
Calcium Signaling and the Response to Heat Shock in Crop Plants


Abstract
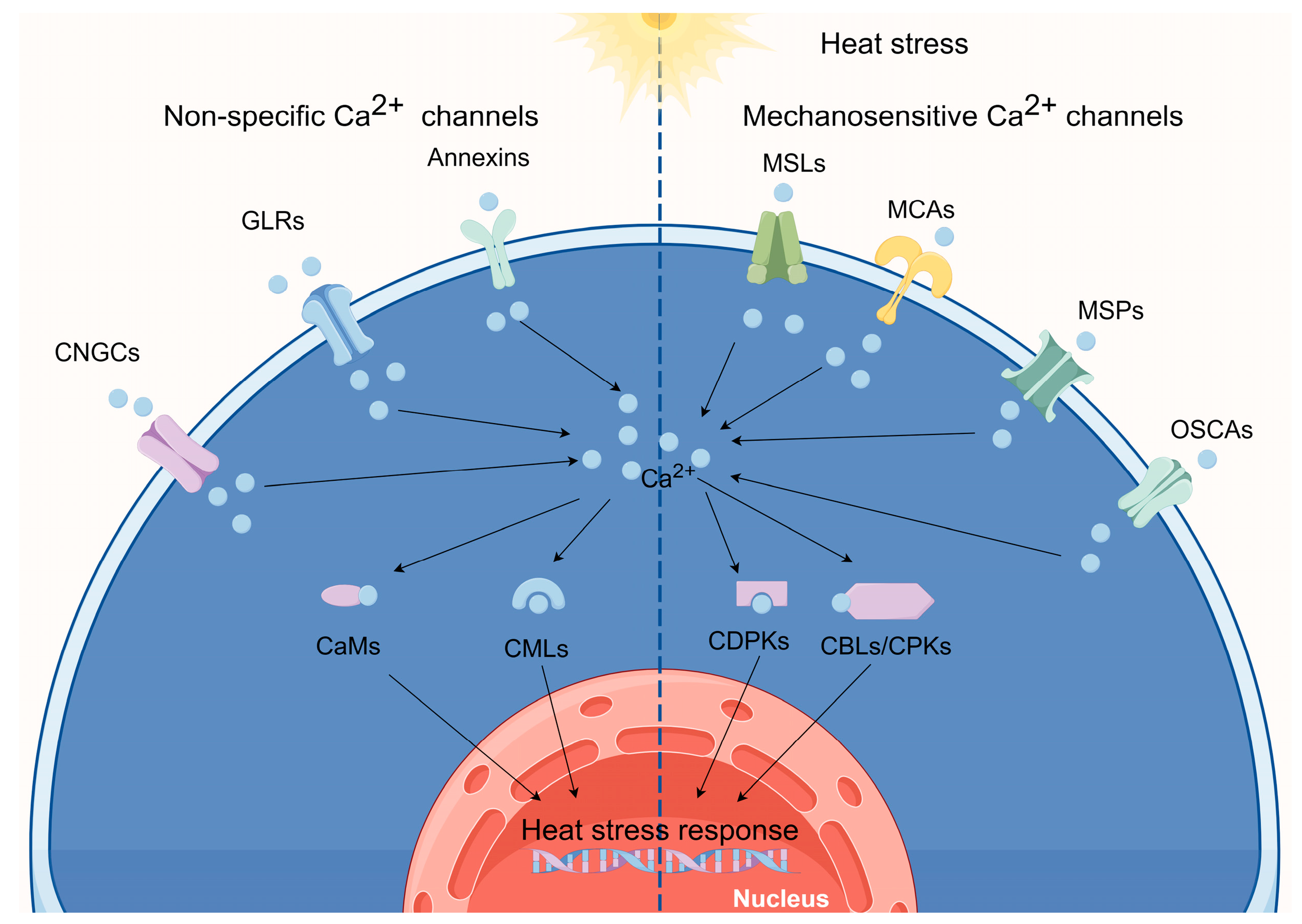
:1. Ca2+ Signaling and Plant Thermotolerance
2. Ca2+-Permeable Channels Perceive Elevated Temperatures
2.1. Heat Sensing via CNGCs
2.2. Heat Sensing via GLRs
2.3. Heat Sensing via Annexins
2.4. Heat Sensing via OSCAs
2.5. The Functions of MSLs, MCAs, and MSPs in Plants following HT Treatment
3. Ca2+-Binding Protein Involvement in the HSR
3.1. CaMs in HS Signaling
3.2. CMLs in HS Signaling
3.3. CDPKs in HS Signaling
3.4. CBLs and CIPKs in HS Signaling
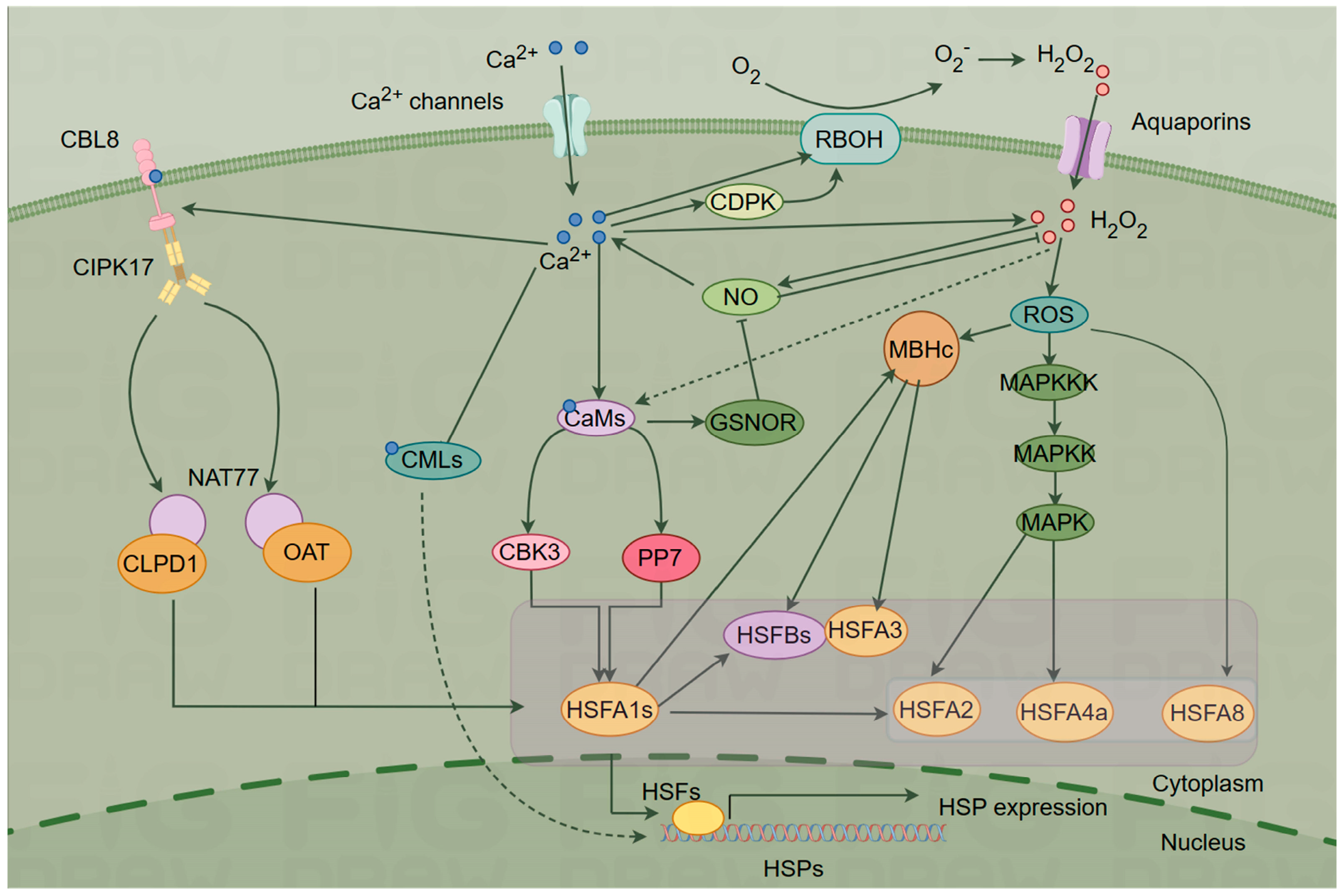
4. Ca2+ Signaling Networks Mediate the Plant HSR
4.1. ROS-Mediated Signaling
4.2. NO Signaling
4.3. HSF–HSP Signaling
5. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fitter, A.H.; Fitter, R.S.R. Rapid changes in flowering time in British plants. Science 2002, 296, 1689–1691. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Liu, B.; Piao, S.; Wang, X.; Lobell, D.B.; Huang, Y.; Huang, M.; Yao, Y.; Bassu, S.; Ciais, P.; et al. Temperature increase reduces global yields of major crops in four independent estimates. Proc. Natl. Acad. Sci. USA 2017, 114, 9326–9331. [Google Scholar] [CrossRef] [PubMed]
- Weigand, C.; Kim, S.H.; Brown, E.; Medina, E.; Mares, M.; Miller, G.; Harper, J.F.; Choi, W.G. A ratiometric calcium reporter CGf reveals calcium dynamics both in the single cell and whole plant levels under heat stress. Front. Plant Sci. 2021, 12, 777975. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Gao, K.; Ren, H.; Tang, W. Molecular mechanisms governing plant responses to high temperatures. J. Integr. Plant Biol. 2018, 60, 757–779. [Google Scholar] [CrossRef] [PubMed]
- Dodd, A.N.; Kudla, J.; Sanders, D. The language of calcium signaling. Annu. Rev. Plant Biol. 2010, 61, 593–620. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.-T.; Li, B.; Shang, Z.-L.; Li, X.-Z.; Mu, R.-L.; Sun, D.-Y.; Zhou, R.-G. Calmodulin is involved in heat shock signal transduction in wheat. Plant Physiol. 2003, 132, 1186–1195. [Google Scholar] [CrossRef]
- Kudla, J.; Batistic, O.; Hashimoto, K. Calcium signals: The lead currency of plant information processing. Plant Cell 2010, 22, 541–563. [Google Scholar] [CrossRef] [PubMed]
- Batistič, O.; Kudla, J. Analysis of calcium signaling pathways in plants. Biochim. Biophys. Acta 2012, 1820, 1283–1293. [Google Scholar] [CrossRef]
- Pirayesh, N.; Giridhar, M.; Ben Khedher, A.; Vothknecht, U.C.; Chigri, F. Organellar calcium signaling in plants: An update. Biochim. Biophys. Acta Mol. Cell Res. 2021, 1868, 118948. [Google Scholar] [CrossRef]
- Swarbreck, S.M.; Colaço, R.; Davies, J.M. Plant calcium-permeable channels. Plant Physiol. 2013, 163, 514–522. [Google Scholar] [CrossRef]
- Jammes, F.; Hu, H.C.; Villiers, F.; Bouten, R.; Kwak, J.M. Calcium-permeable channels in plant cells. FEBS J. 2011, 278, 4262–4276. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, B.; Sherman, T.; Fromm, H. Cyclic nucleotide-gated channels in plants. FEBS Lett. 2007, 581, 2237–2246. [Google Scholar] [CrossRef] [PubMed]
- DeFalco, T.A.; Marshall, C.B.; Munro, K.; Kang, H.G.; Moeder, W.; Ikura, M.; Snedden, W.A.; Yoshioka, K. Multiple calmodulin-binding sites positively and negatively regulate Arabidopsis cyclic nucleotide-gated channel 12. Plant Cell 2016, 28, 1738–1751. [Google Scholar] [CrossRef] [PubMed]
- Fischer, C.; Kugler, A.; Hoth, S.; Dietrich, P. An IQ domain mediates the interaction with calmodulin in a plant cyclic nucleotide-gated channel. Plant Cell Physiol. 2013, 54, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Schuurink, R.C.; Shartzer, S.F.; Fath, A.; Jones, R.L. Characterization of a calmodulin-binding transporter from the plasma membrane of barley aleurone. Proc. Natl. Acad. Sci. USA 1998, 95, 1944–1949. [Google Scholar] [CrossRef]
- Mäser, P.; Thomine, S.; Schroeder, J.I.; Ward, J.M.; Hirschi, K.; Sze, H.; Talke, I.N.; Amtmann, A.; Maathuis, F.J.; Sanders, D.; et al. Phylogenetic relationships within cation transporter families of Arabidopsis. Plant Physiol. 2001, 126, 1646–1667. [Google Scholar] [CrossRef]
- Bridges, D.; Fraser, M.E.; Moorhead, G.B. Cyclic nucleotide binding proteins in the Arabidopsis thaliana and Oryza sativa genomes. BMC Bioinform. 2005, 6, 6. [Google Scholar] [CrossRef]
- Zhou, L.; Lan, W.; Jiang, Y.; Fang, W.; Luan, S. A calcium-dependent protein kinase interacts with and activates a calcium channel to regulate pollen tube growth. Mol. Plant 2014, 7, 369–376. [Google Scholar] [CrossRef]
- Zhang, S.; Pan, Y.; Tian, W.; Dong, M.; Zhu, H.; Luan, S.; Li, L. Arabidopsis CNGC14 mediates calcium influx required for tipgrowth in root hairs. Mol. Plant 2017, 10, 1004–1006. [Google Scholar] [CrossRef]
- Chiasson, D.M.; Haage, K.; Sollweck, K.; Brachmann, A.; Dietrich, P.; Parniske, M. A quantitative hypermorphic CNGC allele confers ectopic calcium flux and impairs cellular development. eLife 2017, 6, e25012. [Google Scholar] [CrossRef]
- Wang, Y.; Kang, Y.; Ma, C.; Miao, R.; Wu, C.; Long, Y.; Ge, T.; Wu, Z.; Hou, X.; Zhang, J.; et al. CNGC2 is a Ca2+ influx channel that prevents accumulation of apoplastic Ca2+ in the leaf. Plant Physiol. 2017, 173, 1342–1354. [Google Scholar] [CrossRef] [PubMed]
- Finka, A.; Cuendet, A.F.; Maathuis, F.J.; Saidi, Y.; Goloubinoff, P. Plasma membrane cyclic nucleotide gated calcium channels control land plant thermal sensing and acquired thermotolerance. Plant Cell 2012, 24, 3333–3348. [Google Scholar] [CrossRef] [PubMed]
- Katano, K.; Kataoka, R.; Fujii, M.; Suzuki, N. Differences between seedlings and flowers in anti-ROS based heat responses of Arabidopsis plants deficient in cyclic nucleotide gated channel 2. Plant Physiol. Biochem. 2018, 123, 288–296. [Google Scholar] [CrossRef]
- Thoen, M.P.; Davila Olivas, N.H.; Kloth, K.J.; Coolen, S.; Huang, P.P.; Aarts, M.G.; Bac-Molenaar, J.A.; Bakker, J.; Bouwmeester, H.J.; Broekgaarden, C.; et al. Genetic architecture of plant stress resistance: Multi-trait genome-wide association mapping. New Phytol. 2017, 213, 1346–1362. [Google Scholar] [CrossRef] [PubMed]
