
Limbic System Response to Psilocybin and Ketamine Administration in Rats: A Neurochemical and Behavioral Study

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. The Effect of Psilocybin and Ketamine on Extracellular Levels of DA and 5-HT in the Rat Nucleus Accumbens
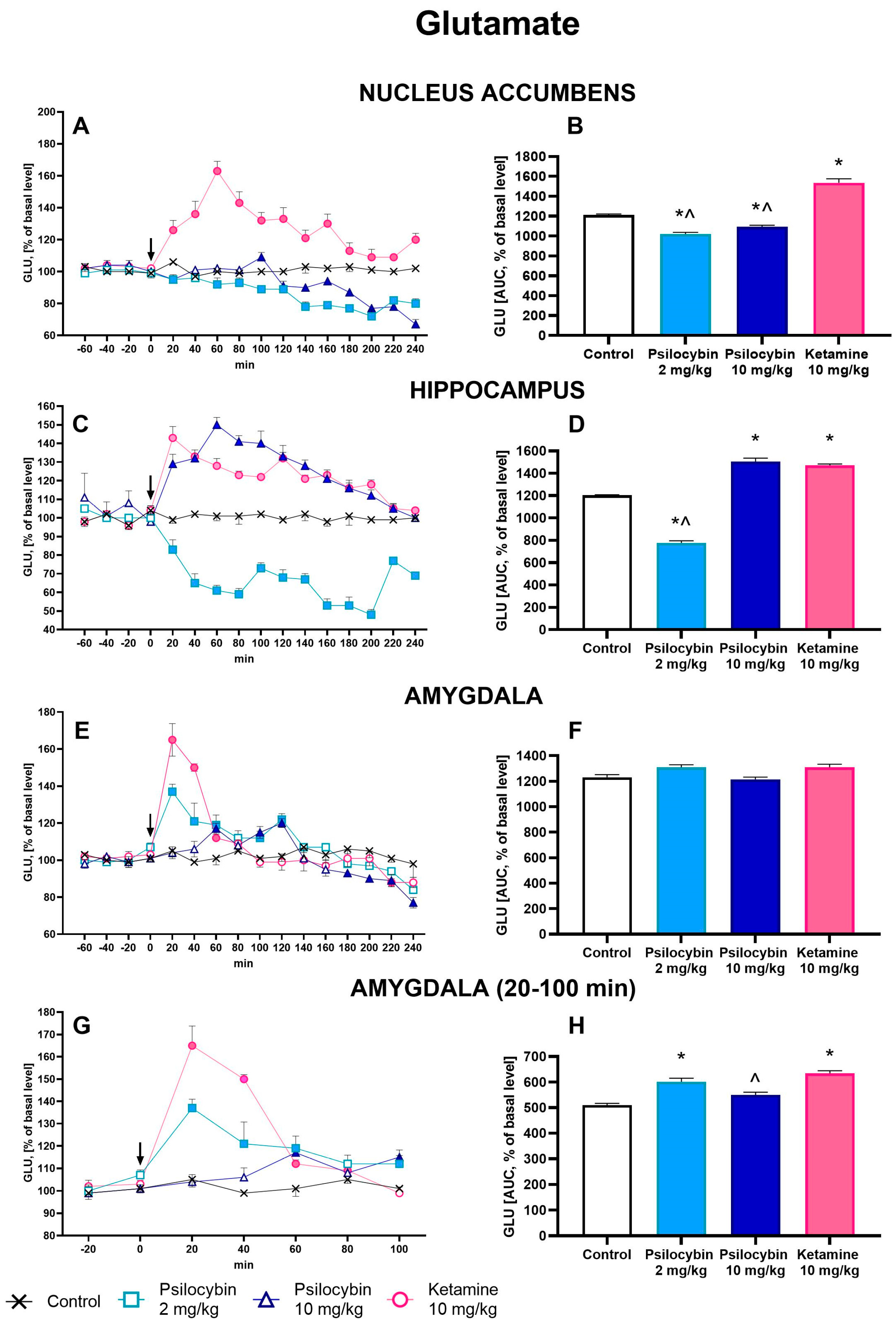
2.2. The Effect of Psilocybin and Ketamine on Extracellular Levels of Glutamate and GABA in the Rat Nucleus Accumbens, Hippocampus and Amygdala
