Abstract
Carotenoid cleavage oxygenases can cleave carotenoids into a range of biologically important products. Carotenoid isomerooxygenase (NinaB) and β, β-carotene 15, 15′-monooxygenase (BCO1) are two important oxygenases. In order to understand the roles that both oxygenases exert in crustaceans, we first investigated NinaB-like (EsNinaBl) and BCO1-like (EsBCO1l) within the genome of Chinese mitten crab (Eriocheir sinensis). Their functions were then deciphered through an analysis of their expression patterns, an in vitro β-carotene degradation assay, and RNA interference. The results showed that both EsNinaBl and EsBCO1l contain an RPE65 domain and exhibit high levels of expression in the hepatopancreas. During the molting stage, EsNinaBl exhibited significant upregulation in stage C, whereas EsBCO1l showed significantly higher expression levels at stage AB. Moreover, dietary supplementation with β-carotene resulted in a notable increase in the expression of EsNinaBl and EsBCO1l in the hepatopancreas. Further functional assays showed that the EsNinaBl expressed in E. coli underwent significant changes in its color, from orange to light; in addition, its β-carotene cleavage was higher than that of EsBCO1l. After the knockdown of EsNinaBl or EsBCO1l in juvenile E. sinensis, the expression levels of both genes were significantly decreased in the hepatopancreas, accompanied by a notable increase in the redness (a*) values. Furthermore, a significant increase in the β-carotene content was observed in the hepatopancreas when EsNinaBl-mRNA was suppressed, which suggests that EsNinaBl plays an important role in carotenoid cleavage, specifically β-carotene. In conclusion, our findings suggest that EsNinaBl and EsBCO1l may exhibit functional co-expression and play a crucial role in carotenoid cleavage in crabs.
1. Introduction
Most animals cannot biosynthesize carotenoids de novo and must obtain them from their diet. In animals, the carotenoid metabolism exerts several vital physiological roles; it exerts antioxidant and immune-enhancing functions and provides retinol precursors [1]. The major metabolic carotenoid conversions that take place in animals include oxidation, reduction, the translation of double bonds, the oxidative cleavage of double bonds, and the cleavage of epoxy bonds [2,3]. At present, research on the carotenoid metabolism in animals primarily focuses on the study of carotenoid cleavage oxygenases (CCOs) [4,5,6,7].
Carotenoid isomerooxygenase and β, β-carotene 15, 15′-monooxygenase (named BCO1 or BCMO), both members of the CCO family, are capable of cleaving various carotenoids into a variety of biologically significant products; this includes retinal, which occurs naturally in the food chain [8]. In mammals, carotenoid oxygenases and isomerases, which are encoded as two separate proteins, collaborate in the bioconversion pathway of retinols. Carotenoid oxygenase BCO1 oxidatively cleaves carotenoids into retinoids, and the retinoid isomerase RPE65 catalyzes the trans-to-cis isomerization of the C10–C11 double bond of retinoids [9,10,11,12,13]. Meanwhile, neither inactivation nor afterpotential mutant B (NinaB), a protein found in insect genomes, perform both functions via the catalysis of oxidative cleavage at the C15–C15′ double bond and trans-to-cis isomerization at the C11–C12 double bonds of carotenoid substrates [7]. NinaB is required by Drosophila for vision and can metabolize carotenoids into visual chromophores [14]. In addition, Chai et al. (2019) discovered that NinaB is involved in the reproduction of the silkworm Bombyx mori and the flour beetle Tribolium castaneum [15]. Furthermore, a gene named Carotenoid Oxygenases and Retinal Isomerase (BmCORI) has been identified in B. mori; this may play a pivotal role in the β-carotene metabolism [16]. To date, only one study has investigated the function of NinaB in crustaceans, and this was carried out using the ridgetail white prawn Exopalaemon carinicauda. This study revealed that two genes, namely carotenoid isomeroxygenase-like 1 and 2, are highly expressed in the eyestalk of E. carinicauda [17]. In pathogen challenge experiments on Vibrio parahaemolyticus and Aeromonas hydrophala, the knockout treatment of E. carinicauda was performed using EcNinaB-X1; this resulted in significantly lower mortality rates than the control, indicating that EcNinaB-X1 plays a role in immune defense [18].