- Gao, F.; Han, X.; Wu, J.; Zheng, S.; Shang, Z.; Sun, D.; Zhou, R.; Li, B. A heat-activated calcium-permeable channel—Arabidopsis cyclic nucleotide-gated ion channel 6—Is involved in heat shock responses. Plant J. 2012, 70, 1056–1069. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Zhang, J.; Ai, L.; Wu, D.; Li, B.; Zhang, L.; Zhao, L. Cyclic nucleotide-gated ion channel 6 mediates thermotolerance in Arabidopsis seedlings by regulating hydrogen peroxide production via cytosolic calcium ions. Front. Plant Sci. 2021, 12, 708672. [Google Scholar] [CrossRef]
- Peng, X.; Zhang, X.; Li, B.; Zhao, L. Cyclic nucleotide-gated ion channel 6 mediates thermotolerance in Arabidopsis seedlings by regulating nitric oxide production via cytosolic calcium ions. BMC Plant Biol. 2019, 19, 368. [Google Scholar] [CrossRef]
- Cui, Y.; Lu, S.; Li, Z.; Cheng, J.; Hu, P.; Zhu, T.; Wang, X.; Jin, M.; Wang, X.; Li, L.; et al. Cyclic nucleotide-gated ion channels 14 and 16 promote tolerance to heat and chilling in rice. Plant Physiol. 2020, 183, 1794–1808. [Google Scholar] [CrossRef]
- Nawaz, Z.; Kakar, K.U.; Saand, M.A.; Shu, Q.Y. Cyclic nucleotide-gated ion channel gene family in rice, identification, characterization and experimental analysis of expression response to plant hormones, biotic and abiotic stresses. BMC Genom. 2014, 15, 853. [Google Scholar] [CrossRef]
- Mayer, M.L. Structural biology of glutamate receptor ion channel complexes. Curr. Opin. Struct. Biol. 2016, 41, 119–127. [Google Scholar] [CrossRef]
- Lam, H.M.; Chiu, J.; Hsieh, M.H.; Meisel, L.; Oliveira, I.C.; Shin, M.; Coruzzi, G. Glutamate-receptor genes in plants. Nature 1998, 396, 125–126. [Google Scholar] [CrossRef] [PubMed]
- Lacombe, B.; Becker, D.; Hedrich, R.; DeSalle, R.; Hollmann, M.; Kwak, J.M.; Schroeder, J.I.; Le Novère, N.; Nam, H.G.; Spalding, E.P.; et al. The identity of plant glutamate receptors. Science 2001, 292, 1486–1487. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Zhu, S.; Song, X.; Shen, Y.; Chen, H.; Yu, J.; Yi, K.; Liu, Y.; Karplus, V.J.; Wu, P.; et al. A rice glutamate receptor-like gene is critical for the division and survival of individual cells in the root apical meristem. Plant Cell 2006, 18, 340–349. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, P.; Anschütz, U.; Kugler, A.; Becker, D. Physiology and biophysics of plant ligand-gated ion channels. Plant Biol. 2010, 12 (Suppl. 1), 80–93. [Google Scholar] [CrossRef] [PubMed]
- Roy, B.C.; Mukherjee, A. Computational analysis of the glutamate receptor gene family of Arabidopsis thaliana. J. Biomol. Struct. Dyn. 2017, 35, 2454–2474. [Google Scholar] [CrossRef] [PubMed]
- Ward, J.M.; Mäser, P.; Schroeder, J.I. Plant ion channels: Gene families, physiology, and functional genomics analyses. Annu. Rev. Physiol. 2009, 71, 59–82. [Google Scholar] [CrossRef] [PubMed]
- Davenport, R. Glutamate receptors in plants. Ann. Bot. 2002, 90, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Salvador-Recatalà, V. New roles for the glutamate receptor-link 3.3, 3.5, and 3.6 genes as on/off switches of wound-induced systemic electrical signals. Plant Signal. Behav. 2016, 11, e1161879. [Google Scholar] [CrossRef]
- Green, M.N.; Gangwar, S.P.; Michard, E.; Simon, A.A.; Portes, M.T.; Barbosa-Caro, J.; Wudick, M.M.; Lizzio, M.A.; Klykov, O.; Yelshanskaya, M.V.; et al. Structure of the Arabidopsis thaliana glutamate receptor-like channel GLR3.4. Mol. Cell 2021, 81, 3216–3226. [Google Scholar] [CrossRef]
- Kong, D.; Hu, H.C.; Okuma, E.; Lee, Y.; Lee, H.S.; Munemasa, S.; Cho, D.; Ju, C.; Pedoeim, L.; Rodriguez, B.; et al. L-Met activates Arabidopsis GLR Ca2+ channels upstream of ROS production and regulates stomatal movement. Cell Rep. 2016, 17, 2553–2561. [Google Scholar] [CrossRef]
- Xue, N.; Zhan, C.; Song, J.; Li, Y.; Zhang, J.; Qi, J.; Wu, J. The glutamate receptor-like 3.3 and 3.6 mediate systemic resistance to insect herbivores in Arabidopsis. J. Exp. Bot. 2022, 73, 7611–7627. [Google Scholar] [CrossRef] [PubMed]
- Zandalinas, S.I.; Fichman, Y.; Mittler, R. Vascular bundles mediate systemic reactive oxygen signaling during light stress. Plant Cell 2020, 32, 3425–3435. [Google Scholar] [CrossRef]
- Manzoor, H.; Kelloniemi, J.; Chiltz, A.; Wendehenne, D.; Pugin, A.; Poinssot, B.; Garcia-Brugger, A. Involvement of the glutamate receptor AtGLR3.3 in plant defense signaling and resistance to Hyaloperonospora arabidopsidis. Plant J. 2013, 76, 466–480. [Google Scholar] [CrossRef]
- Yoshida, R.; Mori, I.C.; Kamizono, N.; Shichiri, Y.; Shimatani, T.; Miyata, F.; Honda, K.; Iwai, S. Glutamate functions in stomatal closure in Arabidopsis and fava bean. J. Plant Res. 2016, 129, 39–49. [Google Scholar] [CrossRef]
- Zhou, H.; Duan, H.; Liu, Y.; Sun, X.; Zhao, J.; Lin, H. Patellin protein family functions in plant development and stress response. J. Plant Physiol. 2019, 234–235, 94–97. [Google Scholar] [CrossRef]
- Li, Z.G.; Ye, X.Y.; Qiu, X.M. Glutamate signaling enhances the heat tolerance of maize seedlings by plant glutamate receptor-like channels-mediated calcium signaling. Protoplasma 2019, 256, 1165–1169. [Google Scholar] [CrossRef]
- Mortimer, J.C.; Laohavisit, A.; Macpherson, N.; Webb, A.; Brownlee, C.; Battey, N.H.; Davies, J.M. Annexins: Multifunctional components of growth and adaptation. J. Exp. Bot. 2008, 59, 533–544. [Google Scholar] [CrossRef]
- Davies, J.M. Annexin-mediated calcium signalling in plants. Plants 2014, 3, 128–140. [Google Scholar] [CrossRef]
- Gorecka, K.M.; Konopka-Postupolska, D.; Hennig, J.; Buchet, R.; Pikula, S. Peroxidase activity of annexin 1 from Arabidopsis thaliana. Biochem. Biophys. Res. Commun. 2005, 336, 868–875. [Google Scholar] [CrossRef]
- Gerke, V.; Creutz, C.E.; Moss, S.E. Annexins: Linking Ca2+ signalling to membrane dynamics. Nat. Rev. Mol. Cell Biol. 2005, 6, 449–461. [Google Scholar] [CrossRef]
- Clark, G.B.; Sessions, A.; Eastburn, D.J.; Roux, S.J. Differential expression of members of the annexin multigene family in Arabidopsis. Plant Physiol. 2001, 126, 1072–1084. [Google Scholar] [CrossRef] [PubMed]
- Jami, S.K.; Clark, G.B.; Ayele, B.T.; Roux, S.J.; Kirti, P.B. Identification and characterization of annexin gene family in rice. Plant Cell Rep. 2012, 31, 813–825. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Tang, Y.; Gao, S.; Su, S.; Hong, L.; Wang, W.; Fang, Z.; Li, X.; Ma, J.; Quan, W.; et al. Comprehensive analyses of the annexin gene family in wheat. BMC Genom. 2016, 17, 415. [Google Scholar] [CrossRef] [PubMed]
- Gerke, V.; Moss, S.E. Annexins: From structure to function. Physiol. Rev. 2002, 82, 331–371. [Google Scholar] [CrossRef] [PubMed]