2.3. The Effect of Psilocybin and Ketamine on GABA/Glutamate Ratio in the Rat Nucleus Accumbens, Hippocampus and Amygdala
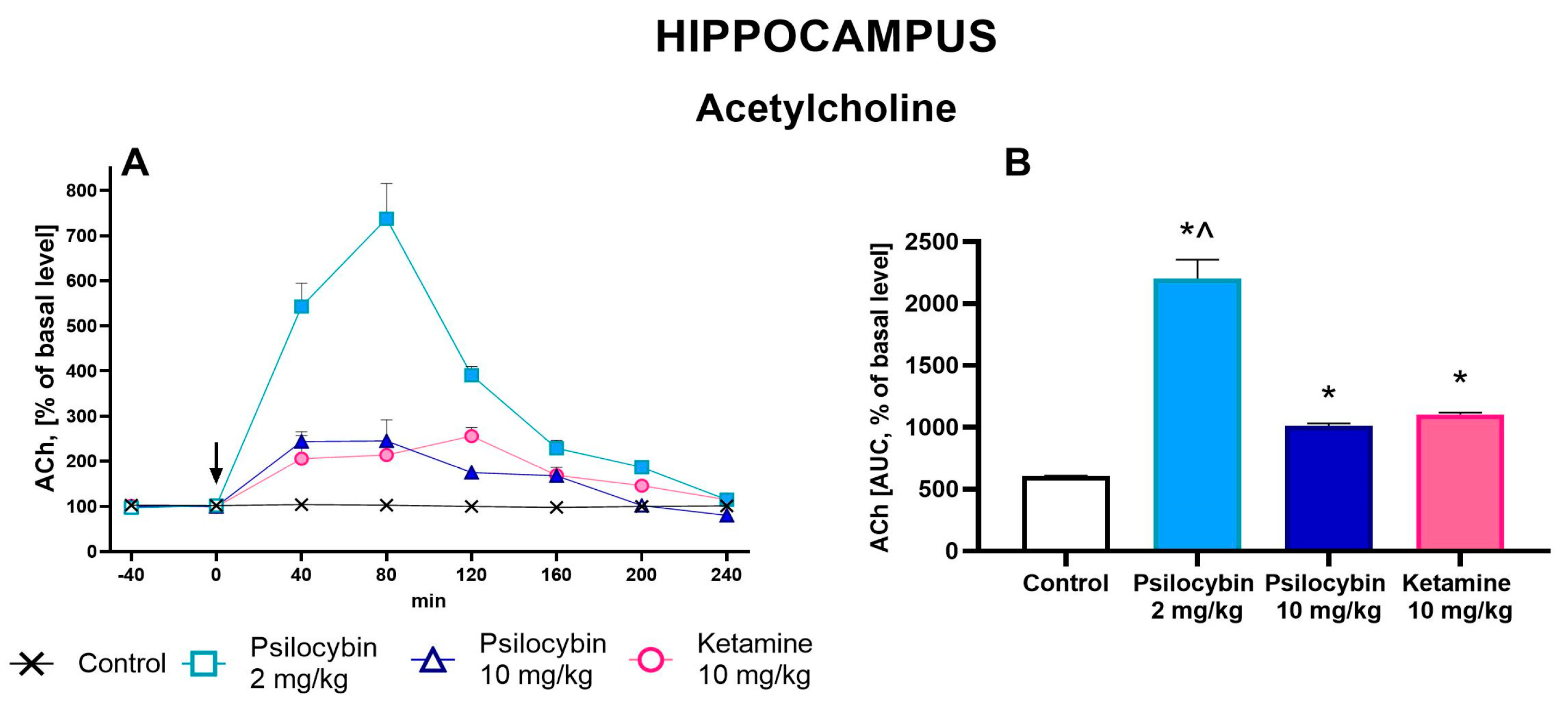
2.4. The Effect of Psilocybin and Ketamine on Extracellular Levels of ACh in the Rat Hippocampus
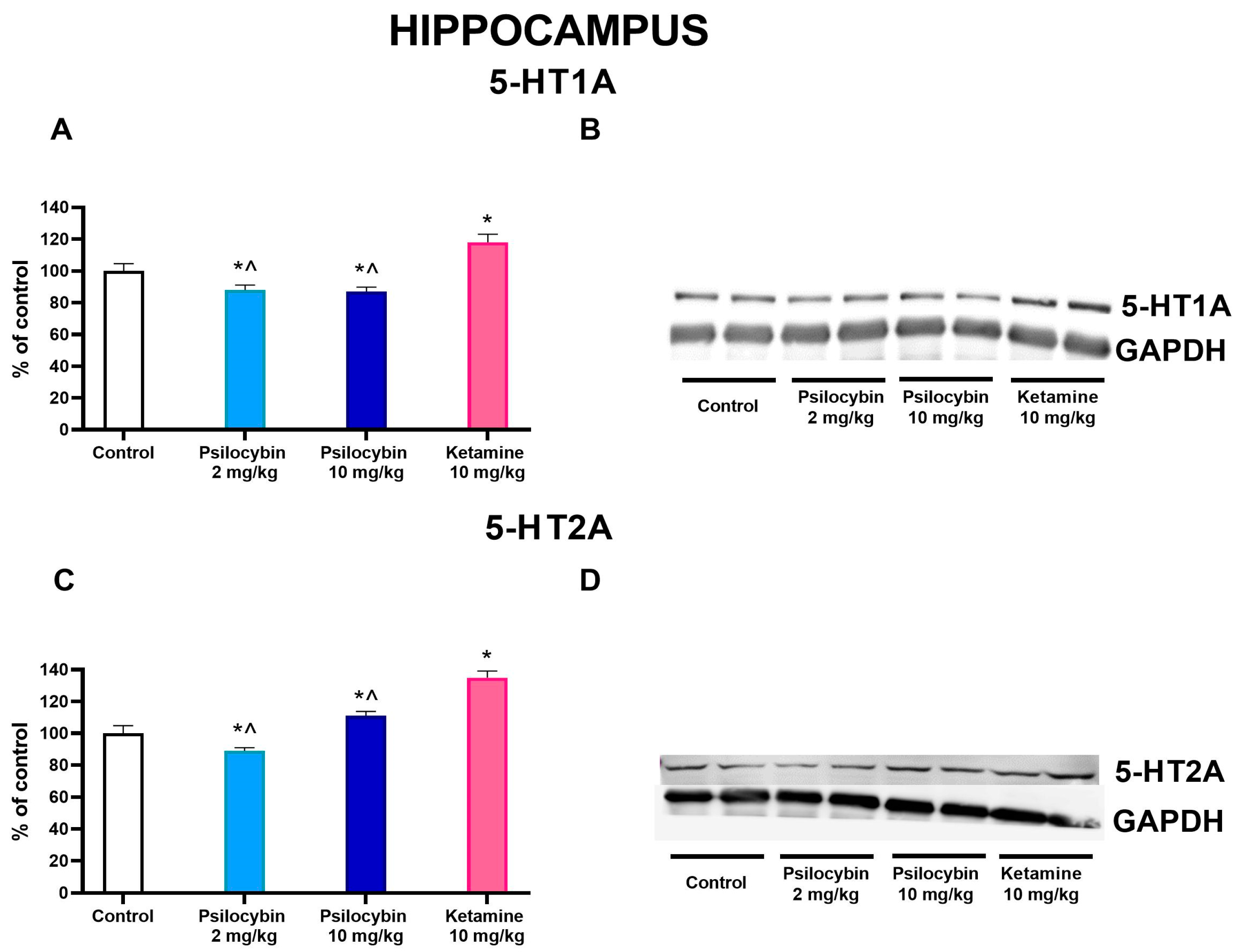
2.5. The Effect of Psilocybin and Ketamine on 5-HT1A and 5-HT2A Receptor Levels in the Rat Hippocampus
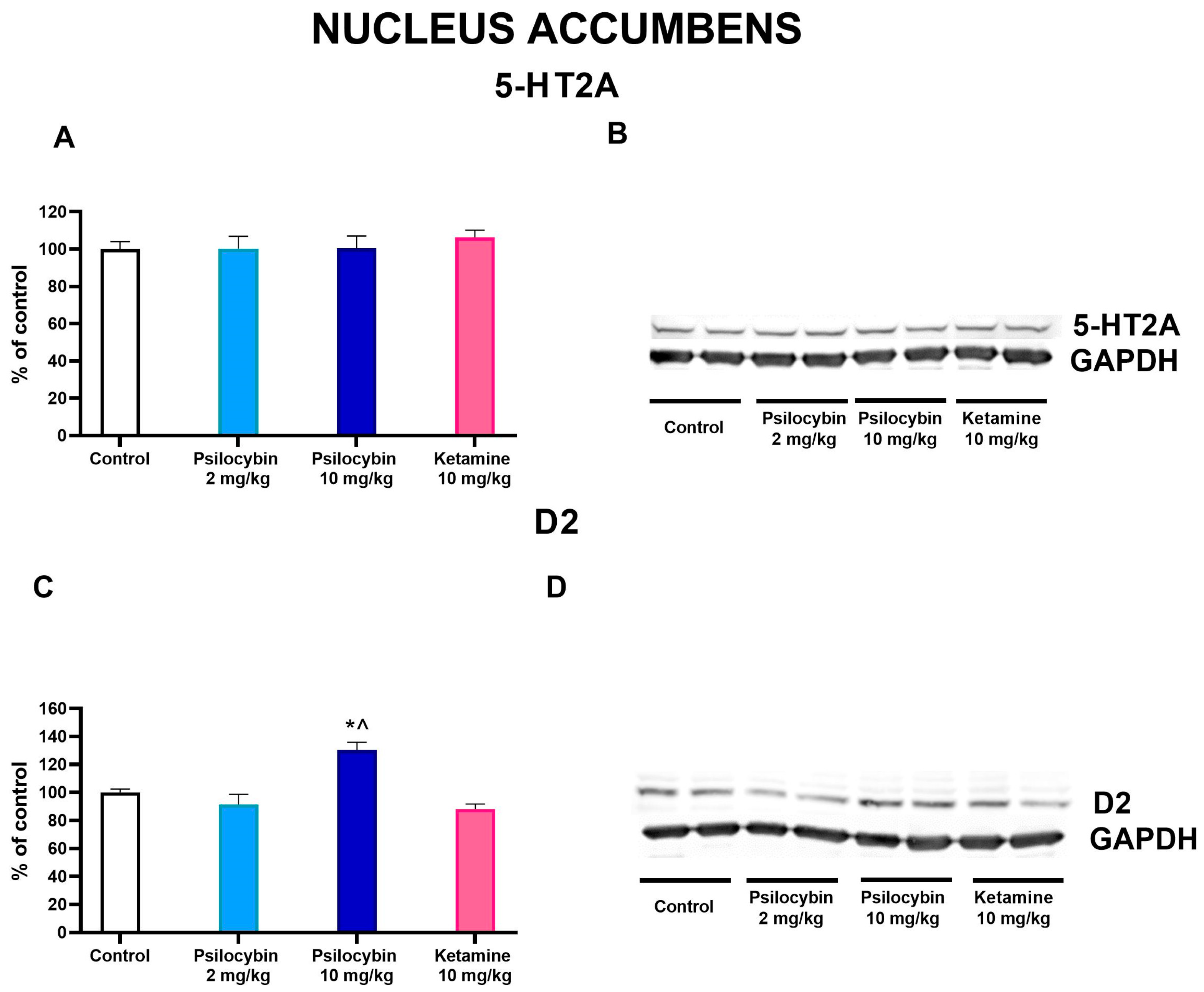
