
Essential Oil Constituents as Anti-Inflammatory and Neuroprotective Agents: An Insight through Microglia Modulation

 ,
,  ,
,  , and
, and 
Abstract
:1. Introduction
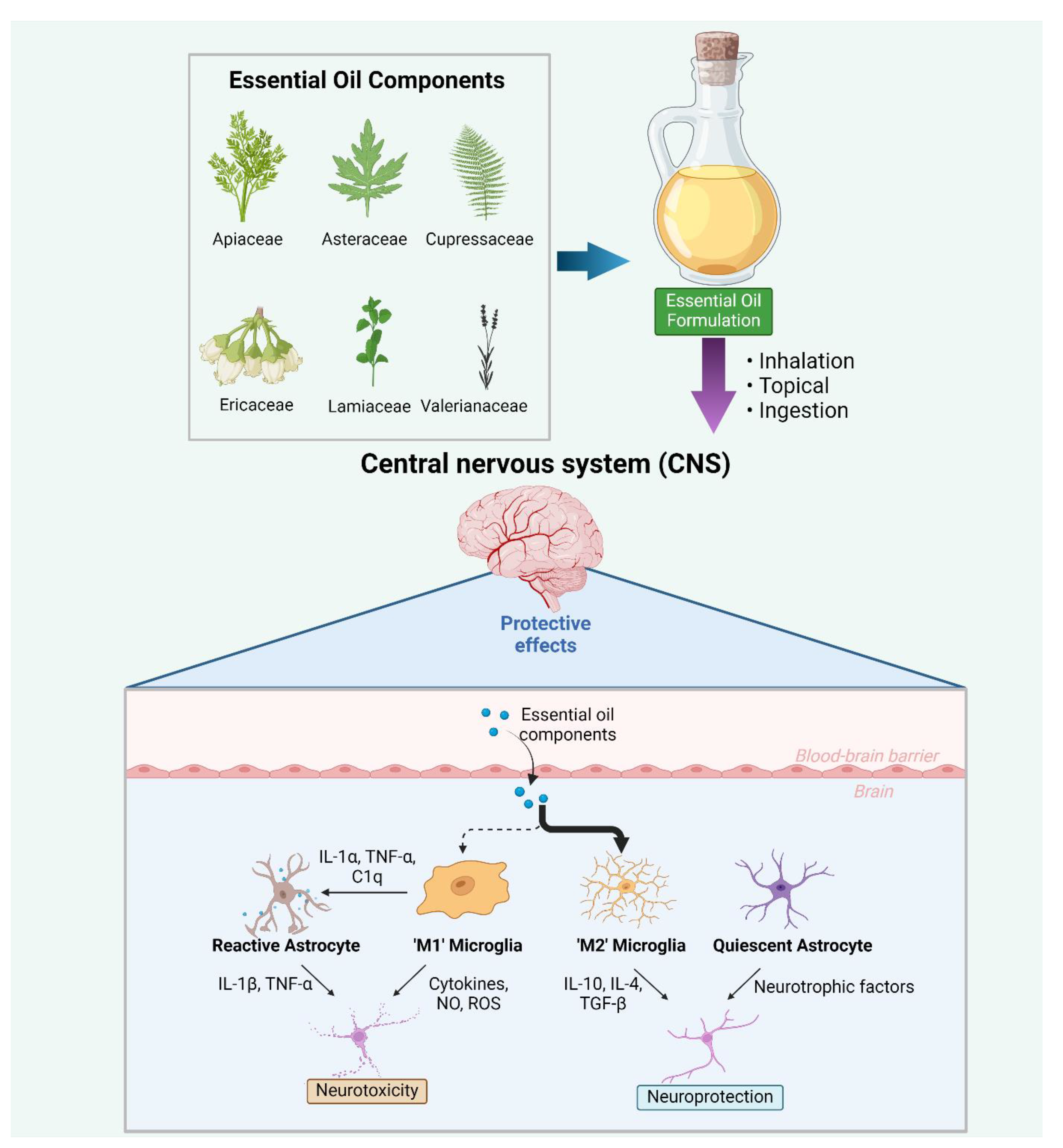
2. Signalling Pathways Associated with Microglia
3. The Potential of Essential Oils to Modulate Microglia and Microglial-Associated Inflammation
4. Key Essential Oils of Interest and Their Anti-Inflammatory Activity
4.1. Lamiaceae Family Associated EOs
4.2. Asteraceae Family Associated EOs
4.3. Fabaceae Family Associated EOs
4.4. Ericaceae Family Associated EOs
4.5. Cannabaceae Family Associated EOs
4.6. Oleaceae Family Associated EOs
4.7. EOs Associated with Plants Used in Chinese Traditional Medicine
4.8. EO Associated with Plants Used as Foodstuff

{kind=link}
{kind=link}
{kind=link}
| Essential Oil/Plant Part | Experimental Setting | Observed Effect | Mechanism of Action | References |
|---|---|---|---|---|
| Acmella oleracea/aerial parts/inflorescence | LPS-activated BV-2 microglia | Decreased iNOS, COX-2, IL-1β, and TNF-α mRNA expression. | NA | [25] |
| Artemsia dracunculus/aerial parts | LPS-activated BV-2 microglia | Increased catalase and superoxide dismutase. | NA | [24] |
| Artemisia herba-alba/aerial parts | LPS-activated BV-2 microglia | Decreased NO production. | NA | [23] |
| Cannabis sativa L./aerial parts/inflorescence | LPS-activated BV-2 microglia | Decreased NO, iNOS, COX-2, NLRP3, IL-6, IL-1β and TNF-α, increases IL-4 Reduced NF-kB translocation and phosphorylation of p38, did not affect Akt phosphorylation. | NF-κB p38 | [56] |
| Cinnamomum camphora L./leaves | LPS-activated BV-2 microglia | Decreased IL-6, IL-18, IL-1β, and TNF-α secretion and mRNA expression. | Affects amino acid metabolism (alanine and aspartate) and tricarboxylic acid cycle | [59] |
| Copaifera sp./oil resin | HMC3 microglial cells | Phosphorylation of pI3K/Akt/mTOR positive regulation of MAPK and JAK/STAT signalling. | pI3K/Akt/mTOR MPAK JAK/STAT | [54] |
| Daucus carota subsp. Maximus/aerial parts | LPS-activated N9 microglia | Decreased NO production. | NA | [73] |
| Jasminum grandiflorum L./flowers | LPS-activated BV-2 microglia | Decreased in NO, IL-1β, and TNF-α, decrease in ROS production. | NA | [57] |
| Pelargonium graveolens/whole plant | LPS-activated BV-2 microglia | Decreased NO production. | Decreases iNOS and COX-2 | [77] |
| Patrinia scabiosaefolia/whole plant | LPS-activated BV-2 microglia | Decreased in IL-1β and IL-6. | NA | [31] |
| Rhododendron albiflorum/flowers and leaves | LPS-activated C20 microglia | Inhibited Ca2+ signalling. | Inhibits Ca2+ mobilisation and influx | [27] |
| Schisandra chinensis/NA | LPS-activated BV-2 microglia | Decreased in NO, IL-1β, IL-6 and TNF-α concentration. | NF-κB MPAK p38/JNK/ERK | [33] |
| Thymus vulgaris/NA | LPS-activated BV-2 microglia | Decreased TNF-α and IL-6 mRNA expression. | NF-κB C/EBPβ | [37] |
| Thymus zygis L. subsp. sylvestris/aerial parts /inflorescence | LPS-activated BV-2 microglia | Decreased NO production. | NA | [28] |
| Thuja orientalis/aerial parts | LPS-activated BV-2 microglia | Decreased NO production. | NA | [26] |
| Valeriana jatamansi Jones Rhizoma et Radix/rhizome and root | LPS-activated primary mouse microglia | Decreased in IL-1β and IL-6 concentration and mRNA. | NA | [29] |
| Zingiber officinale/NA | LPS-activated BV-2 microglia | Decreased TNF-α and IL-1β, NO, MMP-9, PGE2. | TLR4, NF-κB, MAPK, HIF-1α | [36,65,67] |
| Ziziphora clinopodioides/aerial parts/inflorescence | Wild type BV-2 and BV-2 (Acox1−/−) | Decreased in catalase. | NA | [46] |
5. Proposed Anti-Inflammatory Mechanisms of Selected Essential Oil Constituents
