

Hyper-Production of Pullulan by a Novel Fungus of Aureobasidium melanogenum ZH27 through Batch Fermentation


Abstract
:1. Introduction
2. Results
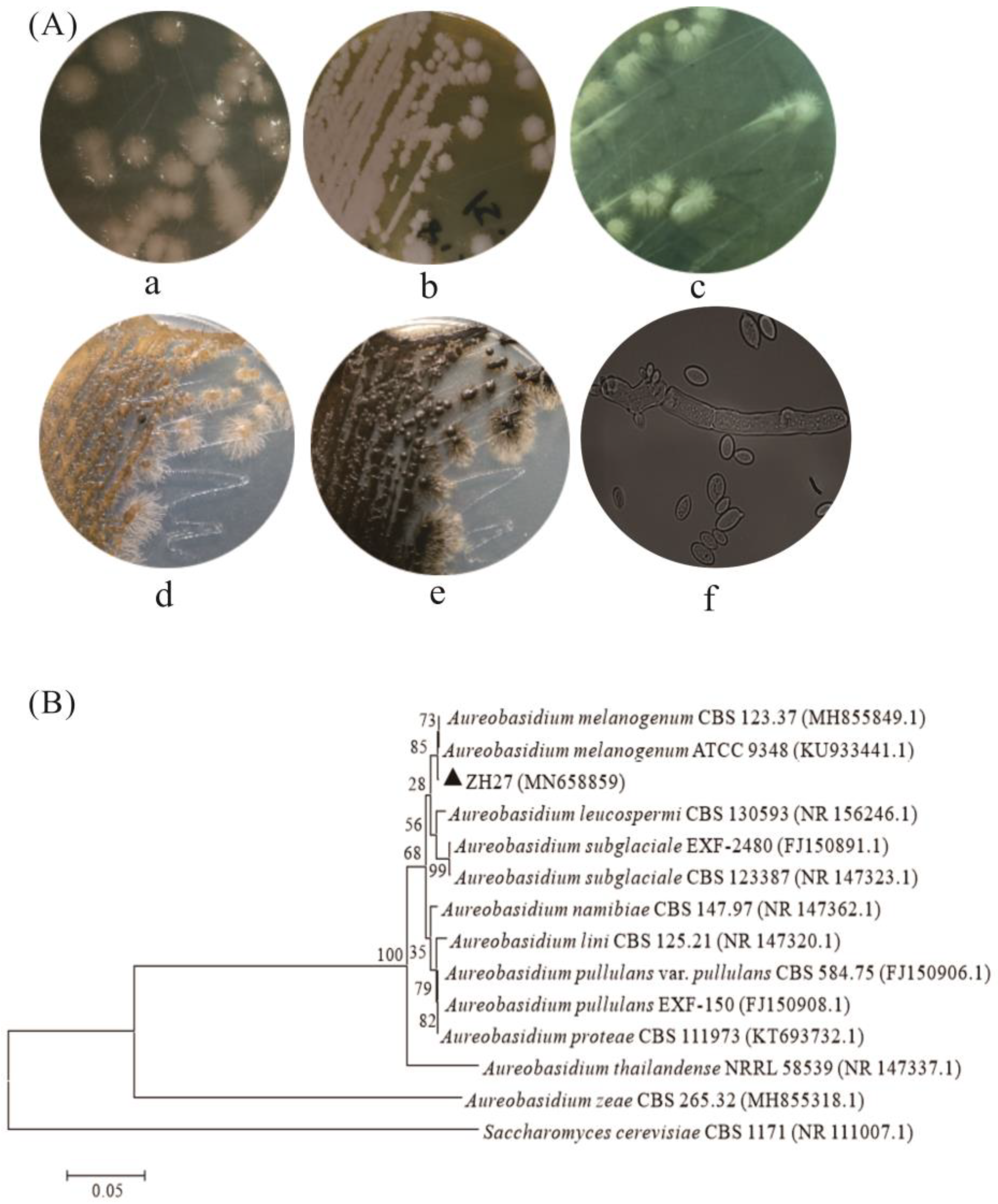
2.1. Screening and Identification of EPS-Producing Strains
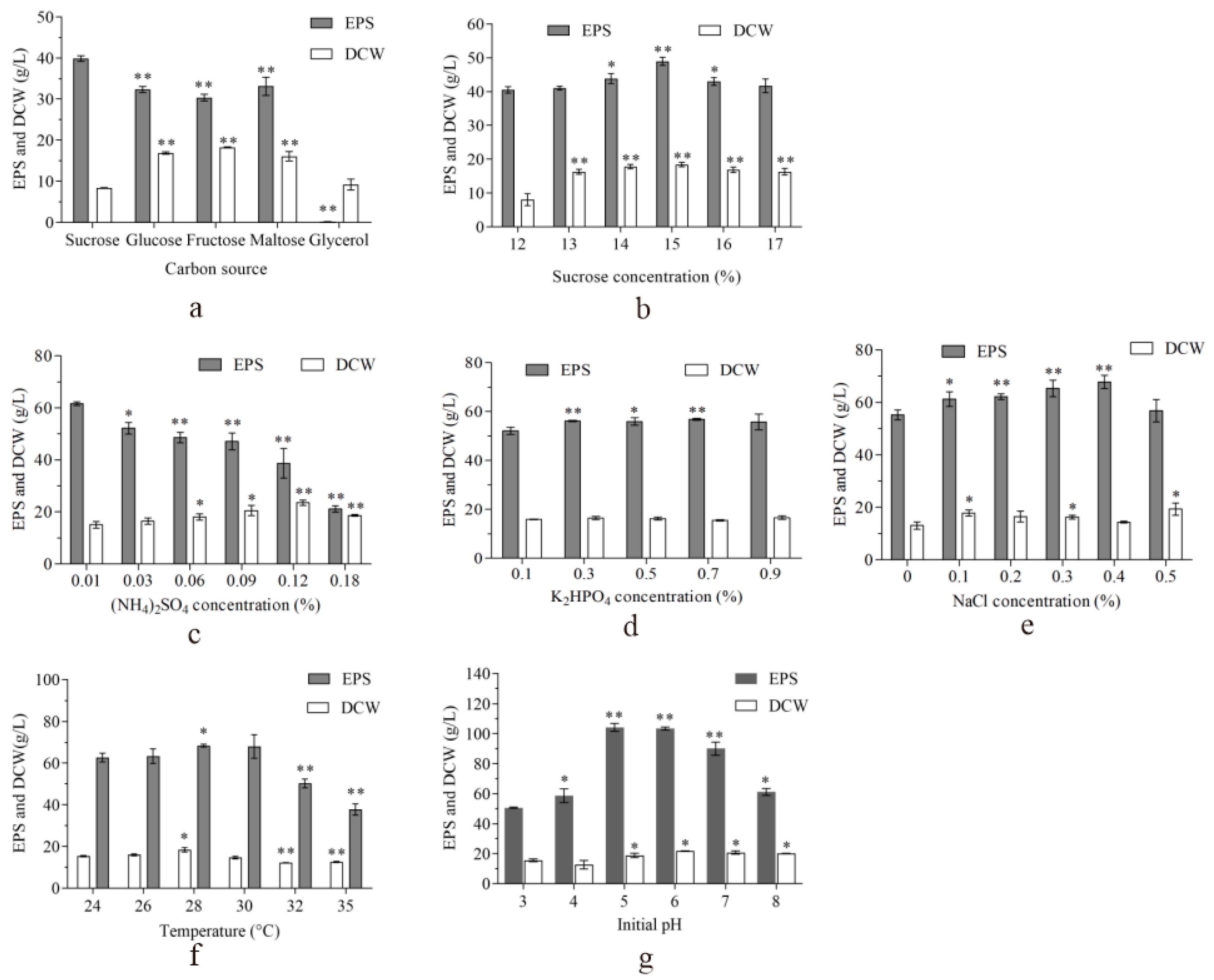
2.2. Optimization for EPS Production
2.3. Structural Analysis of EPS
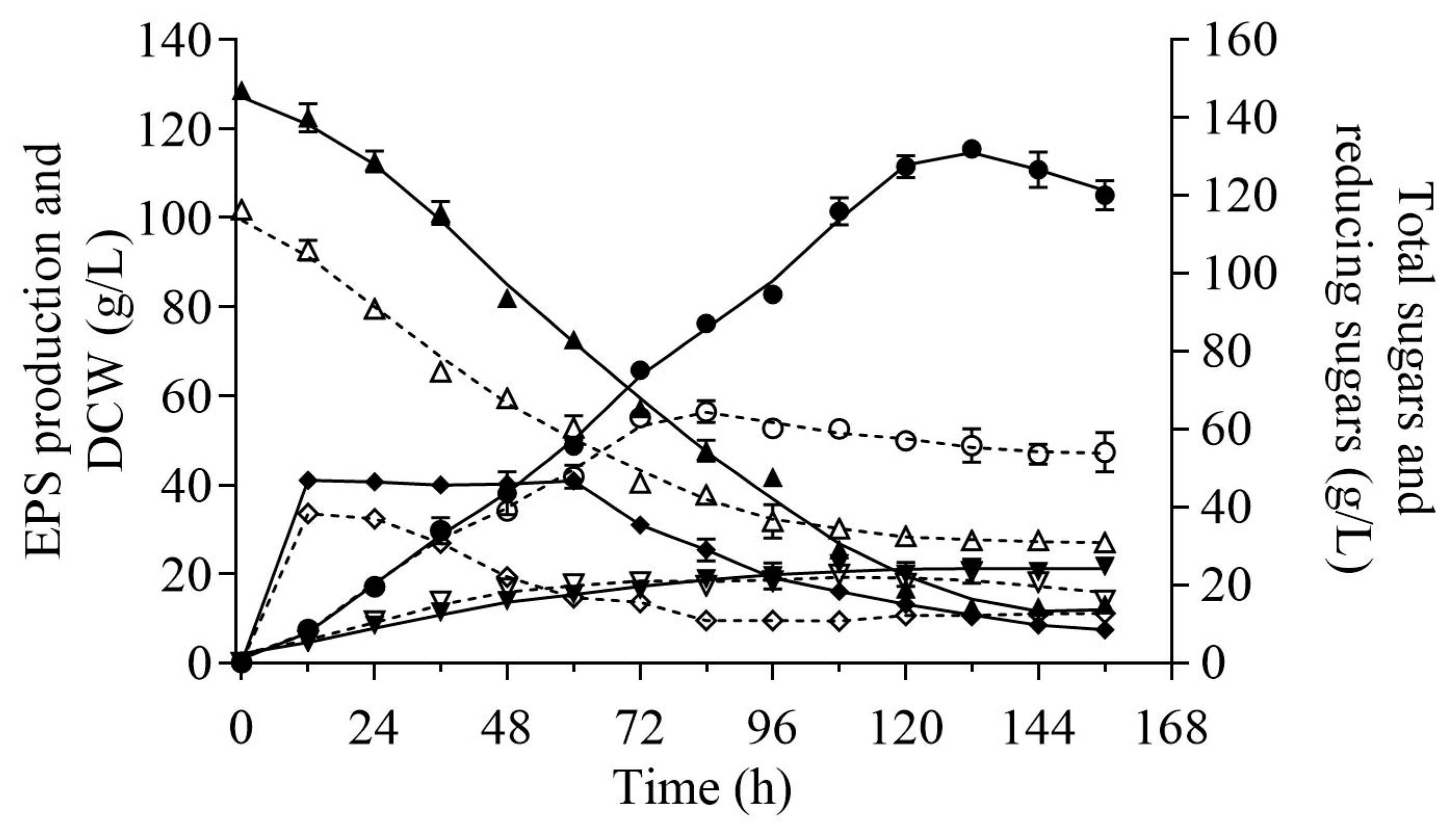
2.4. Comparison of Pullulan Production between the IC and OC
2.5. Probable Reasons for Hyper-Production of Pullulan
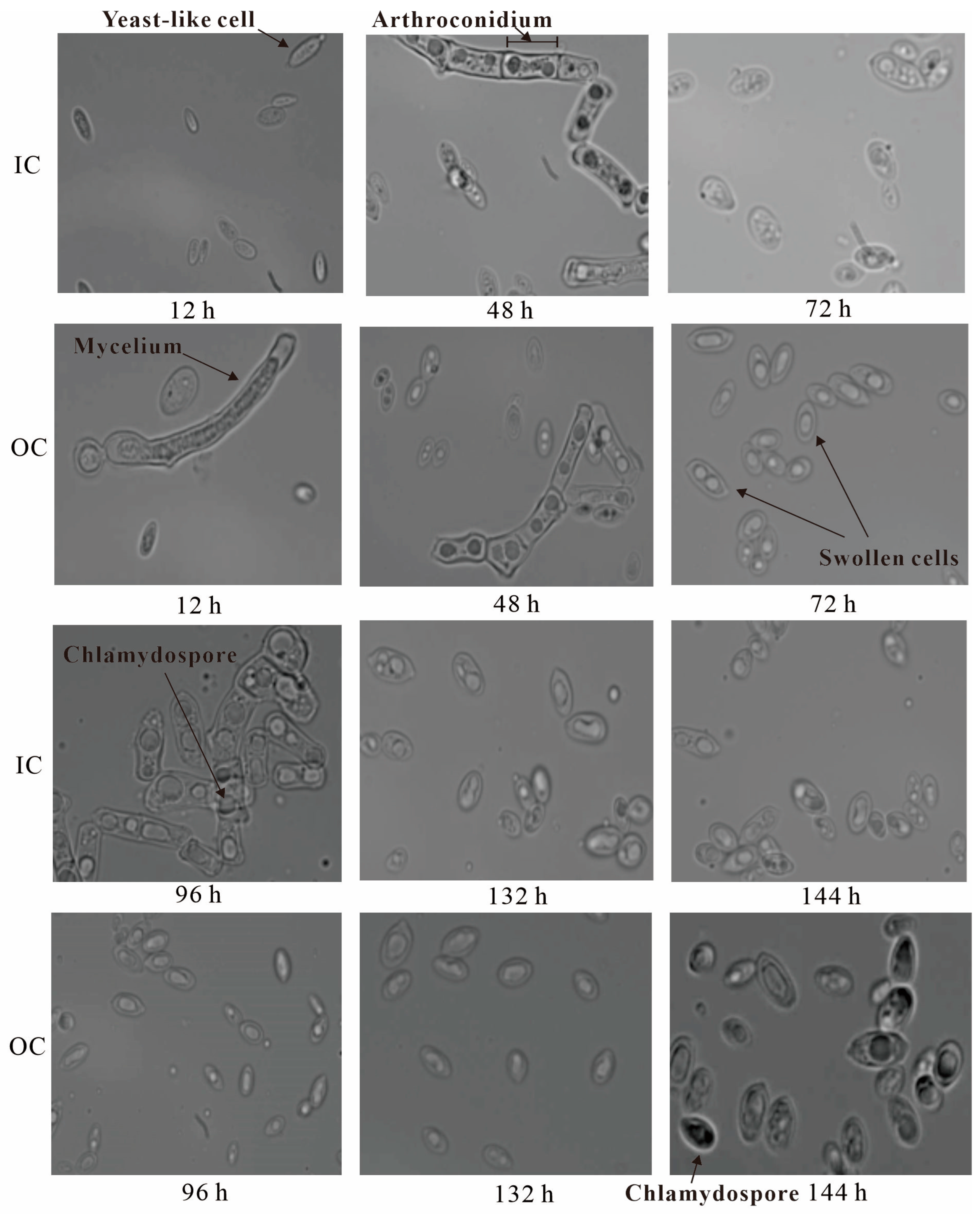
2.5.1. Dynamic Changes in Cell Morphology during Batch Fermentation
2.5.2. Expression Changes of the Genes in Pullulan Synthesis and Carbon Metabolism
3. Discussion
3.1. Strain ZH27 Is a Promising Candidate for Pullulan Synthesis
3.2. Robust Pullulan Synthesis by Strain ZH27 after Optimization
3.3. Swollen Cells, By-Products, and High Expressions of Related Genes May Be Required for Efficient Pullulan Production
4. Materials and Methods
4.1. Sample Collection and Screening for EPS-Producing Strains
4.2. Strain Identification
4.3. Optimization for High-Level EPS Production
4.4. Determination of EPS and DCW
4.5. Purification and Identification of EPS
4.6. Batch Fermentation for Pullulan Production
4.7. Determination of Reducing Sugars, Total Sugars, Cell Morphology, Viscosity, and pH Values
4.8. Detection of Transcriptional Expressions of the Genes Related to Pullulan Synthesis
4.9. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Wei, X.; Liu, G.L.; Jia, S.L.; Chi, Z.; Hu, Z.; Chi, Z.M. Pullulan biosynthesis and its regulation in Aureobasidium spp. Carbohydr. Polym. 2021, 251, 117076. [Google Scholar] [CrossRef]
- Hamidi, M.; Kennedy, J.F.; Khodaiyan, F.; Mousavi, Z.; Hosseini, S.S. Production optimization, characterization and gene expression of pullulan from a new strain of Aureobasidium pullulans. Int. J. Biol. Macromol. 2019, 138, 725–735. [Google Scholar] [CrossRef]
- Haghighatpanah, N.; Mirzaee, H.; Khodaiyan, F.; Kennedy, J.F.; Aghakhani, A.; Hosseini, S.S.; Jahanbin, K. Optimization and characterization of pullulan produced by a newly identified strain of Aureobasidium pullulans. Int. J. Biol. Macromol. 2020, 152, 305–313. [Google Scholar] [CrossRef]
- Singh, R.S.; Kaur, N.; Hassan, M.; Kennedy, J.F. Pullulan in biomedical research and development-A review. Int. J. Biol. Macromol. 2021, 166, 694–706. [Google Scholar] [CrossRef]