2.6. The Effect of Psilocybin and Ketamine on D2 and 5-HT2A Receptor Levels in the Rat Nucleus Accumbens
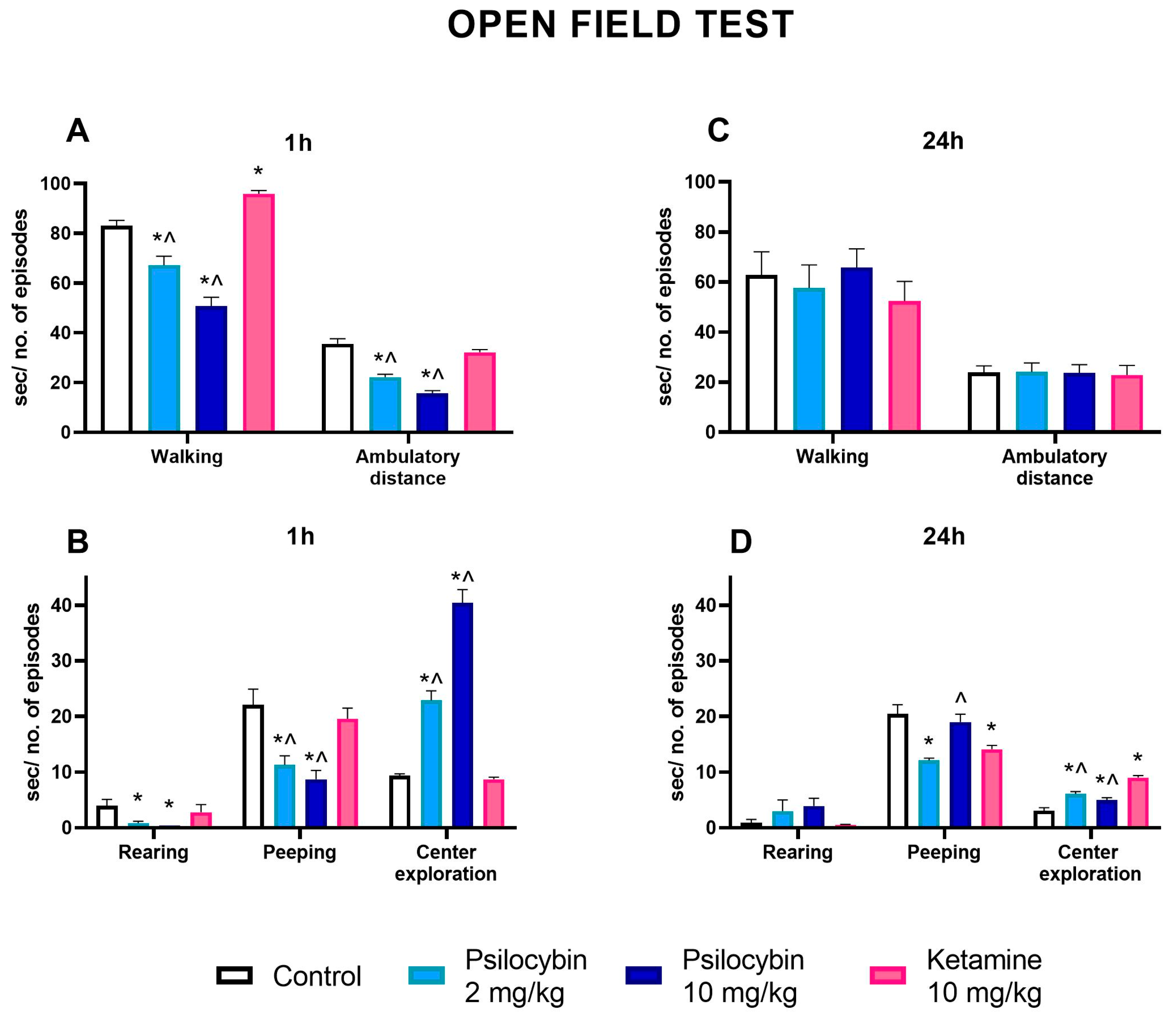
2.7. The Effect of Psilocybin and Ketamine on the Activity of Rats in the Open Field Test
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs and Reagents
4.3. Brain Microdialysis
4.4. Extracellular Concentration of DA, 5-HT, Glutamate, GABA and Acetylcholine
4.5. Western Blotting
4.6. Open Field Test
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Steffen, A.; Nübel, J.; Jacobi, F.; Bätzing, J.; Holstiege, J. Mental and Somatic Comorbidity of Depression: A Comprehensive Cross-Sectional Analysis of 202 Diagnosis Groups Using German Nationwide Ambulatory Claims Data. BMC Psychiatry 2020, 20, 142. [Google Scholar] [CrossRef] [PubMed]
- Chand, S.P.; Arif, H. Depression; StatPearls Publishing: Treasure Island, FL, USA, 2023. Available online: http://www.ncbi.nlm.nih.gov/books/NBK430847 (accessed on 17 July 2023).