- Saad, R.B.; Ben Romdhane, W.; Ben Hsouna, A.; Mihoubi, W.; Harbaoui, M.; Brini, F. Insights into plant annexins function in abiotic and biotic stress tolerance. Plant Signal. Behav. 2020, 15, 1699264. [Google Scholar] [CrossRef] [PubMed]
- Laohavisit, A.; Davies, J.M. Annexins. New Phytol. 2011, 189, 40–53. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Wang, Y.; Bian, Y.; Ren, Y.; Xu, X.; Zhou, F.; Ding, H. A critical review on plant annexin: Structure, function, and mechanism. Plant Physiol. Biochem. 2022, 190, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Chu, P.; Chen, H.; Zhou, Y.; Li, Y.; Ding, Y.; Jiang, L.; Tsang, E.W.; Wu, K.; Huang, S. Proteomic and functional analyses of Nelumbo nucifera annexins involved in seed thermotolerance and germination vigor. Planta 2012, 235, 1271–1288. [Google Scholar] [CrossRef]
- Wei, J.; Shen, Y.; Zhao, H.; Liu, X.; Jia, Y.; Yu, X.; Ma, H. GmANN, a glutathione S-transferase-interacting annexin, is involved in high temperature and humidity tolerance and seed vigor formation in transgenic Arabidopsis. Plant Cell 2019, 138, 583–595. [Google Scholar] [CrossRef]
- Qiao, B.; Zhang, Q.; Liu, D.; Wang, H.; Yin, J.; Wang, R.; He, M.; Cui, M.; Shang, Z.; Wang, D.; et al. A calcium-binding protein, rice annexin OsANN1, enhances heat stress tolerance by modulating the production of H2O2. J. Exp. Bot. 2015, 66, 5853–5866. [Google Scholar] [CrossRef]
- Shen, F.; Ying, J.; Xu, L.; Sun, X.; Wang, J.; Wang, Y.; Mei, Y.; Zhu, Y.; Liu, L. Characterization of Annexin gene family and functional analysis of RsANN1a involved in heat tolerance in radish (Raphanus sativus L.). Physiol. Mol. Biol. Plants 2021, 27, 2027–2041. [Google Scholar] [CrossRef] [PubMed]
- Yuan, F.; Yang, H.; Xue, Y.; Kong, D.; Ye, R.; Li, C.; Zhang, J.; Theprungsirikul, L.; Shrift, T.; Krichilsky, B.; et al. OSCA1 mediates osmotic-stress-evoked Ca2+ increases vital for osmosensing in Arabidopsis. Nature 2014, 514, 367–371. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yuan, F.; Wen, Z.; Li, Y.; Wang, F.; Zhu, T.; Zhuo, W.; Jin, X.; Wang, Y.; Zhao, H.; et al. Genome-wide survey and expression analysis of the OSCA gene family in rice. BMC Plant Biol. 2015, 15, 261. [Google Scholar] [CrossRef] [PubMed]
- Ding, S.; Feng, X.; Du, H.; Wang, H. Genome-wide analysis of maize OSCA family members and their involvement in drought stress. PeerJ 2019, 7, e6765. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Xu, Y.; Yang, F.; Magwanga, R.O.; Cai, X.; Wang, X.; Wang, Y.; Hou, Y.; Wang, K.; Liu, F.; et al. Genome-wide identification of OSCA gene family and their potential function in the regulation of dehydration and salt stress in Gossypium hirsutum. J. Cotton Res. 2019, 2, 11. [Google Scholar] [CrossRef]
- Li, Y.; Zhang, Y.; Li, B.; Hou, L.; Yu, J.; Jia, C.; Wang, Z.; Chen, S.; Zhang, M.; Qin, J.; et al. Preliminary expression analysis of the OSCA gene family in Maize and their involvement in temperature stress. Int. J. Mol. Sci. 2022, 23, 13658. [Google Scholar] [CrossRef]
- Haswell, E.S.; Meyerowitz, E.M. MscS-like proteins control plastid size and shape in Arabidopsis thaliana. Curr. Biol. 2006, 16, 1–11. [Google Scholar] [CrossRef]
- Hamilton, E.S.; Haswell, E.S. The tension-sensitive ion transport activity of MSL8 is critical for its function in pollen hydration and germination. Plant Cell Physiol. 2017, 58, 1222–1237. [Google Scholar] [CrossRef]
- Peyronnet, R.; Haswell, E.S.; Barbier-Brygoo, H.; Frachisse, J.M. AtMSL9 and AtMSL10: Sensors of plasma membrane tension in Arabidopsis roots. Plant Signal. Behav. 2008, 3, 726–729. [Google Scholar] [CrossRef]
- Li, J.; Zhang, M.; Sun, J.; Mao, X.; Wang, J.; Wang, J.; Liu, H.; Zheng, H.; Zhen, Z.; Zhao, H.; et al. Genome-Wide characterization and identification of trihelix transcription factor and expression profiling in response to abiotic stresses in rice (Oryza sativa L.). Int. J. Mol. Sci. 2019, 20, 251. [Google Scholar] [CrossRef]
- Nakagawa, Y.; Katagiri, T.; Shinozaki, K.; Qi, Z.; Tatsumi, H.; Furuichi, T.; Kishigami, A.; Sokabe, M.; Kojima, I.; Sato, S.; et al. Arabidopsis plasma membrane protein crucial for Ca2+ influx and touch sensing in roots. Proc. Natl. Acad. Sci. USA 2007, 104, 3639–3644. [Google Scholar] [CrossRef] [PubMed]
- Kurusu, T.; Yamanaka, T.; Nakano, M.; Takiguchi, A.; Ogasawara, Y.; Hayashi, T.; Iida, K.; Hanamata, S.; Shinozaki, K.; Iida, H.; et al. Involvement of the putative Ca2+-permeable mechanosensitive channels, NtMCA1 and NtMCA2, in Ca2+ uptake, Ca2+-dependent cell proliferation and mechanical stress-induced gene expression in tobacco (Nicotiana tabacum) BY-2 cells. J. Plant Res. 2012, 125, 555–568. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, T.; Nakagawa, Y.; Mori, K.; Nakano, M.; Imamura, T.; Kataoka, H.; Terashima, A.; Iida, K.; Kojima, I.; Katagiri, T.; et al. MCA1 and MCA2 that mediate Ca2+ uptake have distinct and overlapping roles in Arabidopsis. Plant Physiol. 2010, 152, 1284–1296. [Google Scholar] [CrossRef] [PubMed]
- Mori, K.; Renhu, N.; Naito, M.; Nakamura, A.; Shiba, H.; Yamamoto, T.; Suzaki, T.; Iida, H.; Miura, K. Ca2+-permeable mechanosensitive channels MCA1 and MCA2 mediate cold-induced cytosolic Ca2+ increase and cold tolerance in Arabidopsis. Sci. Rep. 2018, 8, 550. [Google Scholar] [CrossRef] [PubMed]
- Coste, B.; Mathur, J.; Schmidt, M.; Earley, T.J.; Ranade, S.; Petrus, M.J.; Dubin, A.E.; Patapoutian, A. Piezo1 and Piezo2 are essential components of distinct mechanically activated cation channels. Science 2010, 330, 55–60. [Google Scholar] [CrossRef] [PubMed]
- Kurusu, T.; Kuchitsu, K.; Nakano, M.; Nakayama, Y.; Iida, H. Plant mechanosensing and Ca2+ transport. Trends Plant Sci. 2013, 18, 227–233. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Tong, X.; Liu, S.Y.; Chai, L.X.; Zhu, F.F.; Zhang, X.P.; Zou, J.Z.; Wang, X.B. Genetic analysis of a Piezo-like protein suppressing systemic movement of plant viruses in Arabidopsis thaliana. Sci. Rep. 2019, 9, 3187. [Google Scholar] [CrossRef]
- Knight, H. Calcium signaling during abiotic stress in plants. Int. Rev. Cytol. 2000, 195, 269–324. [Google Scholar] [CrossRef]
- Mohanta, T.K.; Yadav, D.; Khan, A.L.; Hashem, A.; Abd Allah, E.F.; Al-Harrasi, A. Molecular players of EF-hand containing calcium signaling event in plants. Int. J. Mol. Sci. 2019, 20, 1476. [Google Scholar] [CrossRef]