- Singh, R.; Gaur, R.; Bansal, S.; Biswas, P.; Pandey, P.K.; Jamal, F.; Tiwari, S.; Gaur, M.K. Aureobasidium pullulans—An industrially important pullulan producing black yeast. Int. J. Curr. Microbiol. App. Sci. 2015, 4, 605–622. [Google Scholar]
- Singh, R.S.; Kaur, N.; Kennedy, J.F. Pullulan production from agro-industrial waste and its applications in food industry: A review. Carbohydr. Polym. 2019, 217, 46–57. [Google Scholar] [CrossRef]
- Sugumaran, K.; Ponnusami, V. Review on production, downstream processing and characterization of microbial pullulan. Carbohydr. Polym. 2017, 173, 573–591. [Google Scholar]
- Jiang, H.; Chen, T.J.; Chi, Z.; Hu, Z.; Liu, G.L.; Sun, Y.; Zhang, S.H.; Chi, Z.M. Macromolecular pullulan produced by Aureobasidium melanogenum 13-2 isolated from the Taklimakan desert and its crucial roles in resistance to the stress treatments. Int. J. Biol. Macromol. 2019, 135, 429–436. [Google Scholar] [CrossRef]
- Liu, F.; Zhang, J.; Zhang, L.; Diao, M.; Ling, P.; Wang, F. Correlation between the synthesis of pullulan and melanin in Aureobasidium pullulans. Int. J. Biol. Macromol. 2021, 177, 252–260. [Google Scholar] [CrossRef]
- Chen, G.; Zhu, Y.; Zhang, G.; Liu, H.; Wei, Y.; Wang, P.; Wang, F.; Xian, M.; Xiang, H.; Zhang, H. Optimization and characterization of pullulan production by a newly isolated high-yielding strain Aureobasidium melanogenum. Prep. Biochem. Biotechnol. 2019, 49, 557–566. [Google Scholar] [CrossRef]
- Chen, S.; Zheng, H.; Gao, J.; Song, H.; Bai, W. High-level production of pullulan and its biosynthesis regulation in Aureobasidium pullulans BL06. Front. Bioeng. Biotechnol. 2023, 11, 1131875. [Google Scholar] [CrossRef]
- Ma, Z.-C.; Fu, W.-J.; Liu, G.-L.; Wang, Z.-P.; Chi, Z.-M. High-level pullulan production by Aureobasidium pullulans var. melanogenium P16 isolated from mangrove system. Appl. Microbiol. Biotechnol. 2014, 98, 4865–4873. [Google Scholar] [CrossRef]
- Jiang, H.; Xue, S.-J.; Li, Y.-F.; Liu, G.-L.; Chi, Z.-M.; Hu, Z.; Chi, Z. Efficient transformation of sucrose into high pullulan concentrations by Aureobasidium melanogenum TN1-2 isolated from a natural honey. Food Chem. 2018, 257, 29–35. [Google Scholar] [CrossRef]
- Li, X.; Zhao, S.; Chen, L.; Zhou, Q.; Qiu, J.; Xin, X.; Zhang, Y.; Yuan, W.; Tian, C.; Yang, J.; et al. High-level production of pullulan from high concentration of glucose by mutagenesis and adaptive laboratory evolution of Aureobasidium pullulans. Carbohydr. Polym. 2023, 302, 120426. [Google Scholar] [CrossRef]