- Duman, R.S.; Aghajanian, G.K.; Sanacora, G.; Krystal, J.H. Synaptic plasticity and depression: New insights from stress and rapid-acting antidepressants. Nat. Med. 2016, 22, 238–249. [Google Scholar] [CrossRef] [PubMed]
- Pandya, M.; Altinay, M.; Malone, D.A., Jr.; Anand, A. Where in the brain is depression? Curr. Psychiatry Rep. 2012, 14, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Trifu, S.C.; Trifu, A.C.; Aluaş, E.; Tătaru, M.A.; Costea, R.V. Brain changes in depression. Rom. J. Morphol. Embryol. 2020, 61, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Sheline, Y.I.; Gado, M.H.; Price, J.L. Amygdala core nuclei volumes are decreased in recurrent major depression. NeuroReport 1998, 9, 2023–2028. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, C.G.; Jackowski, A.; Salas, R.; Gupta, S.; Sato, J.R.; Mao, X.; Coplan, J.D.; Shungu, D.C.; Mathew, S.J. The Nucleus Accumbens and Ketamine Treatment in Major Depressive Disorder. Neuropsychopharmacology 2017, 42, 1739–1746. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders; American Psychiatric Association: Arlington, TX, USA, 2013. [Google Scholar] [CrossRef]
- Xu, L.; Nan, J.; Lan, Y. The Nucleus Accumbens: A Common Target in the Comorbidity of Depression and Addiction. Front. Neural Circuits 2020, 14, 37. [Google Scholar] [CrossRef]
- Maeng, S.; Zarate, C.A. The Role of Glutamate in Mood Disorders: Results from the Ketamine in Major Depression Study and the Presumed Cellular Mechanism Underlying Its Antidepressant Effects. Curr. Psychiatry Rep. 2007, 9, 467–474. [Google Scholar] [CrossRef]
- Mathews, D.C.; Henter, I.D.; Zarate, C.A. Targeting the Glutamatergic System to Treat Major Depressive Disorder: Rationale and Progress to Date. Drugs 2012, 72, 1313–1333. [Google Scholar] [CrossRef]
- Yang, C.; Hu, Y.M.; Zhou, Z.Q.; Zhang, G.F.; Yang, J.J. Acute administration of ketamine in rats increases hippocampal BDNF and mTOR levels during forced swimming test. Upsala J. Med. Sci. 2013, 118, 3–8. [Google Scholar] [CrossRef]
- Zhou, Y.; Wu, F.; Liu, W.; Zheng, W.; Wang, C.; Zhan, Y.; Lan, X.; Ning, Y. Volumetric changes in subcortical structures following repeated ketamine treatment in patients with major depressive disorder: A longitudinal analysis. Transl. Psychiatry 2020, 10, 264. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, C.G.; Salas, R.; Jackowski, A.; Baldwin, P.; Sato, J.R.; Mathew, S.J. Hippocampal volume and the rapid antidepressant effect of ketamine. J. Psychopharmacol. 2015, 29, 591–595. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.L.; Wu, F.C.; Wang, C.Y.; Zheng, W.; Lan, X.F.; Deng, X.R.; Ning, Y.P. Relationship between hippocampal volume and inflammatory markers following six infusions of ketamine in major depressive disorder. J. Affect. Disord. 2020, 276, 608–615. [Google Scholar] [CrossRef] [PubMed]
- Hibicke, M.; Landry, A.N.; Kramer, H.M.; Talman, Z.K.; Nichols, C.D. Psychedelics, but Not Ketamine, Produce Persistent Antidepressant-like Effects in a Rodent Experimental System for the Study of Depression. ACS Chem. Neurosci. 2020, 11, 864–871. [Google Scholar] [CrossRef] [PubMed]
- Ly, C.; Greb, A.C.; Cameron, L.P.; Wong, J.M.; Barragan, E.V.; Wilson, P.C.; Burbach, K.F.; Soltanzadeh Zarandi, S.; Sood, A.; Paddy, M.R.; et al. Psychedelics Promote Structural and Functional Neural Plasticity. Cell Rep. 2018, 23, 3170–3182. [Google Scholar] [CrossRef] [PubMed]
- Cameron, L.P.; Benson, C.J.; Dunlap, L.E.; Olson, D.E. Effects of N,N-Dimethyltryptamine on Rat Behaviors Relevant to Anxiety and Depression. ACS Chem. Neurosci. 2018, 9, 1582–1590. [Google Scholar] [CrossRef]
- Hesselgrave, N.; Troppoli, T.A.; Wulff, A.B.; Cole, A.B.; Thompson, S.M. Harnessing psilocybin: Antidepressant-like behavioral and synaptic actions of psilocybin are independent of 5-HT2R activation in mice. Proc. Natl. Acad. Sci. USA 2021, 118, e2022489118. [Google Scholar] [CrossRef]
- Carhart-Harris, R.; Giribaldi, B.; Watts, R.; Baker-Jones, M.; Murphy-Beiner, A.; Murphy, R.; Martell, J.; Blemings, A.; Erritzoe, D.; Nutt, D.J. Trial of Psilocybin versus Escitalopram for Depression. N. Engl. J. Med. 2021, 384, 1402–1411. [Google Scholar] [CrossRef]
- Vollenweider, F.X.; Kometer, M. The neurobiology of psychedelic drugs: Implications for the treatment of mood disorders. Nat. Neurosci. 2010, 11, 642–651. [Google Scholar] [CrossRef]