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Butovsky, O.; Weiner, H.L. Microglial signatures and their role in health and disease. Nat. Rev. Neurosci. 2018, 19, 622–635. [Google Scholar] [CrossRef]
- Guo, S.; Wang, H.; Yin, Y. Microglia Polarization from M1 to M2 in Neurodegenerative Diseases. Front. Aging Neurosci. 2022, 14, 815347. [Google Scholar] [CrossRef]
- Cai, Y.; Liu, J.; Wang, B.; Sun, M.; Yang, H. Microglia in the Neuroinflammatory Pathogenesis of Alzheimer’s Disease and Related Therapeutic Targets. Front. Immunol. 2022, 13, 856376. [Google Scholar] [CrossRef]
- Cherry, J.D.; Olschowka, J.A.; O’Banion, M.K. Neuroinflammation and M2 microglia: The good, the bad, and the inflamed. J. Neuroinflamm. 2014, 11, 98. [Google Scholar] [CrossRef]
- Leng, F.; Edison, P. Neuroinflammation and microglial activation in Alzheimer disease: Where do we go from here? Nat. Rev. Neurol. 2021, 17, 157–172. [Google Scholar] [CrossRef]
- Hansen, D.V.; Hanson, J.E.; Sheng, M. Microglia in Alzheimer’s disease. J. Cell Biol. 2018, 217, 459–472. [Google Scholar] [CrossRef]
- Spangenberg, E.E.; Lee, R.J.; Najafi, A.R.; Rice, R.A.; Elmore, M.R.; Blurton-Jones, M.; West, B.L.; Green, K.N. Eliminating microglia in Alzheimer’s mice prevents neuronal loss without modulating amyloid-β pathology. Brain J. Neurol. 2016, 139 Pt 4, 1265–1281. [Google Scholar] [CrossRef]
- Kenkhuis, B.; Somarakis, A.; de Haan, L.; Dzyubachyk, O.; IJsselsteijn, M.E.; de Miranda, N.; Lelieveldt, B.P.F.; Dijkstra, J.; van Roon-Mom, W.M.C.; Höllt, T.; et al. Iron loading is a prominent feature of activated microglia in Alzheimer’s disease patients. Acta Neuropathol. Commun. 2021, 9, 27. [Google Scholar] [CrossRef]
- Cougnoux, A.; Drummond, R.A.; Collar, A.L.; Iben, J.R.; Salman, A.; Westgarth, H.; Wassif, C.A.; Cawley, N.X.; Farhat, N.Y.; Ozato, K.; et al. Microglia activation in Niemann-Pick disease, type C1 is amendable to therapeutic intervention. Hum. Mol. Genet. 2018, 27, 2076–2089. [Google Scholar] [CrossRef]
- Kannan, S.; Dai, H.; Navath, R.S.; Balakrishnan, B.; Jyoti, A.; Janisse, J.; Romero, R.; Kannan, R.M. Dendrimer-based postnatal therapy for neuroinflammation and cerebral palsy in a rabbit model. Sci. Transl. Med. 2012, 4, 130ra46. [Google Scholar] [CrossRef]
- Kim, C.; Ho, D.H.; Suk, J.E.; You, S.; Michael, S.; Kang, J.; Joong Lee, S.; Masliah, E.; Hwang, D.; Lee, H.J.; et al. Neuron-released oligomeric α-synuclein is an endogenous agonist of TLR2 for paracrine activation of microglia. Nat. Commun. 2013, 4, 1562. [Google Scholar] [CrossRef] [PubMed]
- Boza-Serrano, A.; Yang, Y.; Paulus, A.; Deierborg, T. Innate immune alterations are elicited in microglial cells before plaque deposition in the Alzheimer’s disease mouse model 5xFAD. Sci. Rep. 2018, 8, 1550. [Google Scholar] [CrossRef] [PubMed]
- Bai, D.; Li, X.; Wang, S.; Zhang, T.; Wei, Y.; Wang, Q.; Dong, W.; Song, J.; Gao, P.; Li, Y.; et al. Advances in extraction methods, chemical constituents, pharmacological activities, molecular targets and toxicology of volatile oil from Acorus calamus var. angustatus Besser. Front. Pharmacol. 2022, 13, 1004529. [Google Scholar] [CrossRef]
- Jain, M.; Singh, M.K.; Shyam, H.; Mishra, A.; Kumar, S.; Kumar, A.; Kushwaha, J. Role of JAK/STAT in the neuroinflammation and its association with neurological disorders. Ann. Neurosci. 2021, 28, 191–200. [Google Scholar] [CrossRef]
- Bachstetter, A.D.; Xing, B.; de Almeida, L.; Dimayuga, E.R.; Watterson, D.M.; Van Eldik, L.J. Microglial p38α MAPK is a key regulator of proinflammatory cytokine up-regulation induced by toll-like receptor (TLR) ligands or beta-amyloid (Aβ). J. Neuroinflamm. 2011, 8, 79. [Google Scholar] [CrossRef]