- Chen, T.-J.; Liu, G.-L.; Wei, X.; Wang, K.; Hu, Z.; Chi, Z.; Chi, Z.-M. A multidomain α-glucan synthetase 2 (AmAgs2) is the key enzyme for pullulan biosynthesis in Aureobasidium melanogenum P16. Int. J. Biol. Macromol. 2020, 150, 1037–1045. [Google Scholar] [CrossRef]
- Wang, G.-L.; Din, A.U.; Qiu, Y.-S.; Wang, C.-L.; Wang, D.-H.; Wei, G.-Y. Triton X-100 improves co-production of β-1, 3-D-glucan and pullulan by Aureobasidium pullulans. Appl. Microbiol. Biotechnol. 2020, 104, 10685–10696. [Google Scholar] [CrossRef]
- Chen, X.; Wang, Y.; Zhang, X.-T.; Wu, Y.-N.; Zhang, X.-L.; Zhang, G.-C.; Wang, C.-L.; Zou, X.; Wang, D.-H.; Wei, G.-Y. MAL31, a sugar transporter involved in pullulan biosynthesis in Aureobasidium pullulans. J. Biotechnol. 2022, 359, 176–184. [Google Scholar] [CrossRef]
- Xue, S.-J.; Chen, L.; Jiang, H.; Liu, G.-L.; Chi, Z.-M.; Hu, Z.; Chi, Z. High pullulan biosynthesis from high concentration of glucose by a hyperosmotic resistant, yeast-like fungal strain isolated from a natural comb-honey. Food Chem. 2019, 286, 123–128. [Google Scholar] [CrossRef]
- Yang, J.; Zhang, Y.; Zhao, S.; Zhou, Q.; Xin, X.; Chen, L. Statistical optimization of medium for pullulan production by Aureobasidium pullulans NCPS2016 using fructose and soybean meal hydrolysates. Molecules 2018, 23, 1334. [Google Scholar] [CrossRef]
- Chen, L.; Wei, X.; Liu, G.-L.; Hu, Z.; Chi, Z.-M.; Chi, Z. Glycerol, trehalose and vacuoles had relations to pullulan synthesis and osmotic tolerance by the whole genome duplicated strain Aureobasidium melanogenum TN3-1 isolated from natural honey. Int. J. Biol. Macromol. 2020, 165, 131–140. [Google Scholar] [CrossRef]
- De Paula, N.M.; da Silva, K.; Brugnari, T.; Haminiuk, C.W.I.; Maciel, G.M. Biotechnological potential of fungi from a mangrove ecosystem: Enzymes, salt tolerance and decolorization of a real textile effluent. Microbiol. Res. 2022, 254, 126899. [Google Scholar] [CrossRef] [PubMed]
- Zeng, N.; Zhang, N.; Ma, X.; Wang, Y.; Zhang, Y.; Wang, D.; Pu, F.; Li, B. Transcriptomics Integrated with Metabolomics: Assessing the Central Metabolism of Different Cells after Cell Differentiation in Aureobasidium pullulans NG. J. Fungi 2022, 8, 882. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Zhao, T.; Zhuo, J.; Zhan, C.; Zhang, F.; Linhardt, R.J.; Bai, Z.; Yang, Y. MAPK/HOG signaling pathway induced stress-responsive damage repair is a mechanism for Pichia pastoris to survive from hyperosmotic stress. J. Chem. Technol. Biotechnol. 2021, 96, 412–422. [Google Scholar] [CrossRef]