- Beique, J.C.; Imad, M.; Mladenovic, L.; Gingrich, J.A.; Andrade, R. Mechanism of 5-hydroxytryptamine 2A receptor-mediated facilitation of synaptic activity in prefrontal cortex. Proc. Natl. Acad. Sci. USA 2007, 104, 9870–9875. [Google Scholar] [CrossRef]
- Wojtas, A.; Bysiek, A.; Wawrzczak-Bargiela, A.; Szych, Z.; Majcher-Maślanka, I.; Herian, M.; Maćkowiak, M.; Gołembiowska, K. Effect of Psilocybin and Ketamine on Brain Neurotransmitters, Glutamate Receptors, DNA and Rat Behavior. Int. J. Mol. Sci. 2022, 23, 6713. [Google Scholar] [CrossRef] [PubMed]
- Mason, N.L.; Kuypers, K.P.; Muller, F.; Reckweg, J.; Tse, D.H.Y.; Toennes, S.W.; Hutten, N.R.P.W.; Jansen, J.F.A.; Stiers, P.; Feilding, A.; et al. Me, myself, bye: Regional alterations in glutamate and the experience of ego dissolution with psilocybin. Neuropsychopharmacology 2020, 45, 2003–2011. [Google Scholar] [CrossRef]
- Abdallah, C.G.; Sanacora, G.; Duman, R.S.; Krystal, J.H. The Neurobiology of Depression, Ketamine and Rapid-Acting Antidepressants: Is it Glutamate Inhibition or Activation? Pharmacol. Ther. 2018, 190, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Passie, T.; Seifert, J.; Schneider, U.; Emrich, M.H. The pharmacology of psilocybin. Addict. Biol. 2002, 7, 357–364. [Google Scholar] [CrossRef] [PubMed]
- Rickli, A.; Moning, O.D.; Hoener, M.C.; Liechti, M.E. Receptor interaction profiles of novel psychoactive tryptamines compared with classic hallucinogens. Eur. Neuropsychopharmacol. 2016, 26, 1327–1337. [Google Scholar] [CrossRef] [PubMed]
- Celada, P.; Puig, M.V.; Artigas, F. Serotonin modulation of cortical neurons and networks. Front. Integr. Neurosci. 2013, 7, 25. [Google Scholar] [CrossRef] [PubMed]
- Meccia, J.; Lopez, J.; Bagot, R.C. Probing the antidepressant potential of psilocybin: Integrating insight from human research and animal models towards an understanding of neural circuit mechanisms. Psychopharmacology 2023, 240, 27–40. [Google Scholar] [CrossRef]
- Malenka, R.C.; Nestler, E.J.; Hyman, S.E. Chapter 6: Widely Projecting Systems: Monoamines, Acetylcholine, and Orexin. In Molecular Neuropharmacology: A Foundation for Clinical Neuroscience, 2nd ed.; Sydor, A., Brown, R.Y., Eds.; McGraw-Hill Medical: New York, NY, USA, 2009; pp. 147–148, 154–157. ISBN 9780071481274. [Google Scholar]
- Dailly, E.; Chenu, F.; Renard, C.E.; Bourin, M. Dopamine, depression and antidepressants. Fundam. Clin. Pharmacol. 2004, 18, 601–607. [Google Scholar] [CrossRef]
- De Deurwaerdère, P.; Di Giovanni, G. Serotonergic modulation of the activity of mesencephalic dopaminergic systems: Therapeutic implications. Prog. Neurobiol. 2017, 151, 175–236. [Google Scholar] [CrossRef]
- Sakashita, Y.; Abe, K.; Katagiri, N.; Kambe, T.; Saitoh, T.; Utsunomiya, I.; Horiguchi, Y.; Taguchi, K. Effect of psilocin on extracellular dopamine and serotonin levels in the mesoaccumbens and mesocortical pathway in awake rats. Biol. Pharm. Bull. 2015, 38, 134–138. [Google Scholar] [CrossRef]
- Del Arco, A.; Mora, F. Prefrontal cortex–nucleus accumbens interaction: In vivo modulation by dopamine and glutamate in the prefrontal cortex. Pharmacol. Biochem. Behav. 2008, 90, 226–235. [Google Scholar] [CrossRef] [PubMed]
- Moulédous, L.; Roullet, P.; Guiard, B.P. Brain Circuits Regulated by the 5-HT2A Receptor: Behavioural Consequences on Anxiety and Fear Memory. In 5-HT2A Receptors in the Central Nervous System; Springer: Cham, Switzerland, 2018; pp. 231–258. [Google Scholar] [CrossRef]
- Miller, O.H.; Moran, J.T.; Hall, B.J. Two cellular hypotheses explaining the initiation of ketamine’s antidepressant actions: Direct inhibition and disinhibition. Neuropharmacology 2016, 100, 17–26. [Google Scholar] [CrossRef]
- Ago, Y.; Tanabe, W.; Higuchi, M.; Tsukada, S.; Tanaka, T.; Yamaguchi, T.; Igarashi, H.; Yokoyama, R.; Seiriki, K.; Kasai, A.; et al. (R)-Ketamine Induces a Greater Increase in Prefrontal 5-HT Release Than (S)-Ketamine and Ketamine Metabolites via an AMPA Receptor-Independent Mechanism. Int. J. Neuropsychopharm. 2019, 22, 665–674. [Google Scholar] [CrossRef] [PubMed]