- Braam, J.; Davis, R.W. Rain-, wind-, and touch-induced expression of calmodulin and calmodulin-related genes in Arabidopsis. Cell 1990, 60, 357–364. [Google Scholar] [CrossRef]
- Al-Quraan, N.A.; Locy, R.D.; Singh, N.K. Expression of calmodulin genes in wild type and calmodulin mutants of Arabidopsis thaliana under heat stress. Plant Physiol. Biochem. 2010, 48, 697–702. [Google Scholar] [CrossRef]
- Choi, M.S.; Kim, M.C.; Yoo, J.H.; Moon, B.C.; Koo, S.C.; Park, B.O.; Lee, J.H.; Koo, Y.D.; Han, H.J.; Lee, S.Y.; et al. Isolation of a calmodulin-binding transcription factor from rice (Oryza sativa L.). J. Biol. Chem. 2005, 280, 40820–40831. [Google Scholar] [CrossRef] [PubMed]
- Hardoim, P.R.; de Carvalho, T.L.; Ballesteros, H.G.; Bellieny-Rabelo, D.; Rojas, C.A.; Venancio, T.M.; Ferreira, P.C.; Hemerly, A.S. Genome-wide transcriptome profiling provides insights into the responses of maize (Zea mays L.) to diazotrophic bacteria. Plant Soil 2020, 451, 121–143. [Google Scholar] [CrossRef]
- Gifford, J.L.; Walsh, M.P.; Vogel, H.J. Structures and metal-ion-binding properties of the Ca2+-binding helix-loop-helix EF-hand motifs. Biochem. J. 2007, 405, 199–221. [Google Scholar] [CrossRef] [PubMed]
- Kundu, P.; Nehra, A.; Gill, R.; Tuteja, N.; Gill, S.S. Unraveling the importance of EF-hand-mediated calcium signaling in plants. S. Afr. J. Bot. 2022, 148, 615–633. [Google Scholar] [CrossRef]
- Niu, W.-T.; Han, X.-W.; Wei, S.-S.; Shang, Z.-L.; Wang, J.; Yang, D.-W.; Fan, X.; Gao, F.; Zheng, S.-Z.; Bai, J.-T.; et al. Arabidopsis cyclic nucleotide-gated channel 6 is negatively modulated by multiple calmodulin isoforms during heat shock. J. Exp. Bot. 2020, 71, 90–104. [Google Scholar] [CrossRef]
- Zhang, W.; Zhou, R.G.; Gao, Y.J.; Zheng, S.Z.; Xu, P.; Zhang, S.Q.; Sun, D.Y. Molecular and genetic evidence for the key role of AtCaM3 in heat-shock signal transduction in Arabidopsis. Plant Physiol. 2009, 149, 1773–1784. [Google Scholar] [CrossRef]
- Xuan, Y.; Zhou, S.; Wang, L.; Cheng, Y.; Zhao, L. Nitric oxide functions as a signal and acts upstream of AtCaM3 in thermotolerance in Arabidopsis seedlings. Plant Physiol. 2010, 153, 1895–1906. [Google Scholar] [CrossRef]
- Yu, B.; Yan, S.; Zhou, H.; Dong, R.; Lei, J.; Chen, C.; Cao, B. Overexpression of CsCaM3 improves high temperature tolerance in cucumber. Front. Plant Sci. 2018, 9, 797. [Google Scholar] [CrossRef]
- Wu, H.C.; Luo, D.L.; Vignols, F.; Jinn, T.L. Heat shock-induced biphasic Ca2+ signature and OsCaM1-1 nuclear localization mediate downstream signalling in acquisition of thermotolerance in rice (Oryza sativa L.). Plant Cell Environ. 2012, 35, 1543–1557. [Google Scholar] [CrossRef]
- McCormack, E.; Tsai, Y.C.; Braam, J. Handling calcium signaling: Arabidopsis CaMs and CMLs. Trends Plant Sci. 2005, 10, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Braam, J. Regulated expression of the calmodulin-related TCH genes in cultured Arabidopsis cells: Induction by calcium and heat shock. Proc. Natl. Acad. Sci. USA 1992, 89, 3213–3216. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.Y.; Rocha, P.S.; Wang, M.L.; Xu, M.L.; Cui, Y.C.; Li, L.Y.; Zhu, Y.X.; Xia, X. A novel rice calmodulin-like gene, OsMSR2, enhances drought and salt tolerance and increases ABA sensitivity in Arabidopsis. Planta 2011, 234, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Ding, H.; Qian, Y.; Fang, Y.; Ji, Y.; Sheng, J.; Ge, C. Characteristics of SlCML39, a tomato calmodulin-like gene, and its negative role in high temperature tolerance of Arabidopsis thaliana during germination and seedling growth. Int. J. Mol. Sci. 2021, 22, 11479. [Google Scholar] [CrossRef] [PubMed]
- Atif, R.M.; Shahid, L.; Waqas, M.; Ali, B.; Rashid, M.A.R.; Azeem, F.; Nawaz, M.A.; Wani, S.H.; Chung, G. Insights on calcium-dependent protein kinases (CPKs) signaling for abiotic stress tolerance in plants. Int. J. Mol. Sci. 2019, 20, 5298. [Google Scholar] [CrossRef] [PubMed]
- Yip Delormel, T.; Boudsocq, M. Properties and functions of calcium-dependent protein kinases and their relatives in Arabidopsis thaliana. New Phytol. 2019, 224, 585–604. [Google Scholar] [CrossRef]
- Cheng, S.H.; Willmann, M.R.; Chen, H.C.; Sheen, J. Calcium signaling through protein kinases. The Arabidopsis calcium-dependent protein kinase gene family. Plant Physiol. 2002, 129, 469–485. [Google Scholar] [CrossRef]
- Hu, Z.; Lv, X.; Xia, X.; Zhou, J.; Shi, K.; Yu, J.; Zhou, Y. Genome-wide identification and expression analysis of calcium-dependent protein kinase in tomato. Front. Plant Sci. 2016, 7, 469. [Google Scholar] [CrossRef]
- Kong, X.; Lv, W.; Jiang, S.; Zhang, D.; Cai, G.; Pan, J.; Li, D. Genome-wide identification and expression analysis of calcium-dependent protein kinase in maize. BMC Genom. 2013, 14, 433. [Google Scholar] [CrossRef]
- Veremeichik, G.N.; Shkryl, Y.N.; Gorpenchenko, T.Y.; Silantieva, S.A.; Avramenko, T.V.; Brodovskaya, E.V.; Bulgakov, V.P. Inactivation of the auto-inhibitory domain in Arabidopsis AtCPK1 leads to increased salt, cold and heat tolerance in the AtCPK1-transformed Rubia cordifolia L. cell cultures. Plant Physiol. Biochem. 2021, 159, 372–382. [Google Scholar] [CrossRef]
- Hu, Z.; Li, J.; Ding, S.; Cheng, F.; Li, X.; Jiang, Y.; Yu, J.; Foyer, C.H.; Shi, K. The protein kinase CPK28 phosphorylates ascorbate peroxidase and enhances thermotolerance in tomato. Plant Physiol. 2021, 186, 1302–1317. [Google Scholar] [CrossRef] [PubMed]
- Chang, W.J.; Su, H.S.; Li, W.J.; Zhang, Z.L. Expression profiling of a novel calcium-dependent protein kinase gene, LeCPK2, from tomato (Solanum lycopersicum) under heat and pathogen-related hormones. Biosci. Biotechnol. Biochem. 2009, 73, 2427–2431. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Du, H.; Wang, Y.; Wang, H.; Yang, S.; Li, C.; Chen, N.; Yang, H.; Zhang, Y.; Zhu, Y.; et al. The calcium-dependent protein kinase ZmCDPK7 functions in heat-stress tolerance in maize. J. Int. Plant Biol. 2021, 63, 510–527. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.-T.; Song, W. ZmCK3, a maize calcium-dependent protein kinase gene, endows tolerance to drought and heat stresses in transgenic Arabidopsis. J. Plant Biochem. 2014, 23, 249–256. [Google Scholar] [CrossRef]
- Wei, J.-T.; Hou, Z.-H.; Wang, Y.; Hao, J.-M.; Wang, J.; Wang, W.; Wang, W.; Wang, D.-M.; Xu, Z.-S.; Song, X.; et al. Foxtail millet SiCDPK7 gene enhances tolerance to extreme temperature stress in transgenic plants. Environ. Exp. Bot. 2023, 207, 105197. [Google Scholar] [CrossRef]