- Peng, M.; Liang, Z. Degeneration of industrial bacteria caused by genetic instability. World J. Microbiol. Biotechnol. 2020, 36, 119. [Google Scholar] [CrossRef] [PubMed]
- Wang, Q.-Q.; Lu, Y.; Ren, Z.-Y.; Chi, Z.; Liu, G.-L.; Chi, Z.-M. CreA is directly involved in pullulan biosynthesis and regulation of Aureobasidium melanogenum P16. Curr. Genet. 2017, 63, 471–485. [Google Scholar] [CrossRef] [PubMed]
- Kang, X.-X.; Jia, S.-L.; Wei, X.; Zhang, M.; Liu, G.-L.; Hu, Z.; Chi, Z.; Chi, Z.-M. Liamocins biosynthesis, its regulation in Aureobasidium spp., and their bioactivities. Crit. Rev. Biotechnol. 2022, 42, 93–105. [Google Scholar] [CrossRef] [PubMed]
- Meneses, D.P.; Gudiña, E.J.; Fernandes, F.; Gonçalves, L.R.; Rodrigues, L.R.; Rodrigues, S. The yeast-like fungus Aureobasidium thailandense LB01 produces a new biosurfactant using olive oil mill wastewater as an inducer. Microbiol. Res. 2017, 204, 40–47. [Google Scholar] [CrossRef]
- Wani, S.M.; Masoodi, F.; Mir, S.A.; Khanday, F.A. Pullulan production by Aureobasidium pullulans MTCC 1991 from apple pomace and its characterization. Food Biosci. 2023, 51, 102254. [Google Scholar] [CrossRef]
- Choudhury, A.R.; Saluja, P.; Prasad, G. Pullulan production by an osmotolerant Aureobasidium pullulans RBF-4A3 isolated from flowers of Caesulia axillaris. Carbohydr. Polym. 2011, 83, 1547–1552. [Google Scholar] [CrossRef]
- Wang, D.; Chen, F.; Wei, G.; Jiang, M.; Dong, M. The mechanism of improved pullulan production by nitrogen limitation in batch culture of Aureobasidium pullulans. Carbohydr. Polym. 2015, 127, 325–331. [Google Scholar] [CrossRef]
- Kang, X.-X.; Wang, Q.-Q.; Chi, Z.; Liu, G.-L.; Hu, Z.; Chi, Z.-M. The GATA type transcriptional factors regulate pullulan biosynthesis in Aureobasidium melanogenum P16. Int. J. Biol. Macromol. 2021, 192, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Shen, Q.; Zhang, D.; Hao, Y.; Luo, X.; Yang, J.; Wei, B.; Li, O.; Zhao, H. Optimum conditions and identification of pullulan produced by a new finding strain Aureobasidium pullulans RM1603. Lett. Biotechnol. 2019, 30, 385–390. [Google Scholar]
- Wang, D.-H.; Ni, T.-F.; Ju, X.-M.; Wei, G.-Y. Sodium chloride improves pullulan production by Aureobasidium pullulans but reduces the molecular weight of pullulan. Appl. Microbiol. Biotechnol. 2018, 102, 8921–8930. [Google Scholar] [CrossRef] [PubMed]
- Hohmann, S. An integrated view on a eukaryotic osmoregulation system. Curr. Genet. 2015, 61, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Wu, S.; Chen, H.; Jin, Z.; Tong, Q. Effect of two-stage temperature on pullulan production by Aureobasidium pullulans. World J. Microbiol. Biotechnol. 2010, 26, 737–741. [Google Scholar] [CrossRef]