- da Fonseca, A.C.; Matias, D.; Garcia, C.; Amaral, R.; Geraldo, L.H.; Freitas, C.; Lima, F.R. The impact of microglial activation on blood-brain barrier in brain diseases. Front. Cell. Neurosci. 2014, 8, 362. [Google Scholar] [CrossRef]
- Apolloni, S.; Amadio, S.; Parisi, C.; Matteucci, A.; Potenza, R.L.; Armida, M.; Popoli, P.; D’Ambrosi, N.; Volonté, C. Spinal cord pathology is ameliorated by P2X7 antagonism in a SOD1-mutant mouse model of amyotrophic lateral sclerosis. Dis. Models Mech. 2014, 7, 1101–1109. [Google Scholar]
- Vay, S.U.; Flitsch, L.J.; Rabenstein, M.; Monière, H.; Jakovcevski, I.; Andjus, P.; Bijelic, D.; Blaschke, S.; Walter, H.L.; Fink, G.R.; et al. The impact of hyperpolarization-activated cyclic nucleotide-gated (HCN) and voltage-gated potassium KCNQ/Kv7 channels on primary microglia function. J. Neuroinflamm. 2020, 17, 100. [Google Scholar] [CrossRef]
- Loram, L.C.; Sholar, P.W.; Taylor, F.R.; Wiesler, J.L.; Babb, J.A.; Strand, K.A.; Berkelhammer, D.; Day, H.E.; Maier, S.F.; Watkins, L.R. Sex and estradiol influence glial pro-inflammatory responses to lipopolysaccharide in rats. Psychoneuroendocrinology 2012, 37, 1688–1699. [Google Scholar] [CrossRef]
- Radulović, N.S.; Blagojević, P.D.; Stojanović-Radić, Z.Z.; Stojanović, N.M. Antimicrobial plant metabolites: Structural diversity and mechanism of action. Curr. Med. Chem. 2013, 20, 932–952. [Google Scholar]
- Wang, Z.-J.; Heinbockel, T. Essential Oils and Their Constituents Targeting the GABAergic System and Sodium Channels as Treatment of Neurological Diseases. Molecules 2018, 23, 1061. [Google Scholar] [CrossRef] [PubMed]
- Alves-Silva, J.M.; Zuzarte, M.; Gonçalves, M.J.; Cavaleiro, C.; Cruz, M.T.; Cardoso, S.M.; Salgueiro, L. New Claims for Wild Carrot (Daucus carota subsp. carota) Essential Oil. Evid. Based Complement. Altern. Med 2016, 2016, 9045196. [Google Scholar] [CrossRef] [PubMed]
- Abu-Darwish, M.S.; Cabral, C.; Gonçalves, M.J.; Cavaleiro, C.; Cruz, M.T.; Efferth, T.; Salgueiro, L. Artemisia herba-alba essential oil from Buseirah (South Jordan): Chemical characterization and assessment of safe antifungal and anti-inflammatory doses. J. Ethnopharmacol. 2015, 174, 153–160. [Google Scholar] [CrossRef]
- Sahakyan, N.; Andreoletti, P.; Cherkaoui-Malki, M.; Petrosyan, M.; Trchounian, A. Artemisia dracunculus L. essential oil phytochemical components trigger the activity of cellular antioxidant enzymes. J. Food Biochem. 2021, 45, e13691. [Google Scholar] [CrossRef]
- Spinozzi, E.; Pavela, R.; Bonacucina, G.; Perinelli, D.R.; Cespi, M.; Petrelli, R.; Cappellacci, L.; Fiorini, D.; Scortichini, S.; Garzoli, S.; et al. Spilanthol-rich essential oil obtained by microwave-assisted extraction from Acmella oleracea (L.) R.K. Jansen and its nanoemulsion: Insecticidal, cytotoxic and anti-inflammatory activities. Ind. Crops Prod. 2021, 172, 114027. [Google Scholar] [CrossRef]
- Kim, K.H.; Moon, E.; Kim, S.Y.; Choi, S.U.; Son, M.W.; Choi, S.Z.; Lee, K.R. Bioactive sesquiterpenes from the essential oil of Thuja orientalis. Planta Medica 2013, 79, 1680–1684. [Google Scholar] [CrossRef]
- Schepetkin, I.A.; Özek, G.; Özek, T.; Kirpotina, L.N.; Khlebnikov, A.I.; Quinn, M.T. Chemical composition and immunomodulatory activity of essential oils from Rhododendron albiflorum. Molecules 2021, 26, 3652. [Google Scholar] [CrossRef] [PubMed]
- Rodrigues, V.; Cabral, C.; Evora, L.; Ferreira, I.; Cavaleiro, C.; Cruz, M.T.; Salgueiro, L. Chemical composition, anti-inflammatory activity and cytotoxicity of Thymus zygis L. subsp. sylvestris (Hoffmanns. & Link) Cout. essential oil and its main compounds. Arab. J. Chem. 2019, 12, 3236–3243. [Google Scholar]
- Yang, N.Y.; Li, Q.R.; Zhang, X.; Zeng, F.L.; Shen, X.C.; Long, Q.D. Spectrum-efficacy relationships between GC-MS fingerprints of essential oil from Valerianae Jatamansi Rhizoma et Radix and the efficacy of inhibiting microglial activation. Evid. Based Complement. Altern. Med. 2022, 2022, 9972902. [Google Scholar] [CrossRef]
