
H2O2 Significantly Affects Larix kaempferi × Larix olgensis Somatic Embryogenesis

 ,
,  ,
,
Abstract
1. Introduction
2. Results
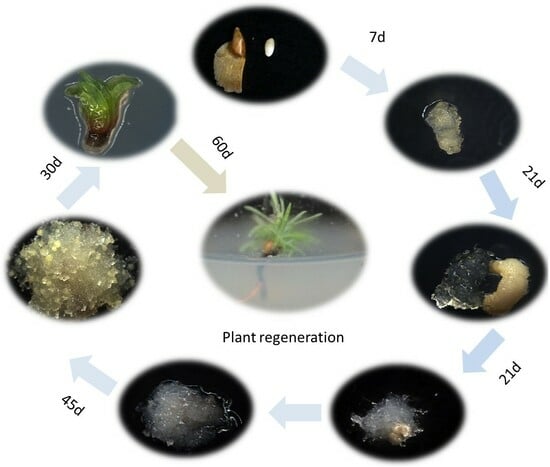
2.1. Establishment of a Somatic Embryogenesis System in Hybrid Larch
2.2. ABA and PEG Are Important Stress Signals for Somatic Embryo Development
2.3. Carbon Source Effect on Somatic Embryo Development
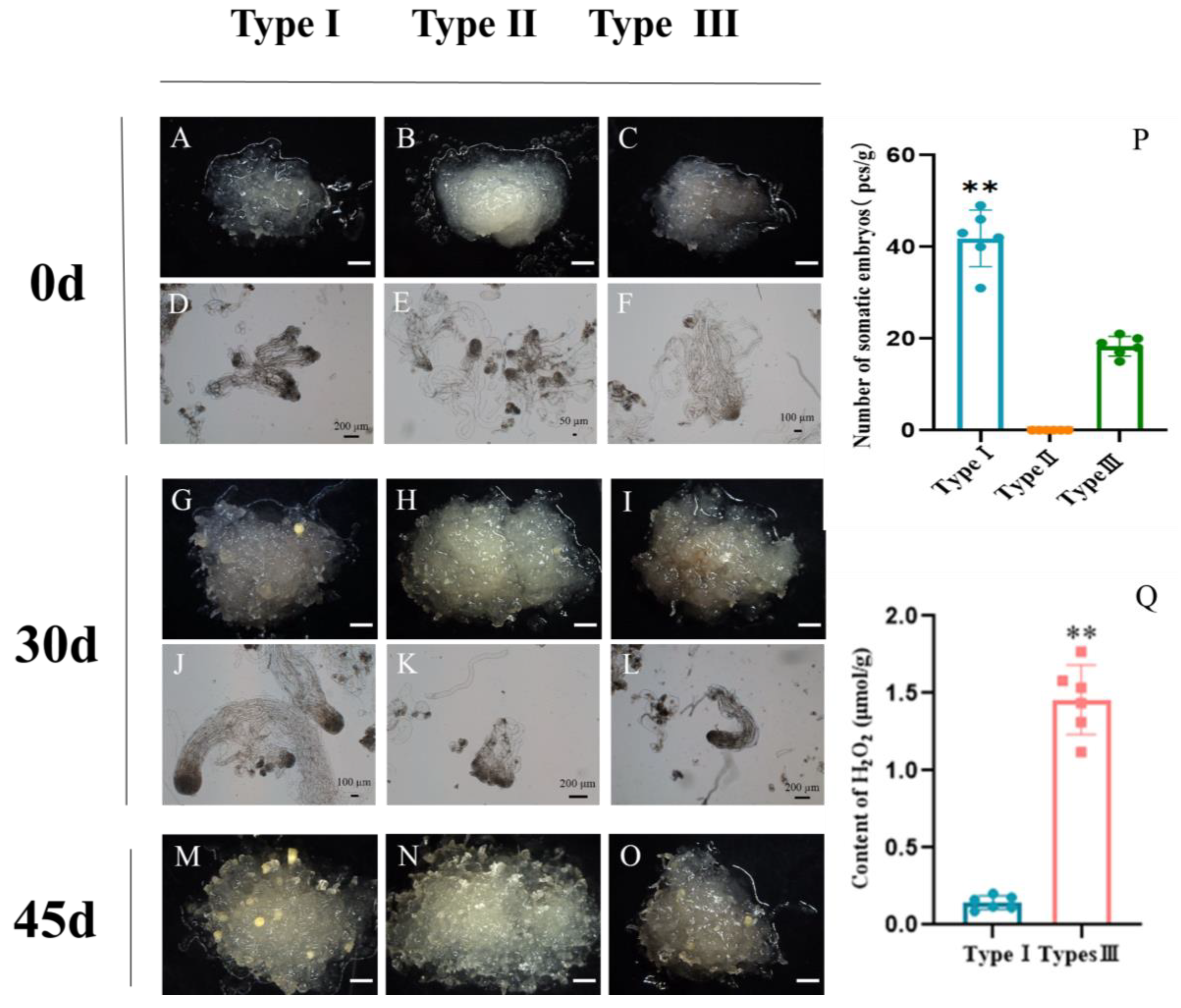
2.4. Effects of ROS Homeostasis on Somatic Embryogenesis in Larch
3. Discussion
3.1. Establishment of a Hybrid Larch Somatic Embryogenesis System
3.2. Homeostasis of Reactive Oxygen Species Affects Somatic Embryogenesis
4. Materials and Methods
4.1. Plant Materials
4.2. Medium and Culture Conditions
4.2.1. ESM Induction
4.2.2. ESM Proliferation
4.2.3. Somatic Embryo Induction
4.2.4. Plant Regeneration
4.3. PEG and ABA Effect on Somatic Embryogenesis
4.4. Carbon Source Effect on Somatic Embryogenesis
4.5. Hydrogen Peroxide Content Detection
4.6. H2O2 Effect on Somatic Embryogenesis
4.7. Statistical Methods
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yan, X.; Wang, K.; Zheng, K.; Zhang, L. Efficient organogenesis and taxifolin production system from mature zygotic embryos and needles in larch. For. Res. 2023, 3, 4. [Google Scholar] [CrossRef]
- Sun, C.; Xie, Y.H.; Li, Z.; Liu, Y.J.; Sun, X.M.; Li, J.J.; Quan, W.P.; Zeng, Q.Y.; Van de Peer, Y.; Zhang, S.G. The Larix kaempferi genome reveals new insights into wood properties. J. Integr. Plant Biol. 2022, 64, 1364–1373. [Google Scholar] [CrossRef] [PubMed]
- Baltunis, B.S.; Greenwood, M.S. Eysteinsson Hybrid vigor in Larix: Growth of intra- and interspecific hybrids of Larix decidua, L-laricina, and L-kaempferi after 5-years. Silvae Genet. 1998, 47, 288–293. [Google Scholar] [CrossRef]
- Sun, X.; Zhang, S.; Wang, X.; Qi, L.; Wang, W.; Chen, B. Genetic Variation on Rooting Ability of Stem Cutting and Juvenile Growth of Larix kaempferi × L. olgensis Interspecific Hybrids. Sci. Silvae Sin. 2008, 44, 41–47. [Google Scholar] [CrossRef]
- Jiang, S.; Chen, X.; Gao, Y.; Cui, Y.; Kong, L.; Zhao, J.; Zhang, J. Plant Regeneration via Somatic Embryogenesis in Larix principis-rupprechtii Mayr. Forests 2021, 12, 1335. [Google Scholar] [CrossRef]
- Fernandes, P.; Rodriguez, E.; Pinto, G.; Roldan-Ruiz, I.; De Loose, M.; Santos, C. Cryopreservation of Quercus suber somatic embryos by encapsulation-dehydration and evaluation of genetic stability. Tree Physiol. 2008, 28, 1841–1850. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.S.; Lelu-Walter, M.A.; Harvengt, L.; Trontin, J.F.; MacEacheron, I.; Klimaszewska, K.; Bonga, J.M. Initiation of somatic embryogenesis in Pinus banksiana, P-strobus, P. pinaster, and P-sylvestris at three laboratories in Canada and France. Plant Cell Tissue Organ Cult. 2006, 86, 87–101. [Google Scholar] [CrossRef]