- Hilares, R.T.; Resende, J.; Orsi, C.A.; Ahmed, M.A.; Lacerda, T.M.; da Silva, S.S.; Santos, J.C. Exopolysaccharide (pullulan) production from sugarcane bagasse hydrolysate aiming to favor the development of biorefineries. Int. J. Biol. Macromol. 2019, 127, 169–177. [Google Scholar] [CrossRef]
- Jo, H.M.; Kim, Y.J.; Yoo, S.H.; Kim, C.M.; Kim, K.W.; Park, C.S. Optimization of cultivation conditions for pullulan production from Aureobasidium pullulans MR by response surface methodology. Korean J. Food Sci. Technol. 2021, 53, 195–203. [Google Scholar]
- Xue, S.-J.; Jiang, H.; Chen, L.; Ge, N.; Liu, G.-L.; Hu, Z.; Chi, Z.-M.; Chi, Z. Over-expression of Vitreoscilla hemoglobin (VHb) and flavohemoglobin (FHb) genes greatly enhances pullulan production. Int. J. Biol. Macromol. 2019, 132, 701–709. [Google Scholar] [CrossRef]
- Chen, L.; Chi, Z.; Liu, G.L.; Xue, S.J.; Wang, Z.P.; Hu, Z.; Chi, Z.M. Improved pullulan production by a mutant of Aureobasidium melanogenum TN3-1 from a natural honey and capsule shell preparation. Int. J. Biol. Macromol. 2019, 141, 268–277. [Google Scholar] [CrossRef]
- Zeng, N.; Zhang, N.; Wang, D.; Long, J.; Wang, Y.; Zhang, Y.; Pu, F.; Li, Z.; Baloch, F.B.; Li, B. Regulation of cell differentiation to promote pullulan synthesis in Aureobasidium pullulans NG. Appl. Microbiol. Biotechnol. 2023, 107, 6761–6773. [Google Scholar] [CrossRef]
- Liu, X.; Wang, H.; Zhang, N.; Li, B. Exploration of cell polymorphisms and factors influencing cell differentiation of Aureobasidium pullulans. Microbiol. China 2019, 46, 1461–1469. [Google Scholar]
- Zhang, K.; Lin, C.; Zhao, S.; Wang, W.; Zhou, W.; Ru, X.; Cong, H.; Yang, Q. The role of pH transcription factor Appacc in upregulation of pullulan biosynthesis in Aureobasidium pullulans using potato waste as a substrate. Int. J. Biol. Macromol. 2023, 242, 124797. [Google Scholar] [CrossRef] [PubMed]
- Zou, X.; Cheng, C.; Feng, J.; Song, X.; Lin, M.; Yang, S.T. Biosynthesis of polymalic acid in fermentation: Advances and prospects for industrial application. Crit. Rev. Biotechnol. 2019, 39, 408–421. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Huang, S.; Chen, Y.; Guo, X.; Xiao, D. Discovering the role of the apolipoprotein gene and the genes in the putative pullulan biosynthesis pathway on the synthesis of pullulan, heavy oil and melanin in Aureobasidium pullulans. World J. Microbiol. Biotechnol. 2018, 34, 11. [Google Scholar] [CrossRef] [PubMed]