- Alberti, T.B.; Marcon, R.; Bicca, M.A.; Raposo, N.R.; Calixto, J.B.; Dutra, R.C. Essential oil from Pterodon emarginatus seeds ameliorates experimental autoimmune encephalomyelitis by modulating Th1/Treg cell balance. J. Ethnopharmacol. 2014, 155, 485–494. [Google Scholar] [CrossRef]
- Lin, J.; Cai, Q.Y.; Xu, W.; Lin, J.M.; Peng, J. Chemical composition, anticancer, anti-neuroinflammatory, and antioxidant activities of the essential Oil of Patrinia scabiosaefolia. Chin. J. Integr. Med. 2018, 24, 207–212. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Pan, C.; Ou, Z.; Liang, X.; Shi, Y.; Chi, L.; Zhang, Z.; Zheng, X.; Li, C.; Xiang, H. Chemical profiling and bioactivity of essential oils from Alpinia officinarum Hance from ten localities in China. Ind. Crops Prod. 2020, 153, 112583. [Google Scholar] [CrossRef]
- Xu, M.; Zhang, X.; Ren, F.; Yan, T.; Wu, B.; Bi, K.; Bi, W.; Jia, Y. Essential oil of Schisandra chinensis ameliorates cognitive decline in mice by alleviating inflammation. Food Funct. 2019, 10, 5827–5842. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Ye, M.; Shi, Y.; Zhu, H.; Chi, L.; Pan, C.; Xu, Y.; Zheng, X.; Xiang, H.; Li, C. Phytochemical components and biological activities of essential oils from three selected medicinal plants. Ind. Crops Prod. 2021, 160, 113127. [Google Scholar] [CrossRef]
- Shi, A.; Long, Y.; Ma, Y.; Yu, S.; Li, D.; Deng, J.; Wen, J.; Li, X.; Wu, Y.; He, X.; et al. Natural essential oils derived from herbal medicines: A promising therapy strategy for treating cognitive impairment. Front. Aging Neurosci. 2023, 15, 1104269. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Chang, Y.Y.; Huang, S.; Xiao, L.H.; Zhou, W.; Zhang, L.Y.; Li, C.; Zhou, R.P.; Tang, J.; Lin, L.; et al. Aromatic-turmerone attenuates LPS-induced neuroinflammation and consequent memory impairment by targeting TLR4-dependent signaling pathway. Mol. Nutr. Food Res. 2018, 62, 1700281. [Google Scholar] [CrossRef] [PubMed]
- Horváth, G.; Horváth, A.; Reichert, G.; Böszörményi, A.; Sipos, K.; Pandur, E. Three chemotypes of thyme (Thymus vulgaris L.) essential oil and their main compounds affect differently the IL-6 and TNFα cytokine secretions of BV-2 microglia by modulating the NF-κB and C/EBPβ signalling pathways. BMC Complement. Med. Ther. 2021, 21, 148. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.L.; Wang, R.; Yakhnitsa, V.; Santos, J.M.; Watson, C.; Kiritoshi, T.; Ji, G.; Hamood, A.N.; Neugebauer, V. Gingerol-Enriched Ginger Supplementation Mitigates Neuropathic Pain via Mitigating Intestinal Permeability and Neuroinflammation: Gut-Brain Connection. Front. Pharmacol. 2022, 13, 912609. [Google Scholar] [CrossRef] [PubMed]
- Bhatt, N.; Joshi, N.; Ghai, K. Chemical composition and in-vitro antimicrobial activity of leaf essential oils of Senecio pedunculatus Edgew. and Nepeta coerulescens Maxim. J. Biol. Act. Prod. Nat. 2021, 11, 1–10. [Google Scholar] [CrossRef]
- Gobbo-Neto, L.; Lopes, N.P. Plantas medicinais: Fatores de influência no conteúdo de metabólitos secundários. Química Nova 2007, 30, 374–381. [Google Scholar] [CrossRef]
- Stahl-Biskup, E.; Venskutonis, R.P. 27—Thyme. In Handbook of Herbs and Spices, 2nd ed.; Peter, K.V., Ed.; Woodhead Publishing: Sawston, UK, 2012; pp. 499–525. [Google Scholar]
- Nikolić, M.; Glamočlija, J.; Ferreira, I.C.; Calhelha, R.C.; Fernandes, Â.; Marković, T.; Marković, D.; Giweli, A.; Soković, M. Chemical composition, antimicrobial, antioxidant and antitumor activity of Thymus serpyllum L., Thymus algeriensis Boiss. and Reut and Thymus vulgaris L. essential oils. Ind. Crops Prod. 2014, 52, 183–190. [Google Scholar] [CrossRef]