- Chaudhury, D.; Walsh, J.J.; Friedman, A.K.; Juarez, B.; Ku, S.M.; Koo, J.W.; Ferguson, D.; Tsai, H.-C.; Pomeranz, L.; Christoffel, D.J.; et al. Rapid regulation of depression-related behaviours by control of midbrain dopamine neurons. Nature 2013, 493, 532–536. [Google Scholar] [CrossRef] [PubMed]
- Walsh, J.J.; Friedman, A.K.; Sun, H.; Heller, E.A.; Ku, S.M.; Juarez, B.; Burnham, V.L.; Mazei-Robison, M.S.; Ferguson, D.; Golden, S.A.; et al. Stress and CRF gate neural activation of BDNF in the mesolimbic reward pathway. Nat. Neurosci. 2014, 17, 27–29. [Google Scholar] [CrossRef] [PubMed]
- Wook Koo, J.; Labonte, B.; Engmann, O.; Calipari, E.S.; Juarez, B.; Lorsch, Z.; Walsh, J.J.; Friedman, A.K.; Yorgason, J.T.; Han, M.-H.; et al. Essential role of mesolimbic brain-derived neurotrophic factor in chronic social stress-induced depressive behaviors. Biol. Psychiatry 2016, 80, 469–478. [Google Scholar] [CrossRef]
- Vollenweider, F.X.; Smallridge, J.W. Classic Psychedelic Drugs: Update on Biological Mechanisms. Pharmacopsychiatry 2022, 55, 121–138. [Google Scholar] [CrossRef]
- Beliveau, V.; Ganz, M.; Feng, L.; Ozenne, B.; Højgaard, L.; Fisher, P.M.; Svarer, C.; Greve, D.N.; Knudsen, G.M. A High-Resolution In Vivo Atlas of the Human Brain’s Serotonin System. J. Neurosci. 2017, 37, 120–128. [Google Scholar] [CrossRef]
- Zhu, Z.; Wang, G.; Ma, K.; Cui, S.; Wang, J.-H. GABAergic neurons in nucleus accumbens are correlated to resilience and vulnerability to chronic stress for major depression. Oncotarget 2017, 8, 35933–35945. [Google Scholar] [CrossRef]
- Dale, E.; Pehrson, A.L.; Jeyarajah, T.; Li, Y.; Leiser, S.C.; Smagin, G.; Olsen, C.K.; Sanchez, C. Effects of serotonin in the hippocampus: How SSRIs and multimodal antidepressants might regulate pyramidal cell function. CNS Spectr. 2016, 21, 143–161. [Google Scholar] [CrossRef]
- Andrade, R.; Nicoll, R.A. Pharmacologically distinct actions of serotonin on single pyramidal neurons of the rat hippocampus recorded in vitro. J. Physiol. 1987, 394, 99–124. [Google Scholar] [CrossRef] [PubMed]
- Bombardi, C. Neuronal localization of 5-HT2A receptor immunoreactivity in the rat hippocampal region. Brain Res. Bull. 2012, 87, 259–273. [Google Scholar] [CrossRef] [PubMed]
- Abdallah, C.G.; Jackowski, A.; Sato, J.R.; Mao, X.; Kang, G.; Cheema, R.; Coplan, J.D.; Mathew, S.J.; Shungu, D.C. Prefrontal Cortical GABA Abnormalities Are Associated with Reduced Hippocampal Volume in Major Depressive Disorder. Eur. Neuropsychopharmacol. 2015, 25, 1082–1090. [Google Scholar] [CrossRef] [PubMed]
- Picciotto, M.R.; Higley, M.J.; Mineur, Y.S. Acetylcholine as a neuromodulator: Cholinergic signaling shapes nervous system function and behavior. Neuron 2012, 76, 116–129. [Google Scholar] [CrossRef] [PubMed]
- Gielow, M.R.; Zaborszky, L. The Input-Output Relationship of the Cholinergic Basal Forebrain. Cell Rep. 2017, 18, 1817–1830. [Google Scholar] [CrossRef] [PubMed]
- Fujii, T.; Yoshizawa, M.; Nakai, K.; Fujimoto, K.; Suzuki, T.; Kawashima, K. Demonstration of the facilitatory role of 8-OH-DPAT on cholinergic transmission in the rat hippocampus using in vivo microdialysis. Brain Res. 1997, 761, 244–249. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.G.; Gudelsky, G.A. Activation of 5-HT2 receptors enhances the release of acetylcholine in the prefrontal cortex and hippocampus of the rat. Synapse 2004, 53, 202–207. [Google Scholar] [CrossRef] [PubMed]
- Pacheco, A.T.; Olson, R.J.; Garza, G.; Moghaddam, B. Acute psilocybin enhances cognitive flexibility in rats. Neuropsychopharmacology 2023, 48, 1011–1020. [Google Scholar] [CrossRef]
- Almeida, J.R.C.; Versace, A.; Hassel, S.; Kupfer, D.J.; Phillips, M.L. Elevated amygdala activity to sad facial expressions: A state marker of bipolar but not unipolar depression. Biol. Psychiatry 2010, 67, 414–421. [Google Scholar] [CrossRef]
- Barrett, F.S.; Doss, M.K.; Sepeda, N.D.; James, J.; Pekar, J.J.; Griffiths, R.R. Emotions and brain function are altered up to one month after a single high dose of psylocybin. Sci. Rep. 2020, 10, 2214. [Google Scholar] [CrossRef]
- Liu, W.Z.; Huang, S.H.; Wang, Y.; Wang, C.Y.; Pan, H.Q.; Zhao, K.; Hu, P.; Pan, B.X.; Zhang, W.H. Medial prefrontal cortex input to basolateral amygdala controls acute stress-induced short-term anxiety-like behavior in mice. Neuropsychopharmacology 2023, 48, 734–744. [Google Scholar] [CrossRef] [PubMed]