- Khatun, M.S.; Harrison, M.D.; Speight, R.E.; O’Hara, I.M.; Zhang, Z. Efficient production of fructo-oligosaccharides from sucrose and molasses by a novel Aureobasidium pullulan strain. Biochem. Eng. J. 2020, 163, 107747. [Google Scholar] [CrossRef]
- Chi, Z.; Liu, N.-N.; Jiang, H.; Wang, Q.-Q.; Chen, J.-T.; Liu, G.-L.; Hu, Z.; Chi, Z.-M. Relationship between β-D-fructofuranosidase activity, fructooligosaccharides and pullulan biosynthesis in Aureobasidium melanogenum P16. Int. J. Biol. Macromol. 2019, 125, 1103–1111. [Google Scholar] [CrossRef]
- Sheng, L.; Tong, Q.; Ma, M. Why sucrose is the most suitable substrate for pullulan fermentation by Aureobasidium pullulans CGMCC1234? Enzym. Microb. Technol. 2016, 92, 49–55. [Google Scholar] [CrossRef]
- Yaakoub, H.; Sanchez, N.S.; Ongay-Larios, L.; Courdavault, V.; Calenda, A.; Bouchara, J.P.; Coria, R.; Papon, N. The high osmolarity glycerol (HOG) pathway in fungi(dagger). Crit. Rev. Microbiol. 2022, 48, 657–695. [Google Scholar] [CrossRef]
- Zhang, G.; Wang, G.; Zhu, C.; Wang, C.; Wang, D.; Wei, G. Metabolic flux and transcriptome analyses provide insights into the mechanism underlying zinc sulfate improved β-1, 3-D-glucan production by Aureobasidium pullulans. Int. J. Biol. Macromol. 2020, 164, 140–148. [Google Scholar] [CrossRef]
- Kejžar, A.; Grötli, M.; Tamás, M.J.; Plemenitaš, A.; Lenassi, M. HwHog1 kinase activity is crucial for survival of Hortaea werneckii in extremely hyperosmolar environments. Fungal Genet. Biol. 2015, 74, 45–58. [Google Scholar] [CrossRef]
- Yang, G.; Liu, G.-L.; Wang, S.-J.; Chi, Z.-M.; Chi, Z. Pullulan biosynthesis in yeast-like fungal cells is regulated by the transcriptional activator Msn2 and cAMP-PKA signaling pathway. Int. J. Biol. Macromol. 2020, 157, 591–603. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Contents/Functions | Relative Gene Expressions of Strain ZH27 (%) | Changes in Gene Expressions under the OC | |
|---|---|---|---|---|
| IC | OC | |||
| GluT1 | Glucose transporter | 100 | 70.9 ± 14.3 * | Downregulation |
| GluT2 | Glucose transporter | 100 | 1386.1 ± 485.8 ** | Upregulation |
| GluT3 | Glucose transporter | 100 | 69.5 ± 3.3 ** | Downregulation |
| GluT4 | Glucose transporter | 100 | 857.1 ± 126 ** | Upregulation |
| GluT5 | Glucose transporter | 100 | 2894.9 ± 750.2 ** | Upregulation |
| SUC2 | Sucrose-proton symporter 2 | 100 | 1432.9 ± 364.1 ** | Upregulation |
| GluK | Glucose kinase | 100 | 1844.6 ± 499.4 ** | Upregulation |