BCO1, a cytosolic enzyme, cleaves β-carotene into retinal, thus playing an important role in retinoid synthesis [19]. The mutations that occur in this gene have been verified in some vertebrates, demonstrating their potential to influence the efficiency of carotenoid conversion and their content [20,21]. However, very few reports on the activities of BCO1 in aquatic animals could be found. For instance, bcox knockdown in zebrafish resulted in severe malformation and reduced pigmentation [12]. BCMO1-mRNA in the nematode Caenorhabditis elegans is localized to intestinal cells, which are the tissues used to detect retinal [22]. In mollusks, βCDOX was expressed in various tissues of the pearl oyster Pinctada fucata martensii, with the highest level of βCDOX found in the hepatopancreas [23]. Moreover, although the ability of PyBCO-like 1 to cleave β, β-carotene in yesso scallop Patinopecten yessoensis has been confirmed, the specific mechanism implicated in carotenoid cleavage in the adductor muscle remains unknown [24]. As insect genomes only contain NinaB as a carotenoid oxygenase, there are few studies on the expression and function of BCO in arthropods. However, both BCO and NinaB genes have been identified in the genome of the ridgetail white prawn E. carinicauda, and six BCOs were found to be significantly expressed in its stomach and hepatopancreas; this suggests that carotenoid cracking may occur in these two tissues [17].
Although previous studies have highlighted the crucial roles exerted by carotenoid oxygenases and isomerases in the carotenoid metabolism of animals, their functions remain largely unclear in crustaceans. Chinese mitten crab E. sinensis is an important economic crustacean; it is particularly popular due to its delicate flavor, red carapace, and deep yellow hepatopancreas [25,26,27]. With regard to the edible tissues of E. sinensis, its hepatopancreas and ovaries are rich in carotenoids [28]. However, very limited information about the CCOs present in this species is available. A recent study identified six CCO genes from the E. sinensis genome, and their expression pattern and evolution were further investigated [29]. Among these genes, one EsBCO1-like gene (EsBCO1l) and one EsNinaB-like gene (EsNinaBl) were screened and validated based on bioinformatics. However, the exact functions exerted by NinaB and BCO1 during the carotenoid catabolism of E. sinensis remain unclear.
In this study, the expression patterns of EsNinaBl and EsBCO1l were first investigated, and their recombinant proteins were then obtained from β-carotene-producing Escherichia coli; this was followed by biochemical characterization. Furthermore, the knockdown of EsNinaBl and EsBCO1l was conducted to further elucidate their functions during carotenoid cleavage. Moreover, the effects of dietary β-carotene supplementation on the gene expression levels of the hepatopancreas were further investigated for EsNinaBl and EsBCO1l. This study explores, for the first time, the differences between these two carotenoid cleavage enzymes, thus filling a gap in the research on carotenoid cleavage in crabs and contributing to revealing the carotenoid metabolism in crustaceans.