- Dubrovina, A.S.; Kiselev, K.V.; Khristenko, V.S.; Aleynova, O.A. The calcium-dependent protein kinase gene VaCPK29 is involved in grapevine responses to heat and osmotic stresses. Plant Growth Regul. 2017, 82, 79–89. [Google Scholar] [CrossRef]
- Zhang, K.; Han, Y.T.; Zhao, F.L.; Hu, Y.; Gao, Y.R.; Ma, Y.F.; Zheng, Y.; Wang, Y.J.; Wen, Y.Q. Genome-wide identification and expression analysis of the CDPK gene family in grape, Vitis spp. BMC Plant Biol. 2015, 15, 164. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Liu, W.Z.; Zhang, Y.; Deng, M.; Niu, F.; Yang, B.; Wang, X.; Wang, B.; Liang, W.; Deyholos, M.K.; et al. Identification, expression and interaction analyses of calcium-dependent protein kinase (CPK) genes in canola (Brassica napus L.). BMC Genom. 2014, 15, 211. [Google Scholar] [CrossRef]
- Wu, P.; Wang, W.; Duan, W.; Li, Y.; Hou, X. Comprehensive analysis of the CDPK-SnRK superfamily genes in Chinese cabbage and its evolutionary implications in plants. Front. Plant Sci. 2017, 8, 162. [Google Scholar] [CrossRef]
- Xu, X.; Liu, M.; Lu, L.; He, M.; Qu, W.; Xu, Q.; Qi, X.; Chen, X. Genome-wide analysis and expression of the calcium-dependent protein kinase gene family in cucumber. Mol. Genet. Genom. 2015, 290, 1403–1414. [Google Scholar] [CrossRef]
- Cai, H.; Cheng, J.; Yan, Y.; Xiao, Z.; Li, J.; Mou, S.; Qiu, A.; Lai, Y.; Guan, D.; He, S. Genome-wide identification and expression analysis of calcium-dependent protein kinase and its closely related kinase genes in Capsicum annuum. Front. Plant Sci. 2015, 6, 737. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Zhu, J.K. A calcium sensor homolog required for plant salt tolerance. Science 1998, 280, 1943–1945. [Google Scholar] [CrossRef] [PubMed]
- Luan, S. The CBL-CIPK network in plant calcium signaling. Trends Plant Sci. 2009, 14, 37–42. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, S.K.; Mahiwal, S.; Nambiar, D.M.; Pandey, G.K. CBL-CIPK module-mediated phosphoregulation: Facts and hypothesis. Biochem. J. 2020, 477, 853–871. [Google Scholar] [CrossRef] [PubMed]
- Gao, C.; Lu, S.; Zhou, R.; Wang, Z.; Li, Y.; Fang, H.; Wang, B.; Chen, M.; Cao, Y. The OsCBL8-OsCIPK17 module regulates seedling growth and confers resistance to heat and drought in rice. Int. J. Mol. Sci. 2022, 23, 12451. [Google Scholar] [CrossRef] [PubMed]
- Aslam, M.; Greaves, J.G.; Jakada, B.H.; Fakher, B.; Wang, X.; Qin, Y. AcCIPK5, a pineapple CBL-interacting protein kinase, confers salt, osmotic and cold stress tolerance in transgenic Arabidopsis. Plant Sci. 2022, 320, 111284. [Google Scholar] [CrossRef] [PubMed]
- He, H.; Garcia-Mata, C.; He, L.-F. Interaction between hydrogen sulfide and hormones in plant physiological responses. Plant Growth Regul. 2018, 87, 175–186. [Google Scholar] [CrossRef]
- Yang, F.; Dong, F.-S.; Hu, F.-H.; Liu, Y.-W.; Chai, J.-F.; Zhao, H.; Lv, M.-Y.; Zhou, S. Genome-wide identification and expression analysis of the calmodulin-binding transcription activator (CAMTA) gene family in wheat (Triticum aestivum L.). BMC Genet. 2020, 21, 105. [Google Scholar] [CrossRef]
- Kidokoro, S.; Konoura, I.; Soma, F.; Shinozaki, K.; Suzuki, T.; Tanokura, M.; Miyakawa, T.; Imaizumi, T.; Yamaguchi-Shinozaki, K. Clock-regulated coactivators selectively control gene expression in response to different temperature stress conditions in Arabidopsis. Proc. Natl. Acad. Sci. USA 2023, 120, e2216183120. [Google Scholar] [CrossRef]
- Li, Z.G.; Gong, M.; Xie, H.; Yang, L.; Li, J. Hydrogen sulfide donor sodium hydrosulfide-induced heat tolerance in tobacco (Nicotiana tabacum L.) suspension cultured cells and involvement of Ca2+ and calmodulin. Plant Sci. 2012, 185, 185–189. [Google Scholar] [CrossRef]
- Wang, L.; Wan, R.; Shi, Y.; Xue, S. Hydrogen sulfide activates S-type anion channel via OST1 and Ca2+ modules. Mol. Plant 2016, 9, 489–491. [Google Scholar] [CrossRef] [PubMed]
- Krebs, J.; Agellon, L.B.; Michalak, M. Ca2+ homeostasis and endoplasmic reticulum (ER) stress: An integrated view of calcium signaling. Biochem. Biophys. Res. Commun. 2015, 460, 114–121. [Google Scholar] [CrossRef]
- Carreras-Sureda, A.; Pihán, P.; Hetz, C. Calcium signaling at the endoplasmic reticulum: Fine-tuning stress responses. Cell Calcium 2018, 70, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Yao, Y.; He, R.J.; Xie, Q.L.; Zhao, X.H.; Deng, X.M.; He, J.B.; Song, L.; He, J.; Marchant, A.; Chen, X.Y.; et al. Ethylene response factor 74 (ERF74) plays an essential role in controlling a respiratory burst oxidase homolog D (RbohD)-dependent mechanism in response to different stresses in Arabidopsis. New Phytol. 2017, 213, 1667–1681. [Google Scholar] [CrossRef] [PubMed]
- Saxena, I.; Srikanth, S.; Chen, Z. Cross talk between H2O2 and interacting signal molecules under plant stress response. Front. Plant Sci. 2016, 7, 570. [Google Scholar] [CrossRef] [PubMed]
- Ogasawara, Y.; Kaya, H.; Hiraoka, G.; Yumoto, F.; Kimura, S.; Kadota, Y.; Hishinuma, H.; Senzaki, E.; Yamagoe, S.; Nagata, K.; et al. Synergistic activation of the Arabidopsis NADPH oxidase AtrbohD by Ca2+ and phosphorylation. J. Biol. Chem. 2008, 283, 8885–8892. [Google Scholar] [CrossRef] [PubMed]
- Kimura, S.; Kaya, H.; Kawarazaki, T.; Hiraoka, G.; Senzaki, E.; Michikawa, M.; Kuchitsu, K. Protein phosphorylation is a prerequisite for the Ca2+-dependent activation of Arabidopsis NADPH oxidases and may function as a trigger for the positive feedback regulation of Ca2+ and reactive oxygen species. Biochim. Biophys. Acta 2012, 1823, 398–405. [Google Scholar] [CrossRef]
- Bienert, G.P.; Møller, A.L.; Kristiansen, K.A.; Schulz, A.; Møller, I.M.; Schjoerring, J.K.; Jahn, T.P. Specific aquaporins facilitate the diffusion of hydrogen peroxide across membranes. J. Biol. Chem. 2007, 282, 1183–1192. [Google Scholar] [CrossRef]
- Jaimes-Miranda, F.; Chávez Montes, R.A. The plant MBF1 protein family: A bridge between stress and transcription. J. Exp. Bot. 2020, 71, 1782–1791. [Google Scholar] [CrossRef]
- Suzuki, N.; Miller, G.; Salazar, C.; Mondal, H.A.; Shulaev, E.; Cortes, D.F.; Shuman, J.L.; Luo, X.; Shah, J.; Schlauch, K.; et al. Temporal-spatial interaction between reactive oxygen species and abscisic acid regulates rapid systemic acclimation in plants. Plant Cell 2013, 25, 3553–3569. [Google Scholar] [CrossRef]