| PGM | Phosphoglucose mutase | 100 | 118.6 ± 20.3 | No significance |
| UGP | UDPG-pyrophosphorylase | 100 | 215.4 ± 29.9 ** | Upregulation |
| UGT1 | UDP-glucosyltranferase | 100 | 4742.8 ± 508.5 ** | Upregulation |
| PUL1 | Pullulan synthetase | 100 | 438.9 ± 70.7 ** | Upregulation |
| Ags2 | Alpha-glucan synthase 2 | 100 | 367.5 ± 77.4 ** | Upregulation |
| Msn2 | Stress-responsive transcriptional activator | 100 | 228.6 ± 13.5 ** | Upregulation |
| Apo | Glycolipid transfer protein | 100 | 153.5 ± 32.4 * | Upregulation |
| Gltp | Glycolipid transfer protein | 100 | 377.7 ± 98.6 ** | Upregulation |
| CreA | Regulation of gene transcription | 100 | 222.9 ± 22 ** | Upregulation |
| α-amylase gene | Hydrolysis of α-1,4-linkage | 100 | 3938.8 ± 200 ** | Upregulation |
| Glucoamylase gene | Hydrolysis of α-1,4- and α-1,6-linkages | 100 | 2040.8 ± 281.3 ** | Upregulation |
| Pullulanase gene | Hydrolysis of α-1,6-linkage | 100 | 1262.1 ± 174 ** | Upregulation |
| TPS1 | α-trehalose-6-phosphate synthase | 100 | 496.4 ± 50.1 ** | Upregulation |
| TPP | Trehalose-6-phosphate phosphatase | 100 | 278.7 ± 42.5 ** | Upregulation |
| GPP | Glycerol-3-phosphatase | 100 | 1341.7 ± 278.3 ** | Upregulation |
| GPD | Glycerol-3-phosphate dehydrogenase | 100 | 376.3 ± 56.4 ** | Upregulation |
| Hog1 | Hyper-osmotic glycerol | 100 | 256.7 ± 33.7 ** | Upregulation |
| Sho1 | High osmolarity signaling protein | 100 | 161.7 ± 4.0 ** | Upregulation |
| FF1 | Fructoslytranserase | 100 | 373.4 ± 174.8 ** | Upregulation |
| FF2 | Fructoslytranserase | 100 | 231.8 ± 56.7 ** | Upregulation |
| FF3 | Fructoslytranserase | 100 | 215.2 ± 37.9 ** | Upregulation |
| FTR1 | Transcriptional factor of fructoslytranserase genes | 100 | 333.1 ± 86.7 ** | Upregulation |
| PFK26 | Phosphofructo-2-kinase | 100 | 257.8 ± 25.5 ** | Upregulation |
| FBA1 | Fructose-bisphosphate aldolase1 | 100 | 524.7 ± 60.5 ** | Upregulation |
| G3P | Glyceraldehyde-3-phosphate dehydrogenase | 100 | 523 ± 102.6 ** | Upregulation |
| PGK | Phosphoglycerate kinase | 100 | 484.1 ± 18.7 ** | Upregulation |
| PYK | Pyruvate kinase | 100 | 203.6 ± 28.9 ** | Upregulation |
| PYD | Pyruvate decarboxylase | 100 | 245.1 ± 64 ** | Upregulation |
| AreA | Transcriptional activator of nitrogen | 100 | 135.7 ± 20 * | Upregulation |
| AreB | Transcriptional repressor of nitrogen | 100 | 85.3 ± 11 * | Downregulation |
| Strains | Fermentation | Production g/L (g/L/h) | Yield (g/g) | References | |
|---|---|---|---|---|---|
| Carbon | Mode and Conditions | ||||