- Yang, M.; Zhang, D.; Li, Z.; Jin, X.; Ding, G. Somatic Embryogenesis with Immature Embryos of Masson Pine (Pinus massoniana Lamb. Plant Physiol. J. 2011, 47, 904–912. [Google Scholar] [CrossRef]
- Dowling, J.K.; Tate, M.D.; Golenbock, D.T.; Mansell, A. Somatic Embryogenesis in Juniperus procera Using Juniperus communis as a Model. Cytokine 2014, 70, 37. [Google Scholar] [CrossRef]
- Filonova, L.H.; Bozhkov, P.V.; von Arnold, S. Developmental pathway of somatic embryogenesis in Picea abies as revealed by time-lapse tracking. J. Exp. Bot. 2000, 51, 249–264. [Google Scholar] [CrossRef]
- Feher, A. Somatic embryogenesis—Stress-induced remodeling of plant cell fate. Biochim Biophys. Acta 2015, 1849, 385–402. [Google Scholar] [CrossRef] [PubMed]
- Qi, L.; Han, Y.; Han, S.; Wang, J. Effects of Maltose, NAA and ABA on Somatic Maturation and Radicle Rooting of Larix principis-rupprechtii. Res. Inst. For. 2004, 40, 53–60. [Google Scholar]
- Salo, H.M.; Sarjala, T.; Jokela, A.; Haggman, H.; Vuosku, J. Moderate stress responses and specific changes in polyamine metabolism characterize Scots pine somatic embryogenesis. Tree Physiol. 2016, 36, 392–402. [Google Scholar] [CrossRef] [PubMed]
- Libik, M.; Konieczny, R.; Pater, B.; Slesak, I.; Miszalski, Z. Differences in the activities of some antioxidant enzymes and in H2O2 content during rhizogenesis and somatic embryogenesis in callus cultures of the ice plant. Plant Cell Rep. 2005, 23, 834–841. [Google Scholar] [CrossRef] [PubMed]
- Hirt, H. Connecting oxidative stress, auxin, and cell cycle regulation through a plant mitogen-activated protein kinase pathway. Proc. Natl. Acad. Sci. USA 2000, 97, 2405–2407. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Han, S.; Ding, X. Transcriptome Analysis of mRNA and miRNA in Somatic Embryos of Larix leptolepis Subjected to Hydrogen Treatment. Int. J. Mol. Sci. 2016, 17, 1951. [Google Scholar] [CrossRef]
- Fatima, S.; Mujib, A.; Samaj, J. Anti-oxidant enzyme responses during in vitro embryogenesis in Catharanthus roseus. J. Horticult. Sci. Biotechnol. 2011, 86, 569–574. [Google Scholar] [CrossRef]
- Hao, Z.; Wu, H.; Zheng, R.; Li, R.; Zhu, Z.; Chen, Y.; Lu, Y.; Cheng, T.; Shi, J.; Chen, J. The plant peptide hormone phytosulfokine promotes somatic embryogenesis by maintaining redox homeostasis in Cunninghamia lanceolata. Plant J. 2023, 113, 716–733. [Google Scholar] [CrossRef]
- Cui, K.; Pei, X. Effects of modulation of abscisic acid during somatic embryogenesis in Lycium barbarum L. Acta Biol. Exp. Sin. 1998, 02, 195–201. [Google Scholar]
- Hakman, I.; Arnold, S.V. Plantlet Regeneration through Somatic Embryogenesis in Picea abies (Norway Spruce). J. Plant Physiol. 1985, 121, 149–158. [Google Scholar] [CrossRef]