- Palmieri, S.; Pellegrini, M.; Ricci, A.; Compagnone, D.; Lo Sterzo, C. Chemical composition and antioxidant activity of thyme, hemp and coriander extracts: A comparison study of maceration, Soxhlet, UAE and RSLDE Techniques. Foods 2020, 9, 1221. [Google Scholar] [CrossRef] [PubMed]
- Agatonovic-Kustrin, S.; Kustrin, E.; Morton, D.W. Essential oils and functional herbs for healthy aging. Neural Regen. Res. 2019, 14, 441–445. [Google Scholar] [CrossRef] [PubMed]
- Coimbra, A.; Miguel, S.; Ribeiro, M.; Coutinho, P.; Silva, L.; Duarte, A.P.; Ferreira, S. Thymus zygis Essential Oil: Phytochemical Characterization, Bioactivity Evaluation and Synergistic Effect with Antibiotics against Staphylococcus aureus. Antibiotics 2022, 11, 146. [Google Scholar] [CrossRef] [PubMed]
- Sahakyan, N.; Petrosyan, M. Possible Preventive Effect of Ziziphora clinopodioides Lam. Essential Oil on Some Neurodegenerative Disorders. OBM Neurobiol. 2022, 6, 140. [Google Scholar] [CrossRef]
- Al-Musawi, M.; Ali, A.E.-M.; Humadi, A.; Al-kaisie, B. Antioxidant effects of 1,8-cineole against long term dl-polychlorinated biphenyls (PCBs) toxicity in domestic Hen’s Liver. Syst. Rev. Pharm. 2020, 11, 1150–1157. [Google Scholar]
- Raas, Q.; Saih, F.E.; Gondcaille, C.; Trompier, D.; Hamon, Y.; Leoni, V.; Caccia, C.; Nasser, B.; Jadot, M.; Ménétrier, F.; et al. A microglial cell model for acyl-CoA oxidase 1 deficiency. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 567–576. [Google Scholar] [CrossRef]
- Radulović, N.S.; Randjelović, P.J.; Stojanović, N.M.; Blagojević, P.D.; Stojanović-Radić, Z.Z.; Ilić, I.R.; Djordjević, V.B. Toxic essential oils. Part II: Chemical, toxicological, pharmacological and microbiological profiles of Artemisia annua L. volatiles. Food Chem. Toxicol. 2013, 58, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Ekiert, H.; Świątkowska, J.; Knut, E.; Klin, P.; Rzepiela, A.; Tomczyk, M.; Szopa, A. Artemisia dracunculus (Tarragon): A Review of Its Traditional Uses, Phytochemistry and Pharmacology. Front. Pharmacol. 2021, 12, 653993. [Google Scholar] [CrossRef]
- Abdul Rahim, R.; Jayusman, P.A.; Muhammad, N.; Mohamed, N.; Lim, V.; Ahmad, N.H.; Mohamad, S.; Abdul Hamid, Z.A.; Ahmad, F.; Mokhtar, N.; et al. Potential antioxidant and anti-inflammatory effects of Spilanthes acmella and its health beneficial effects: A Review. Int. J. Environ. Res. Public Health 2021, 18, 3532. [Google Scholar] [CrossRef]
- da Trindade, R.; da Silva, J.K.; Setzer, W.N. Copaifera of the neotropics: A review of the phytochemistry and pharmacology. Int. J. Mol. Sci. 2018, 19, 1511. [Google Scholar] [CrossRef] [PubMed]
- Arruda, C.; Aldana Mejia, J.A.; Ribeiro, V.P.; Gambeta Borges, C.H.; Martins, C.H.G.; Sola Veneziani, R.C.; Ambrosio, S.R.; Bastos, J.K. Occurrence, chemical composition, biological activities and analytical methods on Copaifera genus-A review. Biomed. Pharmacother. 2019, 109, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Urasaki, Y.; Beaumont, C.; Talbot, J.N.; Hill, D.K.; Le, T.T. Akt3 regulates the tissue-specific response to copaiba essential oil. Int. J. Mol. Sci. 2020, 21, 2851. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Biehl, A.; Gadina, M.; Hasni, S.; Schwartz, D.M. JAK-STAT Signaling as a target for inflammatory and autoimmune diseases: Current and future prospects. Drugs 2017, 77, 521–546. [Google Scholar] [CrossRef] [PubMed]