- Suzuki, N.; Sejima, H.; Tam, R.; Schlauch, K.; Mittler, R. Identification of the MBF1 heat-response regulon of Arabidopsis thaliana. Plant J. 2011, 66, 844–851. [Google Scholar] [CrossRef] [PubMed]
- Giesguth, M.; Sahm, A.; Simon, S.; Dietz, K.J. Redox-dependent translocation of the heat shock transcription factor AtHSFA8 from the cytosol to the nucleus in Arabidopsis thaliana. FEBS Lett. 2015, 589, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Miller, G.; Mittler, R. Could heat shock transcription factors function as hydrogen peroxide sensors in plants? Ann. Bot. 2006, 98, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Kovtun, Y.; Chiu, W.L.; Tena, G.; Sheen, J. Functional analysis of oxidative stress-activated mitogen-activated protein kinase cascade in plants. Proc. Natl. Acad. Sci. USA 2000, 97, 2940–2945. [Google Scholar] [CrossRef] [PubMed]
- Evrard, A.; Kumar, M.; Lecourieux, D.; Lucks, J.; von Koskull-Döring, P.; Hirt, H. Regulation of the heat stress response in Arabidopsis by MPK6-targeted phosphorylation of the heat stress factor HsfA2. PeerJ 2013, 1, e59. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Salamó, I.; Papdi, C.; Rigó, G.; Zsigmond, L.; Vilela, B.; Lumbreras, V.; Nagy, I.; Horváth, B.; Domoki, M.; Darula, Z.; et al. The heat shock factor A4A confers salt tolerance and is regulated by oxidative stress and the mitogen-activated protein kinases MPK3 and MPK6. Plant Physiol. 2014, 165, 319–334. [Google Scholar] [CrossRef]
- Wang, L.; Guo, Y.; Jia, L.; Chu, H.; Zhou, S.; Chen, K.; Wu, D.; Zhao, L. Hydrogen peroxide acts upstream of nitric oxide in the heat shock pathway in Arabidopsis seedlings. Plant Physiol. 2014, 164, 2184–2196. [Google Scholar] [CrossRef]
- Wu, D.; Chu, H.; Jia, L.; Chen, K.; Zhao, L. A feedback inhibition between nitric oxide and hydrogen peroxide in the heat shock pathway in Arabidopsis seedlings. Plant Growth Regul. 2015, 75, 503–509. [Google Scholar] [CrossRef]
- Jia, D.; Liu, Y.H.; Zhang, B.; Ji, Z.Y.; Wang, Y.X.; Gao, L.L.; Ma, R.Y. Induction of heat shock protein genes is the hallmark of egg heat tolerance in Agasicles hygrophila (coleoptera: Chrysomelidae). J. Econ. Entomol. 2020, 113, 1972–1981. [Google Scholar] [CrossRef]
- Hu, J.; Huang, X.; Chen, L.; Sun, X.; Lu, C.; Zhang, L.; Wang, Y.; Zuo, J. Site-specific nitrosoproteomic identification of endogenously S-nitrosylated proteins in Arabidopsis. Plant Physiol. 2015, 167, 1731–1746. [Google Scholar] [CrossRef]
- Gao, H.; Brandizzi, F.; Benning, C.; Larkin, R.M. A membrane-tethered transcription factor defines a branch of the heat stress response in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 2008, 105, 16398–16403. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.T.; Li, G.L.; Chang, H.; Sun, D.Y.; Zhou, R.G.; Li, B. Calmodulin-binding protein phosphatase PP7 is involved in thermotolerance in Arabidopsis. Plant Cell Environ. 2007, 30, 156–164. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.T.; Gao, F.; Li, G.L.; Han, J.L.; Liu, D.L.; Sun, D.Y.; Zhou, R.G. The calmodulin-binding protein kinase 3 is part of heat-shock signal transduction in Arabidopsis thaliana. Plant J. 2008, 55, 760–773. [Google Scholar] [CrossRef] [PubMed]
- Mogk, A.; Bukau, B. Role of sHsps in organizing cytosolic protein aggregation and disaggregation. Cell Stress Chaperones 2017, 22, 493–502. [Google Scholar] [CrossRef] [PubMed]
- Haslbeck, M.; Vierling, E. A first line of stress defense: Small heat shock proteins and their function in protein hmeostasis. J. Mol. Biol. 2015, 427, 1537–1548. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, W.; Kang, X.; Zhao, L. Arabidopsis CaM3 inhibits nitric oxide accumulation and improves thermotolerance by promoting S-nitrosoglutathione reductase via direct binding. Plant Growth Regul. 2019, 90, 41–50. [Google Scholar] [CrossRef]
- Alamri, S.A.; Siddiqui, M.H.; Al-Khaishany, M.Y.; Khan, M.N.; Ali, H.M.; Alakeel, K.A. Nitric oxide-mediated cross-talk of proline and heat shock proteins induce thermotolerance in Vicia faba L. Environ. Exp. Bot. 2019, 161, 290–302. [Google Scholar] [CrossRef]
- Piterková, J.; Luhová, L.; Mieslerová, B.; Lebeda, A.; Petrivalsky, M. Nitric oxide and reactive oxygen species regulate the accumulation of heat shock proteins in tomato leaves in response to heat shock and pathogen infection. Plant Sci. 2013, 207, 57–65. [Google Scholar] [CrossRef]
- Link, V.; Sinha, A.K.; Vashista, P.; Hofmann, M.G.; Proels, R.K.; Ehness, R.; Roitsch, T. A heat-activated MAP kinase in tomato: A possible regulator of the heat stress response. FEBS Lett. 2002, 531, 179–183. [Google Scholar] [CrossRef]
- Sunkar, R.; Kapoor, A.; Zhu, J.K. Posttranscriptional induction of two Cu/Zn superoxide dismutase genes in Arabidopsis is mediated by downregulation of miR398 and important for oxidative stress tolerance. Plant Cell 2006, 18, 2415. [Google Scholar] [CrossRef]
- Jiang, C.H.; Xu, J.Y.; Zhang, H.; Zhang, X.; Shi, J.L.; Li, M.; Ming, F. A cytosolic class I small heat shock protein, RcHSP17.8, of Rosa chinensis confers resistance to a variety of stresses to Escherichia coli, yeast and Arabidopsis thaliana. Plant Cell Environ. 2009, 32, 1046–1059. [Google Scholar] [CrossRef] [PubMed]
- Sun, L.P.; Liu, Y.; Kong, X.P.; Zhang, D.; Pan, J.W.; Zhou, Y.; Wang, L.; Li, D.Q.; Yang, X.H. ZmHSP16.9, a cytosolic class I small heat shock protein in maize (Zea mays), confers heat tolerance in transgenic tobacco. Plant Cell Rep. 2012, 31, 1473–1484. [Google Scholar] [CrossRef] [PubMed]
- Lin, S.; Liu, Z.; Sun, S.Y.; Xue, F.Y.; Li, H.J.; Tursun, A.; Cao, L.C.; Zhang, L.; Wilson, Z.A.; Zhang, D.B.; et al. Rice heat shock protein 60-3B maintains male fertility under high temperature by starch granule biogenesis. Plant Physiol. 2023, 192, 2301–2317. [Google Scholar] [CrossRef] [PubMed]
- Mu, C.J.; Wang, S.B.; Zhang, S.J.; Pan, J.J.; Chen, N.; Li, X.F.; Wang, Z.Y.; Liu, H. Small heat shock protein LimHSP16.45 protects pollen mother cells and tapetal cells against extreme temperatures during late zygotene to pachytene stages of meiotic prophase I in David Lily. Plant Cell Rep. 2011, 30, 1981–1989. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, Q.X.; Gao, Y.K.; Pan, H.T.; Shi, S.C.; Wang, Y. Overexpression of heat shock protein gene PfHSP21.4 in Arabidopsis thaliana enhances heat tolerance. Acta Physiol. Plant 2014, 36, 1555–1564. [Google Scholar] [CrossRef]