- Vaelimaeki, S.; Hazubska-Przybyl, T.; Ratajczak, E.; Tikkinen, M.; Varis, S.; Aronen, T. Somatic Embryo Yield and Quality From Norway Spruce Embryogenic Tissue Proliferated in Suspension Culture. Front. Plant Sci. 2021, 12, 791549. [Google Scholar] [CrossRef] [PubMed]
- Castander-Olarieta, A.; Montalban, I.A.; Moncalean, P. Multi-strategy approach towards optimization of maturation and germination in radiata pine somatic embryogenesis. Plant Cell Tissue Organ Cult. 2023, 153, 173–190. [Google Scholar] [CrossRef]
- Zavattieri, M.A.; Frederico, A.M.; Lima, M.; Sabino, R.; Arnholdt-Schmitt, B. Induction of somatic embryogenesis as an example of stress-related plant reactions. Electron. J. Biotechnol. 2010, 13, 12–13. [Google Scholar] [CrossRef]
- Zhou, X.; Zheng, R. Desiccation Treatment and Endogenous IAA Levels Are Key Factors Influencing High Frequency Somatic Embryogenesis in Cunninghamia lanceolata (Lamb.) Hook. Front. Plant Sci. 2017, 8, 2054. [Google Scholar] [CrossRef] [PubMed]
- Lelu-Walter, M.A.; Paques, L.E. Simplified and improved somatic embryogenesis of hybrid larches (Larix imeseurolepis and Larix imesmarschlinsii). Perspectives for breeding. Ann. For. Sci. 2009, 66, 104. [Google Scholar] [CrossRef]
- Liu, C.; Xia, X.; Yin, W.; Huang, L.; Zhou, J. Shoot regeneration and somatic embryogenesis from needles of redwood (Sequoia sempervirens (D.Don.) Endl.). Plant Cell Rep. 2006, 25, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Malabadi, R.B.; Van Staden, J. Somatic embryogenesis from vegetative shoot apices of mature trees of Pinus patula. Tree Physiol. 2005, 25, 11–16. [Google Scholar] [CrossRef]
- Tret’Yakova, I.N.; Barsukova, A.V. Somatic embryogenesis in in vitro culture of three larch species. Russ. J. Dev. Biol. 2012, 43, 353–361. [Google Scholar] [CrossRef]
- Krutovsky, K.V.; Tretyakova, I.N.; Oreshkova, N.V.; Pak, M.E.; Kvitko, O.V.; Vaganov, E.A. Somaclonal variation of haploid in vitro tissue culture obtained from Siberian larch (Larix sibirica Ledeb.) megagametophytes for whole genome de novo sequencing. Vitr. Cell. Dev. Biol.-Plant 2014, 50, 655–664. [Google Scholar] [CrossRef]
- Potters, G.; Pasternak, T.P.; Guisez, Y.; Palme, K.J.; Jansen, M.A. Stress-induced morphogenic responses: Growing out of trouble? Trends Plant Sci. 2007, 12, 98–105. [Google Scholar] [CrossRef]
- Maruyama, T.E. Polyethylene glycol improves somatic embryo maturation in big-leaf mahogany (Swietenia macrophylla King, Meliaceae). Bull. For. For. Prod. Res. Inst. 2009, 8, 167–173. [Google Scholar]
- Acanda, Y.; Martínez, Ó.; Prado, M.J.; González, M.V.; Rey, M. Changes in abscisic acid metabolism in relation to the maturation of grapevine (Vitis vinifera L., cv. Mencía) somatic embryos. BMC Plant Biol. 2020, 20, 487. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Attree, S.M.; Fowke, L.C. Changes of endogenous hormone levels in developing seeds, zygotic embryos and megagametophytes in Picea glauca. Physiol. Plant. 2010, 101, 23–30. [Google Scholar] [CrossRef]