2. Results
2.1. Sequence Characterization and Phylogenetic Analysis of NinaB and BCO1
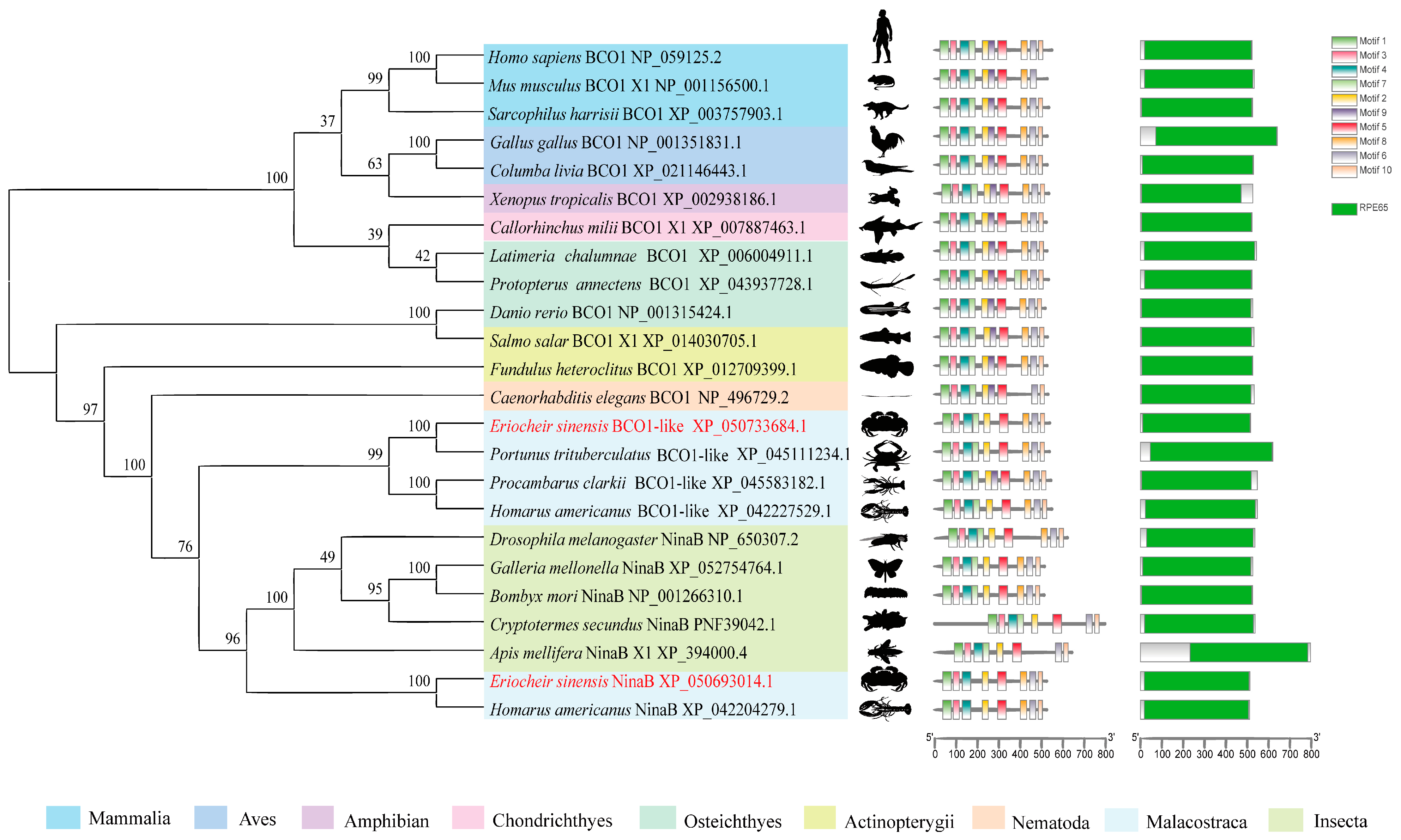
The multiple sequence alignment analysis revealed that EsNinaB shared a remarkably high degree of similarity with the NinaB obtained from closely related species (Figure S1). The phylogenetic analysis showed that the NinaB of E. sinensis formed a branch with Homarus americanus and that they constituted a cluster with the NinaB present in other insects (Figure 1). Simultaneously, the analysis highlighted that the BCO1 of E. sinensis, together with other crustacean BCO1s, formed a separate cluster that was closely associated with NinaB; thus, a supercluster with a bootstrap support of 76% was formed. Furthermore, the conservation motif analysis unveiled the presence of motifs 1–9 in all vertebrates, while insecta and malacostraca (excluding Procambarus clarkii) lacked motif 9. Additionally, compared to the NinaB in insecta and BCO1 in malacostraca, the NinaB in malacostraca lacked motif 7. The domain analysis revealed that a RPE65 domain exists in both BCO1 and NinaB across all species.