- Huang, L.J.; Cheng, G.X.; Khan, A.; Wei, A.M.; Yu, Q.H.; Yang, S.B.; Luo, D.X.; Gong, Z.H. CaHSP16.4, a small heat shock protein gene in pepper, is involved in heat and drought tolerance. Protoplasma 2019, 256, 39–51. [Google Scholar] [CrossRef]
- Zhou, Y.Z.; Wang, Y.; Xu, F.X.; Song, C.X.; Yang, X.; Zhang, Z.; Yi, M.F.; Ma, N.; Zhou, X.F.; He, J.N. Small HSPs play an important role in crosstalk between HSF-HSP and ROS pathways in heat stress response through transcriptomic analysis in lilies (Lilium longiflorum). BMC Plant Biol. 2022, 22, 202. [Google Scholar] [CrossRef]
- Ding, X.L.; Lv, M.L.; Liu, Y.; Guo, Q.L.; Gai, J.Y.; Yang, S.P. A small heat shock protein GmHSP18.5a improves the male fertility restorability of cytoplasmic male sterility-based restorer line under high temperature stress in soybean. Plant Sci. 2023, 337, 111867. [Google Scholar] [CrossRef]
- Ma, W.; Li, J.; Liu, F.J.; Zhang, T.Z.; Guan, X.Y. GhHSP24.7 mediates mitochondrial protein acetylation to regulate stomatal conductance in response to abiotic stress in cotton. Crop J. 2023, 11, 1128–1139. [Google Scholar] [CrossRef]
- Do, J.M.; Kim, H.J.; Shin, S.Y.; Park, S.I.; Kim, J.J.; Yoon, H.S. OsHSP17.9, a small heat shock protein, confers improved productivity and tolerance to high temperature and salinity in a natural paddy field in transgenic rice plants. Agriculture 2023, 13, 931. [Google Scholar] [CrossRef]
- Zhao, L.Y.; Zhao, Y.Q.; Wang, L.; Hou, Y.Y.; Bao, Y.Q.; Jia, Z.Y.; Zheng, Y.H.; Jin, P. Hot water treatment improves peach fruit cold resistance through PpHSFA4c-mediated HSF-HSP and ROS pathways. Postharvest Biol. Technol. 2023, 199, 112272. [Google Scholar] [CrossRef]
- Ding, Y.F.; Zhou, M.; Wang, K.; Qu, A.L.; Hu, S.S.; Jiang, Q.; Yi, K.K.; Wang, F.J.; Cai, C.; Zhu, C.; et al. Rice DST transcription factor negatively regulates heat tolerance through ROS-mediated stomatal movement and heat-responsive gene expression. Front. Plant Sci. 2023, 14, 1068296. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.H.; Wu, Y.L.; Luo, L.; Ma, Y.Z.; Li, Y.L.; Ma, H.H.; Luo, A.Y.; Zhang, R.; Zhu, L.F.; Lin, Y.J.; et al. Proteomic analysis reveals that the heat shock proteins 70-17 and BiP5 enhance cotton male fertility under high-temperature stress by reducing the accumulation of ROS in anthers. Ind. Crop. Prod. 2022, 188, 115693. [Google Scholar] [CrossRef]
- Qin, Q.Q.; Zhao, Y.J.; Zhang, J.J.; Chen, L.; Si, W.N.; Jiang, H.Y. A maize heat shock factor ZmHsf11 negatively regulates heat stress tolerance in transgenic plants. BMC Plant Biol. 2022, 22, 406. [Google Scholar] [CrossRef] [PubMed]
- Khatun, M.; Borphukan, B.; Alam, I.; Keya, C.A.; Panditi, V.; Khan, H.; Huq, S.; Reddy, M.K.; Salimullah, M. Mitochondria-targeted SmsHSP24.1 overexpression stimulates early seedling vigor and stress tolerance by multi-pathway transcriptome-reprogramming. Front. Plant Sci. 2021, 12, 741898. [Google Scholar] [CrossRef] [PubMed]
- Ni, Z.Y.; Liu, N.; Yu, Y.H.; Bi, C.X.; Chen, Q.J.; Qu, Y.Y. The cotton 70-kDa heat shock protein GhHSP70-26 plays a positive role in the drought stress response. Environ. Exp. Bot. 2021, 191, 104628. [Google Scholar] [CrossRef]
- Gangadhar, B.H.; Mishra, R.K.; Kappachery, S.; Baskar, V.; Venkatesh, J.; Nookaraju, A.; Thiruvengadam, M. Enhanced thermotolerance in transgenic potato overexpressing hydrogen peroxide-producing germin-like protein (GLP). Genomics 2021, 113, 3224–3234. [Google Scholar] [CrossRef] [PubMed]
- Bi, A.Y.; Wang, T.; Wang, G.Y.; Zhang, L.; Wassie, M.; Amee, M.; Xu, H.W.; Hu, Z.R.; Liu, A.; Fu, J.M.; et al. Stress memory gene FaHSP17.8-CII controls thermotolerance via remodeling PSII and ROS signaling in tall fescue. Plant Physiol. 2021, 187, 1163–1176. [Google Scholar] [CrossRef]
- Pollastri, S.; Sukiran, N.A.; Jacobs, B.; Knight, M.R. Chloroplast calcium signalling regulates thermomemory. J. Plant Physiol. 2021, 264, 153470. [Google Scholar] [CrossRef]
- Fan, M.H.; Tang, X.W.; Yang, Z.X.; Wang, J.X.; Zhang, X.L.; Yan, X.J.; Li, P.; Xu, N.J.; Liao, Z. Integration of the transcriptome and proteome provides insights into the mechanism calcium regulated of Ulva prolifera in response to high-temperature stress. Aquaculture 2022, 557, 738344. [Google Scholar] [CrossRef]
- Ding, L.P.; Wu, Z.; Teng, R.D.; Xu, S.J.; Cao, X.; Yuan, G.Z.; Zhang, D.H.; Teng, N.J. LlWRKY39 is involved in thermotolerance by activating LlMBF1c and interacting with LlCaM3 in lily (Lilium longiflorum). Hort. Res. 2021, 8, 36. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.T.; Sun, H.Y.; Ding, Y.; Zang, X.N.; Zhang, X.C. A novel heat shock protein from Gracilariopsis lemaneiformis: Gene cloning and transcription analysis in response to heat stress. J. Appl. Phycol. 2018, 30, 3623–3631. [Google Scholar] [CrossRef]
- Cha, J.Y.; Su’udi, M.; Kim, W.Y.; Kim, D.R.; Kwak, Y.S.; Son, D. Functional characterization of orchardgrass cytosolic Hsp70 (DgHsp70) and the negative regulation by Ca2+/AtCaM2 binding. Plant Physiol. Biochem. 2012, 58, 29–36. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, T.; von Braun, J. Climate change impacts on global food security. Science 2013, 341, 508–513. [Google Scholar] [CrossRef]
- Zhu, T.; De Lima, C.F.F.; De Smet, I. The heat is on: How crop growth, development and yield respond to high temperature. J. Exp. Bot. 2021, 58, 635–645. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Gene Type | Species | Gene Names | Functions | References |
|---|---|---|---|---|
| Cyclic nucleotide -gated channels (CNGCs) | Physcomitrella patens | CNGCb | Sensitive to heat stress | [21] |
| Arabidopsis thaliana | AtCNGC2 | Sensitive to heat stress at the seedling stage; Tolerance to heat stress at the reproductive stage | [21,22,23] | |
| AtCNGC4 | Tolerance to extreme temperatures; Response to pathogen infection | [24] | ||
| AtCNGC6 | Regulates tolerance to extreme temperatures together with hydrogen peroxide (H2O2) and Nitric oxide (NO) | [25,26,27] | ||
| Oryza sativa | OsCNGC14 | Tolerance to extreme temperatures | [28] | |
| OsCNGC16 | Tolerance to extreme temperatures | [28] | ||
| Glutamate receptor-like channels (GLRs) | Arabidopsis thaliana | AtGLR3.3 | Response to pathogen infection | [43] |
| Vicia faba | VfGLR3.5 | Tolerance to drought | [44] | |