| A. pullulans MG271838 | Sucrose | Batch, 28 °C, 200 rpm, 168 h | 37.6 (0.22) | 0.63 | [2] |
| A. pullulans H31 | Sucrose | Batch, 28 °C, 200 rpm, 96 h | 51.4 (0.54) | 0.93 | [3] |
| A. melanogenum 13-2 | Sucrose | Batch, 28 °C, 250 rpm, 6.5 L/min, 120 h | 78.1 (0.65) | 0.56 | [8] |
| A. melanogenum A4 | Maltose | Fed-batch, 30 °C, 180 rpm, ~3 L/min, 120 h | 122.3 (1.02) | 0.40 | [10] |
| A. melanogenum TN1-2 | Sucrose | Batch, 28 °C, 250 rpm, 6.5 L/min, 132 h | 114 (0.86) | 0.81 | [13] |
| A. melanogenum TN3-1 | Glucose | Batch, 28 °C, 250 rpm, 6.5 L/min, 132 h | 110.3 (0.84) | 0.79 | [18] |
| A. Pullulans M233-20 a | Glucose | Batch, 28 °C, 500 rpm, 20 L/min, 144 h | 162.3 (1.13) | 0.82 | [14] |
| A. pullulans LB83 | SBH | Batch, 25.3 °C, 232 rpm, 96 h | 25.2 (0.28) | 0.48 | [36] |
| A. melanogenum V6 a/F44 a | Sucrose | Batch, 28 °C, 180 rpm, 144 h | 102.3 (0.7)/ 101.4 (0.7) | 0.89/ 0.88 | [38] |
| A. melanogenum AMY-PKS-11 a | Glucose | Batch, 28 °C, 300 rpm, 6.5 L/min, 120 h | 103.5 (0.86) | 0.75 | [39] |
| A. melanogenum DG41 a | Glucose | Batch, 28 °C, 205 rpm, 8.0 L/min, 132 h | 64.9 (0.49) | 0.55 | [25] |
| A. Pullulans BL06 ΔPMAs a | Sucrose | Fed-batch, 28 °C, 500 rpm, 120 h | 140.2 (1.17) | <0.5 | [11] |
| A. melanogenum ZH27 | Sucrose | Batch, 28 °C, 200–220 rpm, 4.5–5.5 L/min, 132 h | 115.4 (0.87) | 0.77 | This study |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.-Q.; Lin, J.; Zhou, Q.-Z.; Peng, J.; Zhang, Q.; Wang, J.-H. Hyper-Production of Pullulan by a Novel Fungus of Aureobasidium melanogenum ZH27 through Batch Fermentation. Int. J. Mol. Sci. 2024, 25, 319. https://doi.org/10.3390/ijms25010319
Wang Q-Q, Lin J, Zhou Q-Z, Peng J, Zhang Q, Wang J-H. Hyper-Production of Pullulan by a Novel Fungus of Aureobasidium melanogenum ZH27 through Batch Fermentation. International Journal of Molecular Sciences. 2024; 25(1):319. https://doi.org/10.3390/ijms25010319
Chicago/Turabian StyleWang, Qin-Qing, Jia Lin, Qian-Zhi Zhou, Juan Peng, Qi Zhang, and Jiang-Hai Wang. 2024. "Hyper-Production of Pullulan by a Novel Fungus of Aureobasidium melanogenum ZH27 through Batch Fermentation" International Journal of Molecular Sciences 25, no. 1: 319. https://doi.org/10.3390/ijms25010319
APA StyleWang, Q.-Q., Lin, J., Zhou, Q.-Z., Peng, J., Zhang, Q., & Wang, J.-H. (2024). Hyper-Production of Pullulan by a Novel Fungus of Aureobasidium melanogenum ZH27 through Batch Fermentation. International Journal of Molecular Sciences, 25(1), 319. https://doi.org/10.3390/ijms25010319