRef]
- Mundt, E.; Vakharia, V.N. Synthetic transcripts of double-stranded Birnavirus genome are infectious. Proc. Natl. Acad. Sci. USA 1996, 93, 11131–11136. [Google Scholar] [CrossRef]
- Qi, X.; Gao, Y.; Gao, H.; Deng, X.; Bu, Z.; Wang, X.; Fu, C.; Wang, X. An improved method for infectious bursal disease virus rescue using RNA polymerase II system. J. Virol. Methods 2007, 142, 81–88. [Google Scholar] [CrossRef]
- Durairaj, V.; Sellers, H.S.; Linnemann, E.G.; Icard, A.H.; Mundt, E. Investigation of the antigenic evolution of field isolates using the reverse genetics system of infectious bursal disease virus (IBDV). Arch Virol. 2011, 156, 1717–1728. [Google Scholar] [CrossRef] [PubMed]
- Yao, K.; Goodwin, M.A.; Vakharia, V.N. Generation of a Mutant Infectious Bursal Disease Virus That Does Not Cause Bursal Lesions. J. Virol. 1998, 72, 2647–2654. [Google Scholar] [CrossRef] [PubMed]
- Kurukulsuriya, S.; Ahmed, K.A.; Ojkic, D.; Gunawardana, T.; Gupta, A.; Goonewardene, K.; Karunaratne, R.; Popowich, S.; Willson, P.; Tikoo, S.K.; et al. Circulating strains of variant infectious bursal disease virus may pose a challenge for antibiotic-free chicken farming in Canada. Res. Vet. Sci. 2016, 108, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Elankumaran, S.; Yunus, A.S.; Samal, S.K. A recombinant Newcastle disease virus (NDV) expressing VP2 protein of infectious bursal disease virus (IBDV) protects against NDV and IBDV. J. Virol. 2004, 78, 10054–10063. [Google Scholar] [CrossRef]
- Shah, A.U.; Wang, Z.; Zheng, Y.; Guo, R.; Chen, S.; Xu, M.; Zhang, C.; Liu, Y.; Wang, J. Construction of a Novel Infectious Clone of Recombinant Herpesvirus of Turkey Fc-126 Expressing VP2 of IBDV. Vaccines 2022, 10, 1391. [Google Scholar] [CrossRef]
- Qin, L.; Qi, X.; Gao, Y.; Gao, H.; Lu, X.; Wang, Y.; Bu, Z.; Wang, X. VP5-deficient mutant virus induced protection against challenge with very virulent infectious bursal disease virus of chickens. Vaccine 2010, 28, 3735–3740. [Google Scholar] [CrossRef]
- Yang, H.; Wang, Y.; Ye, C. Rapid Generation of Attenuated Infectious Bursal Disease Virus from Dual-Promoter Plasmids by Reduction of Viral Ribonucleoprotein Activity. J. Virol. 2020, 94, e01569-19. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).