- Pompeiano, M.; Palacios, J.M.; Mengod, G. Distribution of serotonin 5-HT2 receptor family mRNAs: Comparison between 5-HT2A and 5-HT2C receptors. Mol. Brain Res. 1994, 23, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Cornea-Hebert, V.; Riad, M.; Wu, C.; Singh, S.K.; Descarries, L. Cellular and subcellular distribution of the serotonin 5-HT2A receptor in the central nervous system of adult rat. J. Comp. Neurol. 1999, 409, 187–209. [Google Scholar] [CrossRef]
- Nikolaus, S.; Müller, H.-W.; Hautzel, H. Different patterns of 5-HT receptor and transporter dysfunction in neuropsychiatric disorders—A comparative analysis of in vivo imaging findings. Rev. Neurosci. 2016, 27, 27–59. [Google Scholar] [CrossRef] [PubMed]
- Raval, N.R.; Johansen, A.; Donovan, L.L.; Ros, N.F.; Ozenne, B.; Hansen, H.D.; Knudsen, G.M. A Single Dose of Psilocybin Increases Synaptic Density and Decreases 5-HT2A Receptor Density in the Pig Brain. Int. J. Mol. Sci. 2021, 22, 835. [Google Scholar] [CrossRef] [PubMed]
- Wedzony, K.; Maćkowiak, M.; Czyrak, A.; Fijał, K.; Michalska, B. Single doses of MK-801, a non-competitive antagonist of NMDA receptors, increase the number of 5-HT1A serotonin receptors in the rat brain. Brain Res. 1997, 756, 84–91. [Google Scholar] [CrossRef]
- Jefsen, O.; Højgaard, K.; Christiansen, S.L.; Elfving, B.; Nutt, D.J.; Wegener, G.; Müller, H.K. Psilocybin Lacks Antidepressant-like Effect in the Flinders Sensitive Line Rat. Acta Neuropsychiatr. 2019, 31, 213–219. [Google Scholar] [CrossRef]
- Shirota, O.; Hakamata, W.; Goda, Y. Concise Large-Scale Synthesis of Psilocin and Psilocybin, Principal Hallucinogenic Constituents of “Magic Mushroom”. J. Nat. Prod. 2003, 66, 885–887. [Google Scholar] [CrossRef]
- Popik, P.; Hogendorf, A.; Bugno, R.; Khoo, S.Y.S.; Zajdel, P.; Malikowska-Racia, N.; Nikiforuk, A.; Golebiowska, J. Effects of Ketamine Optical Isomers, Psilocybin, Psilocin and Norpsilocin on Time Estimation and Cognition in Rats. Psychopharmacology 2022, 239, 1689–1703. [Google Scholar] [CrossRef]
- Paxinos, G.; Watson, G. The Rat Brain in Stereotaxic Coordinates; Academic Press: San Diego, CA, USA, 1998. [Google Scholar]
- Herian, M.; Skawski, M.; Wojtas, A.; Sobocińska, M.K.; Noworyta, K.; Gołembiowska, K. Tolerance to Neurochemical and Behavioral Effects of the Hallucinogen 25I-NBOMe. Psychopharmacology 2021, 238, 2349–2364. [Google Scholar] [CrossRef]
- Wojtas, A.; Herian, M.; Skawski, M.; Sobocińska, M.; González-Marín, A.; Noworyta-Sokołowska, K.; Gołembiowska, K. Neurochemical and Behavioral Effects of a New Hallucinogenic Compound 25B-NBOMe in Rats. Neurotox. Res. 2021, 39, 305–326. [Google Scholar] [CrossRef] [PubMed]
- Rogóz, Z.; Skuza, G. Anxiolytic-like Effects of Olanzapine, Risperidone and Fluoxetine in the Elevated plus-Maze Test in Rats. Pharmacol. Rep. 2011, 63, 1547–1552. [Google Scholar] [CrossRef] [PubMed]
- Shao, L.-X.; Liao, C.; Gregg, I.; Davoudian, P.A.; Savalia, N.K.; Delagarza, K.; Kwan, A.C. Psilocybin induces rapid and persistent growth of dendritic spines in frontal cortex in vivo. Neuron 2021, 109, 2535–2544. [Google Scholar] [CrossRef] [PubMed]
- Tylš, F.; Páleniček, T.; Kadeřabek, L.; Lipski, M.; Kubešová, A.; Horáček, J. Sex differences and serotonergic mechanisms in the behavioural effects of psilocin. Behav. Pharmacol. 2016, 27, 309–320. [Google Scholar] [CrossRef]



Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wojtas, A.; Bysiek, A.; Wawrzczak-Bargiela, A.; Maćkowiak, M.; Gołembiowska, K. Limbic System Response to Psilocybin and Ketamine Administration in Rats: A Neurochemical and Behavioral Study. Int. J. Mol. Sci. 2024, 25, 100. https://doi.org/10.3390/ijms25010100
Wojtas A, Bysiek A, Wawrzczak-Bargiela A, Maćkowiak M, Gołembiowska K. Limbic System Response to Psilocybin and Ketamine Administration in Rats: A Neurochemical and Behavioral Study. International Journal of Molecular Sciences. 2024; 25(1):100. https://doi.org/10.3390/ijms25010100
Chicago/Turabian StyleWojtas, Adam, Agnieszka Bysiek, Agnieszka Wawrzczak-Bargiela, Marzena Maćkowiak, and Krystyna Gołembiowska. 2024. "Limbic System Response to Psilocybin and Ketamine Administration in Rats: A Neurochemical and Behavioral Study" International Journal of Molecular Sciences 25, no. 1: 100. https://doi.org/10.3390/ijms25010100