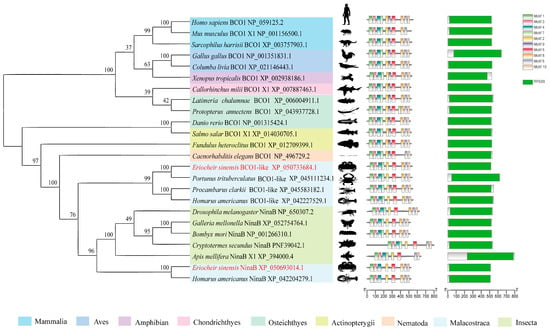
Figure 1.
Alignment of the phylogenetic tree of NinaB and BCO1. Values on the line of the phylogenetic tree are bootstrap values showing the percentage confidence in relatedness.
2.2. Gene Expression Patterns of EsNinaBl and EsBCO1l
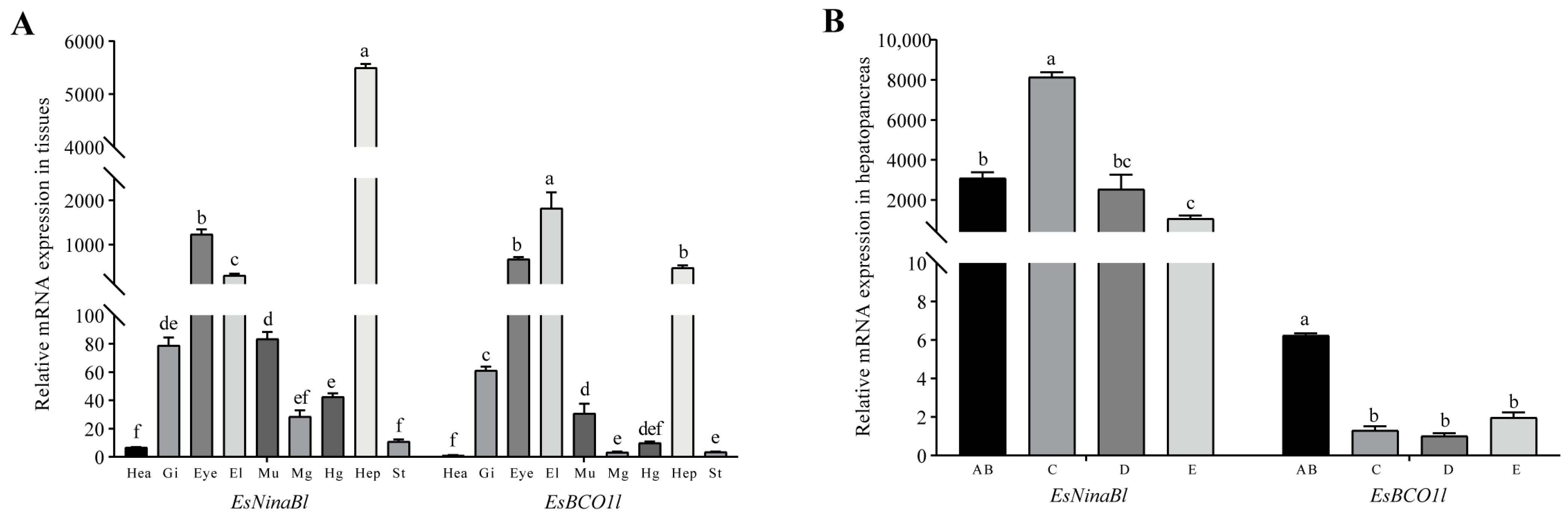
The gene expression patterns of EsNinaBl and EsBCO1l were analyzed in different tissues of E. sinensis and at different molting stages. The results showed that EsNinaBl was strongly expressed in the hepatopancreas (p < 0.05) and that EsBCO1l exhibited the highest expression in the epidermal layer; this was followed by the eyestalk and hepatopancreas (p < 0.05). Both genes were also expressed in other tissues (Figure 2A). Moreover, the expression of EsNinaBl increased during stage C (molting preparation period). Later, EsNinaBl was continuously expressed at a low level until the next molting period. As for EsBCO1l, its expression was highest during the AB stage (p < 0.05); this was followed by a significant decrease during stage C and a noticeable upward trend during stages D and E (Figure 2B).