| Solanum lycopersicum | SlGLR3.3 | Tolerance to cold stress by regulating apoplastic H2O2 production and redox homeostasis | [45] | |
| SlGLR3.5 | Tolerance to cold stress by regulating apoplastic H2O2 production and redox homeostasis | [45] | ||
| Zea mays | ZmGLR | Tolerance to heat stress | [46] | |
| Annexins | Nelumbo nucifera | NnANN1 | Tolerance to heat stress | [58] |
| Glycine max | GmANN | Tolerance to high temperatures and humidity stress | [59] | |
| Oryza sativa | OsANA1 | Tolerance to heat shock, H2O2 treatment, and abiotic stress | [60] | |
| Raphanus sativus | RsANN | Tolerance to heat, drought, salinity, oxidation, and ABA stress | [61] | |
| Reduced hyperosmolarity-induced [Ca2+] increase channels (OSCAs) | Zea mays | ZmOSCA1.4 | Gene expression increases in response to heat stress | [66] |
| ZmOSCA2.1 | Gene expression increases in response to heat stress | [66] | ||
| ZmOSCA2.2 | Gene expression increases in response to heat stress | [66] | ||
| ZmOSCA2.5 | Gene expression increases in response to heat stress | [66] | ||
| ZmOSCA3.1 | Gene expression increases in response to heat stress | [66] | ||
| ZmOSCA4.1 | Gene expression increases in response to heat stress | [66] | ||
| ZmOSCA1.3 | Gene expression decreases in response to heat stress | [66] | ||
| ZmOSCA1.5 | Gene expression decreases in response to heat stress | [66] | ||
| ZmOSCA2.4 | Gene expression decreases in response to heat stress | [66] | ||
| Mechanosensitive-like channels (MSLs) | Arabidopsis thaliana | AtMSL2 | Tolerance to high osmotic stress | [67] |
| AtMSL3 | Tolerance to high osmotic stress | [67] | ||
| AtMSL8 | Response to PM distortion during pollen grain rehydration and germination | [68] | ||
| AtMSL9 | Exhibits MS ion channel activity | [69] | ||
| AtMSL10 | Exhibits MS ion channel activity | [69] | ||
| Oryza sativa | OsMSLs | Responses to plant growth, development, and various stressors | [70] | |
| Mid1-complementing activity” channels (MCAs) | Arabidopsis thaliana | AtMCA1 | Tolerance to cold stress | [73,74] |
| AtMCA2 | Tolerance to cold stress | [73,74] | ||
| Piezo channels (MSPs) | Arabidopsis thaliana | AtPiezo | Response to virus infection | [77] |
| Gene Type | Species | Gene Names | Functions | References |
|---|---|---|---|---|
| Calmodulins (CaMs) | Arabidopsis thaliana | AtCaM3 | Tolerance to heat stress | [87,88] |
| Cucumis sativus | CsCaM3 | Tolerance to heat stress; Safeguards against oxidative damage | [89] | |
| Oryza sativa | OsCaM1-1 | Tolerance to heat stress | [90] | |
| CaM-like proteins (CMLs) | Arabidopsis thaliana | AtCML12 | Gene expression significantly increased under heat stress | [92] |
| AtCML24 | Gene expression significantly increased under heat stress | [92] | ||
| Oryza sativa | OsMSR2 | Response to cold, drought, and heat stress | [93] | |
| Solanum lycopersicum | SlCML39 | Negative impact on high-temperature tolerance | [94] | |
| Ca2+-dependent protein kinases (CDPKs) | Arabidopsis thaliana | AtCPK1 | Tolerance to salt, cold, and heat | [100] |
| Lycopersicon esculentum | LeCPK28 | Tolerance to heat stress | [101] | |
| Lycopersicon esculentum | LeCPK2 | Tolerance to heat stress | [102] | |
| Zea mays | ZmCDPK7 | Tolerance to heat stress | [103] | |
| ZmCK3 | Exhibits increased transcription in response to drought, salt, and heat stress | [104] | ||
| Setaria italica | SiCDPK7 | Response to extreme temperature stress | [105] | |
| Vitis amurensis | VaCPK29 | Response to heat and osmotic stress | [106] | |
| Calcineurin B-like proteins (CBLs) | Oryza sativa | OsCBL8 | Enhances resistance to high temperatures and pathogens | [115] |
| CBL-interacting protein kinases (CIPKs) | Oryza sativa | OsCIPK17 | Enhances resistance to high temperatures and pathogens | [115] |
| Ananas comosus | AcCIPK5 | Promotes tolerance to salt, osmotic stress, and cold stress while negatively regulating heat stress responses | [116] |
| Species | HS proteins (HSPs) Names | References | |
|---|---|---|---|
| NO | Arabidopsis thaliana | HSP18.2 | [88] |
| HSP17.7, HSP21 | [137,146] | ||
| Vicia faba | Hsp17.6, Hsp70, Hsp90-1, and Hsp101 | [147] | |
| Solanum lycopersicum | HSP70 | [148] | |
| Solanum chmielewskii | |||
| Reactive oxygen species (ROS) | Arabidopsis thaliana | HSP17.7, HSP21 | [26] |
| Solanum lycopersicum | HSP40 | [149] | |
| Arabidopsis thaliana | HSP17.6, HSP18.6 | [150] | |
| Rosa chinensis | HSP17.8 | [151] | |
| Zea mays | HSP16.9 | [152] | |
| Oryza sativa | HSP60-B | [153] | |
| Lilium davidii | HSP16.45 | [154] | |
| Primula malacoides | HSP21.4 | [155] | |
| Capsicum annuum | HSP16.4 | [156] | |
| Lilium longiflorum | HSP70, HSP22.1, HSP22.2, HSP17.6, HSP20 | [157] | |
| Glycine max | HSP18.5a | [158] | |
| Gossypium hirsutum | HSP24.7 | [159] | |
| Oryza sativa | HSP17.9 | [160] | |
| Prunus persica | HSP18.5, HSP70, HSP80 | [161] | |
| Oryza sativa | HSP80, HSP74, HSP58, sHSPs | [162] | |
| Gossypium hirsutum | HSP70-17 | [163] | |
| Zea mays | HSP17 | [164] | |
| Solanum melongena | HSP24.1 | [165] | |
| Gossypium hirsutum | HSP70-26 | [166] | |
| Solanum tuberosum | HSP70, HSP90, HSP20 | [167] | |
| Festuca arundinacea | HSP17.8 | [168] | |
| Ca2+ sensor | Arabidopsis thaliana | HSP18.2, HSP25.3, HSP70 | [25] |
| Arabidopsis thaliana | HSP18.2 | [87,88] | |
| Gracilariopsis lemaneiformis | HSP70s, HSP90s | [168] | |
| Arabidopsis thaliana | HSP17.6 | [169] | |
| Ulva prolifera | HSP70, HSP90 | [170] | |
| Lilium longiflorum | HSP101 | [171] | |
| Gracilariopsis lemaneiformis | HSP70-1, HSP70-2 | [172] | |
| Dactylis glomerata | HSP70 | [173] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kang, X.; Zhao, L.; Liu, X. Calcium Signaling and the Response to Heat Shock in Crop Plants. Int. J. Mol. Sci. 2024, 25, 324. https://doi.org/10.3390/ijms25010324
Kang X, Zhao L, Liu X. Calcium Signaling and the Response to Heat Shock in Crop Plants. International Journal of Molecular Sciences. 2024; 25(1):324. https://doi.org/10.3390/ijms25010324
Chicago/Turabian StyleKang, Xinmiao, Liqun Zhao, and Xiaotong Liu. 2024. "Calcium Signaling and the Response to Heat Shock in Crop Plants" International Journal of Molecular Sciences 25, no. 1: 324. https://doi.org/10.3390/ijms25010324
APA StyleKang, X., Zhao, L., & Liu, X. (2024). Calcium Signaling and the Response to Heat Shock in Crop Plants. International Journal of Molecular Sciences, 25(1), 324. https://doi.org/10.3390/ijms25010324