- Nunes, S.; Marum, L.; Farinha, N.; Pereira, V.T.; Almeida, T.; Sousa, D.; Mano, N.; Figueiredo, J.; Dias, M.C.; Santos, C. Somatic embryogenesis of hybrid Pinus elliottii var. elliottii x P-caribaea var. hondurensis and ploidy assessment of somatic plants. Plant Cell 2018, 132, 71–84. [Google Scholar] [CrossRef]
- Gupta, P.K.; Pullman, G.S. Method for reproducing coniferous plants by somatic embryogenesis using abscisic acid and osmotic potential variation: Pramod gupta, Gerald S Pullman assigned to Weyerhaeuser Company. Biotechnol. Adv. 1991, 9, 684. [Google Scholar] [CrossRef]
- Kanchan, V.; Neha, U.; Nitin, K.; Gaurav, Y.; Jaspreet, S.; Mishra, R.K.; Vivek, K.; Rishi, V.; Upadhyay, R.G.; Mayank, P. Abscisic Acid Signaling and Abiotic Stress Tolerance in Plants: A Review on Current Knowledge and Future Prospects. Front. Plant Sci. 2017, 8, 161. [Google Scholar] [CrossRef]
- Satoshi, K.; Kaoru, S.; Paul, T.; Takamasa, S.; Sachihiro, M. Characterization of somatic embryogenesis initiated from the Arabidopsis shoot apex. Dev. Biol. 2018, 442, 13–27. [Google Scholar] [CrossRef]
- Chen, B.; Martijn, F.; Dekkers, B.J.W.; Lena, M.; Wilma, V.E.G.; Angenent, G.C.; Yang, Z.; Kim, B. ABA signalling promotes cell totipotency in the shoot apex of germinating embryos. J. Exp. Bot. 2021, 72, 6418–6436. [Google Scholar] [CrossRef]
- Han, M.; Wang, B.; Song, G.; Shi, S. Comparative study of alleviation effects of DMTU and PCIB on root growth inhibition in two tall fescue varieties under cadmium stress. Ecotoxicol. Environ. Saf. 2020, 196, 110528. [Google Scholar] [CrossRef]
- Huang, Z.; Li, H. Control of oxidative stress by a combination of PBU, BAP and DMTU enhances adventitious shoot formation in Eucalyptus urophylla. Plant Cell Tissue Organ Cult. 2020, 141, 533–541. [Google Scholar] [CrossRef]
- Taiba, S.; Anwar, S. High frequency plant regeneration in Indian Siris via cyclic somatic embryogenesis with biochemical, histological and SEM investigations. Ind. Crops Prod. 2015, 76, 623–637. [Google Scholar] [CrossRef]
- Cheng, W.; Wang, F.; Cheng, X.; Zhu, Q.; Sun, Y.; Zhu, H.; Sun, J. Polyamine and Its Metabolite H2O2 Play a Key Role in the Conversion of Embryogenic Callus into Somatic Embryos in Upland Cotton (Gossypium hirsutum L.). Front. Plant Sci. 2015, 6, 1063. [Google Scholar] [CrossRef] [PubMed]
- Cui, K.R.; Li, J.; Xing, G.M.; Li, J.L.; Wang, L.H.; Wang, Y.F. Effect of hydrogen peroxide on synthesis of proteins during somatic embryogenesis in Lycium barbarum. Plant Cell Tissue Organ Cult. 2002, 68, 187–193. [Google Scholar] [CrossRef]