- Barbalace, M.C.; Freschi, M.; Rinaldi, I.; Mazzara, E.; Maraldi, T.; Malaguti, M.; Prata, C.; Maggi, F.; Petrelli, R.; Hrelia, S.; et al. Identification of Anti-Neuroinflammatory Bioactive Compounds in Essential Oils and Aqueous Distillation Residues Obtained from Commercial Varieties of Cannabis sativa L. Int. J. Mol. Sci. 2023, 24, 16601. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Zeng, X.; Feng, Y.; Li, S.; Wang, Y.; Liu, Y.; Chen, F.; Guan, Z.; Chen, T.; Wei, F. Inhibitory effects of Jasminum grandiflorum L. essential oil on lipopolysaccharide-induced microglia activation-integrated characteristic analysis of volatile compounds, network pharmacology, and BV-2 cell. Front. Pharmacol. 2023, 14, 1180618. [Google Scholar] [CrossRef] [PubMed]
- Matos, L.C.; Machado, J.P.; Monteiro, F.J.; Greten, H.J. Understanding traditional chinese medicine therapeutics: An overview of the basics and clinical applications. Healthcare 2021, 9, 257. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tang, C.; Zhou, Y.; Zhang, R.; Ye, S.; Zhao, Z.; Lin, L.; Yang, D. Anti-Inflammatory Property of the Essential Oil from Cinnamomum camphora (Linn.) Presl Leaves and the Evaluation of Its Underlying Mechanism by Using Metabolomics Analysis. Molecules 2020, 25, 4796. [Google Scholar] [CrossRef] [PubMed]
- Cho, E.-J.; Shin, J.-S.; Noh, Y.-S.; Cho, Y.-W.; Hong, S.-J.; Park, J.-H.; Lee, J.Y.; Lee, J.-Y.; Lee, K.-T. Anti-inflammatory effects of methanol extract of Patrinia scabiosaefolia in mice with ulcerative colitis. J. Ethnopharmacol. 2011, 136, 428–435. [Google Scholar] [CrossRef]
- Guleria, S.; Kumar, A.; Tiku, A.K. Chemical composition and fungitoxic activity of essential oil of Thuja orientalis L. grown in the north-western Himalaya. Z. Naturforsch. C 2008, 63, 211–214. [Google Scholar] [CrossRef]
- Mao, Q.Q.; Xu, X.Y.; Cao, S.Y.; Gan, R.Y.; Corke, H.; Beta, T.; Li, H.B. Bioactive compounds and bioactivities of ginger (Zingiber officinale Roscoe). Foods 2019, 8, 185. [Google Scholar] [CrossRef] [PubMed]
- Mohd Sahardi, N.F.N.; Makpol, S. Ginger (Zingiber officinale Roscoe) in the prevention of ageing and degenerative diseases: Review of current evidence. Evid. Based Complement. Altern. Med. 2019, 2019, 5054395. [Google Scholar] [CrossRef]
- Huang, Z.; Yuan, Y.; Tan, Z.; Zheng, J.; Zhang, W.; Huang, S.; Wang, Y.; Chen, M.; Zhang, L.; Li, H. Metabolomics in combination with network pharmacology reveals the potential anti-neuroinflammatory mechanism of essential oils from four Curcuma species. Ind. Crops Prod. 2023, 195, 116411. [Google Scholar] [CrossRef]
- Park, S.Y.; Jin, M.L.; Kim, Y.H.; Kim, Y.; Lee, S.J. Anti-inflammatory effects of aromatic-turmerone through blocking of NF-κB, JNK, and p38 MAPK signaling pathways in amyloid β-stimulated microglia. Int. Immunopharmacol. 2012, 14, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Hao, S.; Cho, B.O.; Wang, F.; Shin, J.Y.; Shin, D.; Jang, S.I. Zingiber officinale attenuates neuroinflammation in LPS-stimulated mouse microglia by AKT/STAT3, MAPK, and NF-κB signaling. Food Sci. Technol. 2022, 42, e104221. [Google Scholar] [CrossRef]
- Liu, B.G.; Mo, X.N.; Mo, X.H.; Zou, M.; Mei, X.A.; Chen, W.; Wu, L.; Wu, L. Gingerol ameliorates neuronal damage induced by hypoxia-reoxygenation via the miR-210/brain-derived neurotrophic factor axis. Kaohsiung J. Med. Sci. 2021, 38, 367–377. [Google Scholar]
- Chinese Pharmacopoeia. Chinese Pharmacopoeia, 9th ed.; China Medical Science and Technology Press: Beijing, China, 2015. [Google Scholar]
- Xu, Z.; Zhou, X.; Hong, X.; Wang, S.; Wei, J.; Huang, J.; Ji, L.; Yang, Y.; Efferth, T.; Hong, C.; et al. Essential oil of Acorus tatarinowii Schott inhibits neuroinflammation by suppressing NLRP3 inflammasome activation in 3 × Tg-AD transgenic mice. Phytomedicine 2023, 112, 154695. [Google Scholar] [CrossRef] [PubMed]