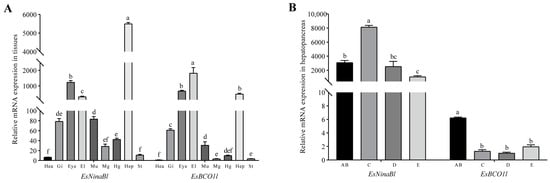
Figure 2.
Gene expression patterns of EsNinaBl and EsBCO1l in E. sinensis. (A) Gene expression levels of EsNinaBl and EsBCO1l in the different tissues of juvenile crabs. Hea: heart; Gi: gill; Eye: eyestalks; El: epidermal layer; Mu: muscle; Mg: midgut; Hg: hindgut; Hep: hepatopancreas; St: stomach. (B) The gene expression pattern of EsNinaBl and EsBCO1l in the hepatopancreas during the molting cycle of juvenile crabs. AB: postmolt; C: intermolt; D: premolt; E: ecdysis process. The amount of EsNinaBl and EsBCO1l mRNA was normalized to the β-actin transcript level. The data are shown as the means ± SEM (n = 5). The columns with different letters on the top indicate that they are significantly different for the same parameter (p < 0.05).
2.3. Recombinant EsBCO1l and EsNinaBl Could Cleave β-Carotene
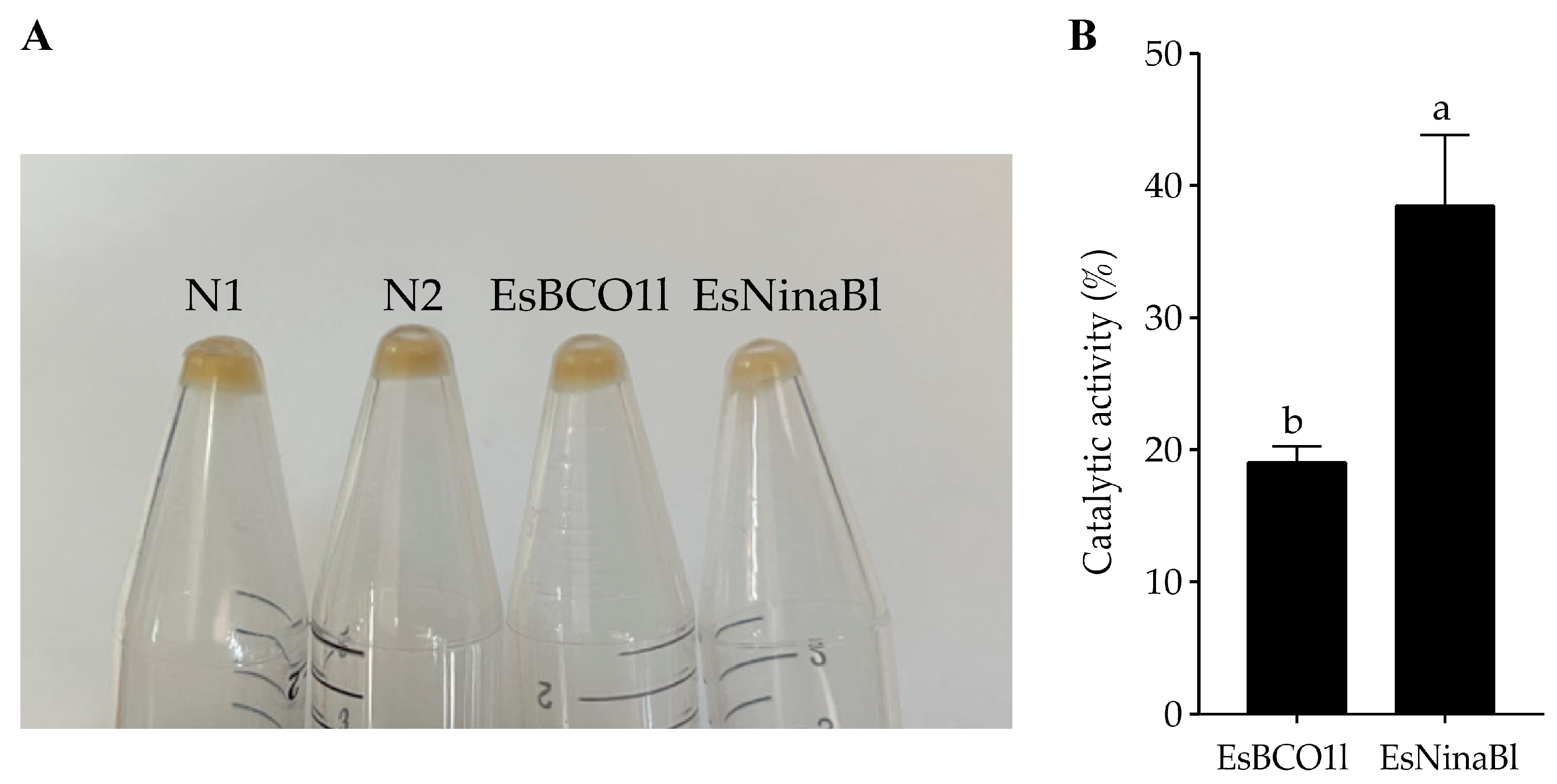
To further investigate the function of EsBCO1l and EsNinaBl in cleaving carotenoids, the recombinant EsBCO1l/EsNinaBl-pGEX-4T plasmid was transformed into E. coli strains, which accumulate β-carotene and exhibit a yellow color. In comparison to the E. coli controls that were transformed