- Otvos, K.; Pasternak, T.P.; Miskolczi, P.; Domoki, M.; Dorjgotov, D.; Szucs, A.; Bottka, S.; Dudits, D.; Feher, A. Nitric oxide is required for, and promotes auxin-mediated activation of, cell division and embryogenic cell formation but does not influence cell cycle progression in alfalfa cell cultures. Plant J. 2005, 43, 849–860. [Google Scholar] [CrossRef] [PubMed]
- Graham, N.; Caroline, L.; Amna, M. The metabolomics of oxidative stress. Phytochemistry 2015, 112, 33–53. [Google Scholar]
- Gallie, D.R. The role of L-ascorbic acid recycling in responding to environmental stress and in promoting plant growth. J. Exp. Bot. 2013, 64, 433–443. [Google Scholar] [CrossRef]
- Shi, J.; Zhen, Y.; Zheng, R. Proteome profiling of early seed development in Cunninghamia lanceolata (Lamb.) Hook. J. Exp. Bot. 2010, 61, 2367–2381. [Google Scholar] [CrossRef]
- Pullman, G.S.; Johnson, S. Somatic embryogenesis in loblolly pine (Pinus taeda L.): Improving culture initiation rates. Ann. For. Sci. 2002, 59, 663–668. [Google Scholar] [CrossRef]
- Gupta, P.K.; Durzan, D.J. Shoot multiplication from mature trees of Douglas-fir (Pseudotsuga menziesii) and sugar pine (Pinus lambertiana). Plant Cell Rep. 1985, 4, 177–179. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Collection Date | Developmental Period | Inoculation Number | ESM Induction Rate (±SD)/% |
|---|---|---|---|
| 30th May | Proembryo | 233 | 0 c |
| 10th June | Columnar embryo | 241 | 0 c |
| 28th June | Early cotyledonary embryo | 252 | 0 c |
| 8th July | Mid-cotyledon embryo | 250 | 0.8 ± 0.53 b |
| 15th July | Late cotyledon embryo | 250 | 2 ± 0.21 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, J.; Zhang, K.; Xiong, H.; Xie, Y.; Li, R.; Wu, X.; Yang, Y.; Wu, H.; Hao, Z.; Sun, X.; et al. H2O2 Significantly Affects Larix kaempferi × Larix olgensis Somatic Embryogenesis. Int. J. Mol. Sci. 2024, 25, 669. https://doi.org/10.3390/ijms25010669
Zhu J, Zhang K, Xiong H, Xie Y, Li R, Wu X, Yang Y, Wu H, Hao Z, Sun X, et al. H2O2 Significantly Affects Larix kaempferi × Larix olgensis Somatic Embryogenesis. International Journal of Molecular Sciences. 2024; 25(1):669. https://doi.org/10.3390/ijms25010669
Chicago/Turabian StyleZhu, Junjie, Kaikai Zhang, Huiru Xiong, Yunhui Xie, Rui Li, Xinru Wu, Yun Yang, Hua Wu, Zhaodong Hao, Xiaomei Sun, and et al. 2024. "H2O2 Significantly Affects Larix kaempferi × Larix olgensis Somatic Embryogenesis" International Journal of Molecular Sciences 25, no. 1: 669. https://doi.org/10.3390/ijms25010669
APA StyleZhu, J., Zhang, K., Xiong, H., Xie, Y., Li, R., Wu, X., Yang, Y., Wu, H., Hao, Z., Sun, X., & Chen, J. (2024). H2O2 Significantly Affects Larix kaempferi × Larix olgensis Somatic Embryogenesis. International Journal of Molecular Sciences, 25(1), 669. https://doi.org/10.3390/ijms25010669