- Ullah, H.; Di Minno, A.; Santarcangelo, C.; Khan, H.; Daglia, M. Improvement of Oxidative Stress and Mitochondrial Dysfunction by β-Caryophyllene: A Focus on the Nervous System. Antioxidants 2021, 10, 546. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Li, Y.; Yin, R.; Guo, P.; Lei, N.; Li, G.; Xiong, L.; Xie, Y. Therapeutic potential of aromatic plant extracts in Alzheimer’s disease: Comprehensive review of their underlying mechanisms. CNS Neurosci. Ther. 2023, 29, 2045–2059. [Google Scholar] [CrossRef]
- He, X.; Wang, S.; Shi, J.; Sun, Z.; Lei, Z.; Yin, Z.; Qian, Z.; Tang, H.; Xie, H. Genotypic and environmental effects on the volatile chemotype of Valeriana jatamansi Jones. Front. Plant Sci. 2018, 9, 1003. [Google Scholar] [CrossRef]
- Valente, J.; Resende, R.; Zuzarte, M.J.; Gonçalves, M.; Lopes, M.C.; Cavaleiro, C.; Pereira, C.; Salgueiro, L.; Cruz, M.T. Bioactivity and safety profile of Daucus carota subsp. maximus essential oil. Ind. Crops Prod. 2015, 77, 218–224. [Google Scholar] [CrossRef]
- Batiha, G.E.; Shaheen, H.M.; Elhawary, E.A.; Mostafa, N.M.; Eldahshan, O.A.; Sabatier, J.M. Phytochemical constituents, folk medicinal uses, and biological activities of genus angelica: A review. Molecules 2022, 28, 267. [Google Scholar] [CrossRef] [PubMed]
- Ashokkumar, K.; Simal-Gandara, J.; Murugan, M.; Dhanya, M.K.; Pandian, A. Nutmeg (Myristica fragrans Houtt.) essential oil: A review on its composition, biological, and pharmacological activities. Phytother. Res. 2022, 36, 2839–2851. [Google Scholar] [CrossRef] [PubMed]
- Kumar, L.N.; Singh, J.; Sudhir, P.S. The phytochemical composition, biological effects and biotechnological approaches to the production of high-value essential oil from geranium. In Essential Oil Research; Springer: Cham, Switzerland, 2019; pp. 327–352. [Google Scholar]
- Elmann, A.; Mordechay, S.; Rindner, M.; Ravid, U. Anti-neuroinflammatory effects of geranium oil in microglial cells. J. Funct. Foods. 2010, 2, 17–22. [Google Scholar] [CrossRef]
- Linghu, K.G.; Wu, G.P.; Fu, L.Y.; Yang, H.; Li, H.Z.; Chen, Y.; Yu, H.; Tao, L.; Shen, X.C. 1,8-cineole ameliorates lps-induced vascular endothelium dysfunction in mice via PPAR-γ dependent regulation of NF-κB. Front. Pharmacol. 2019, 10, 178. [Google Scholar] [CrossRef]
- Fidyt, K.; Fiedorowicz, A.; Strządała, L.; Szumny, A. β-Caryophyllene and β-Caryophyllene oxide—Natural compounds of anticancer and analgesic properties. Cancer Med. 2016, 5, 3007–3017. [Google Scholar] [CrossRef]
- Viveros-Paredes, J.M.; González-Castañeda, R.E.; Gertsch, J.; Chaparro-Huerta, V.; López-Roa, R.I.; Vázquez-Valls, E.; Beas-Zarate, C.; Camins-Espuny, A.; Flores-Soto, M.E. Neuroprotective effects of β-caryophyllene against dopaminergic neuron injury in a murine model of parkinson’s disease induced by MPTP. Pharmaceuticals 2017, 10, 60. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Dong, Z.; Liu, S. β-caryophyllene ameliorates the alzheimer-like phenotype in APP/PS1 mice through CB2 receptor activation and the PPARγ pathway. Pharmacology 2014, 94, 1–12. [Google Scholar] [CrossRef]
- El Azab, E.F.; Abdulmalek, S. Amelioration of age-related multiple neuronal impairments and inflammation in high-fat diet-fed rats: The prospective multitargets of geraniol. Oxidative Med. Cell. Longev. 2022, 2022, 4812993. [Google Scholar] [CrossRef]
- Wang, J.; Su, B.; Zhu, H.; Chen, C.; Zhao, G. Protective effect of geraniol inhibits inflammatory response, oxidative stress and apoptosis in traumatic injury of the spinal cord through modulation of NF-κB and p38 MAPK. Exp. Ther. Med. 2016, 12, 3607–3613f. [Google Scholar] [CrossRef]
- Ben Ammar, R.; Mohamed, M.E.; Alfwuaires, M.; Abdulaziz Alamer, S.; Bani Ismail, M.; Veeraraghavan, V.P.; Sekar, A.K.; Ksouri, R.; Rajendran, P. Anti-inflammatory activity of geraniol isolated from lemon grass on Ox-LDL-stimulated endothelial cells by upregulation of Heme Oxygenase-1 via PI3K/Akt and Nrf-2 signaling pathways. Nutrients 2022, 14, 4817. [Google Scholar] [CrossRef]
- Du, J.; Liu, D.; Zhang, X.; Zhou, A.; Su, Y.; He, D.; Fu, S.; Gao, F. Menthol protects dopaminergic neurons against inflammation-mediated damage in lipopolysaccharide (LPS)-Evoked model of Parkinson’s disease. Int. Immunopharmacol. 2020, 85, 106679. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.J.; Yang, H.J.; Li, W.; Oh, Y.C.; Go, Y. Menthae Herba attenuates neuroinflammation by regulating CREB/Nrf2/HO-1 pathway in BV2 microglial cells. Antioxidants 2022, 11, 649. [Google Scholar] [CrossRef]
- Li, Y.; Lv, O.; Zhou, F.; Li, Q.; Wu, Z.; Zheng, Y. Linalool inhibits LPS-induced inflammation in BV2 microglia cells by activating Nrf2. Neurochem. Res. 2015, 40, 1520–1525. [Google Scholar] [CrossRef] [PubMed]
- Xu, F.; Na, L.; Li, Y.; Chen, L. Roles of the PI3K/AKT/mTOR signalling pathways in neurodegenerative diseases and tumours. Cell Biosci. 2020, 10, 54. [Google Scholar] [CrossRef] [PubMed]
- Kang, C.H.; Jayasooriya, R.G.; Choi, Y.H.; Moon, S.K.; Kim, W.J.; Kim, G.Y. β-Ionone attenuates LPS-induced pro-inflammatory mediators such as NO, PGE2 and TNF-α in BV2 microglial cells via suppression of the NF-κB and MAPK pathway. Toxicol. In Vitro 2013, 27, 782–787. [Google Scholar] [CrossRef]
- Chen, J.; Xie, X.; Li, M.; Xiong, Q.; Li, G.; Hang, H.; Chen, G.; Xu, X.; Yin, Y.; Peng, F.; et al. Pharmacological activities and mechanisms of action of Pogostemon cablin Benth: A review. Chin. Med. 2021, 16, 5. [Google Scholar]
- Nagoor Meeran, M.F.; Javed, H.; Al Taee, H.; Azimullah, S.; Ojha, S.K. Pharmacological properties and molecular mechanisms of thymol: Prospects for its therapeutic potential and pharmaceutical development. Front. Pharmacol. 2017, 8, 380. [Google Scholar] [CrossRef]
- Javed, H.; Azimullah, S.; Meeran, M.F.N.; Ansari, S.A.; Ojha, S. Neuroprotective Effects of thymol, a dietary monoterpene against dopaminergic neurodegeneration in rotenone-induced rat model of Parkinson’s disease. Int. J. Mol. Sci. 2019, 20, 1538. [Google Scholar] [CrossRef]
- Capatina, L.; Todirascu-Ciornea, E.; Napoli, E.M.; Ruberto, G.; Hritcu, L.; Dumitru, G. Thymus vulgaris essential oil protects zebrafish against cognitive dysfunction by regulating cholinergic and antioxidants systems. Antioxidants 2020, 9, 1083. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stojanović, N.M.; Ranđelović, P.J.; Simonović, M.; Radić, M.; Todorović, S.; Corrigan, M.; Harkin, A.; Boylan, F. Essential Oil Constituents as Anti-Inflammatory and Neuroprotective Agents: An Insight through Microglia Modulation. Int. J. Mol. Sci. 2024, 25, 5168. https://doi.org/10.3390/ijms25105168
Stojanović NM, Ranđelović PJ, Simonović M, Radić M, Todorović S, Corrigan M, Harkin A, Boylan F. Essential Oil Constituents as Anti-Inflammatory and Neuroprotective Agents: An Insight through Microglia Modulation. International Journal of Molecular Sciences. 2024; 25(10):5168. https://doi.org/10.3390/ijms25105168
Chicago/Turabian StyleStojanović, Nikola M., Pavle J. Ranđelović, Maja Simonović, Milica Radić, Stefan Todorović, Myles Corrigan, Andrew Harkin, and Fabio Boylan. 2024. "Essential Oil Constituents as Anti-Inflammatory and Neuroprotective Agents: An Insight through Microglia Modulation" International Journal of Molecular Sciences 25, no. 10: 5168. https://doi.org/10.3390/ijms25105168
APA StyleStojanović, N. M., Ranđelović, P. J., Simonović, M., Radić, M., Todorović, S., Corrigan, M., Harkin, A., & Boylan, F. (2024). Essential Oil Constituents as Anti-Inflammatory and Neuroprotective Agents: An Insight through Microglia Modulation. International Journal of Molecular Sciences, 25(10), 5168. https://doi.org/10.3390/ijms25105168