with the vector alone, the bacteria expressing EsBCO1l exhibited no significant change in color; meanwhile, evident decolorization was observed in the β-carotene-accumulated bacteria expressing EsNinaBl (Figure 3A). Furthermore, we investigated the ability of EsBCO1l and EsNinaBl to degrade β-carotene. It was evident that EsNinaBl exhibited a higher capacity for β-carotene degradation than EsBCO1l (Figure 3B).
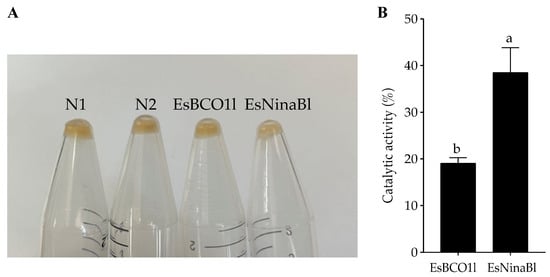
Figure 3.
Induced expression of pGEX-NinaBl and pGEX-BCO1l plasmids in E. coli strains engineered to accumulate β-carotene. (A) N1: pellet from the pGEX-4T vector in E. coli without expression; N2: pellet from the pGEX-4T vector with expressed E. coli; EsBCO1l: pellet from β-carotene-producing and pGEX-4T-EsBCO1l-expressed E. coli; EsNinaBl: pellet from β-carotene-producing and pGEX-4T-EsNinaBl-expressed E. coli. (B) Comparison of the degradation capacity of β-carotene among EsBCO1l and EsNinaBl. The columns with different letters on the top indicate that they are significantly different for the same parameter (p < 0.05).
2.4. Knockdown of EsNinaBl and EsBCO1l Increases β-Carotene Deposition in the Hepatopancreas
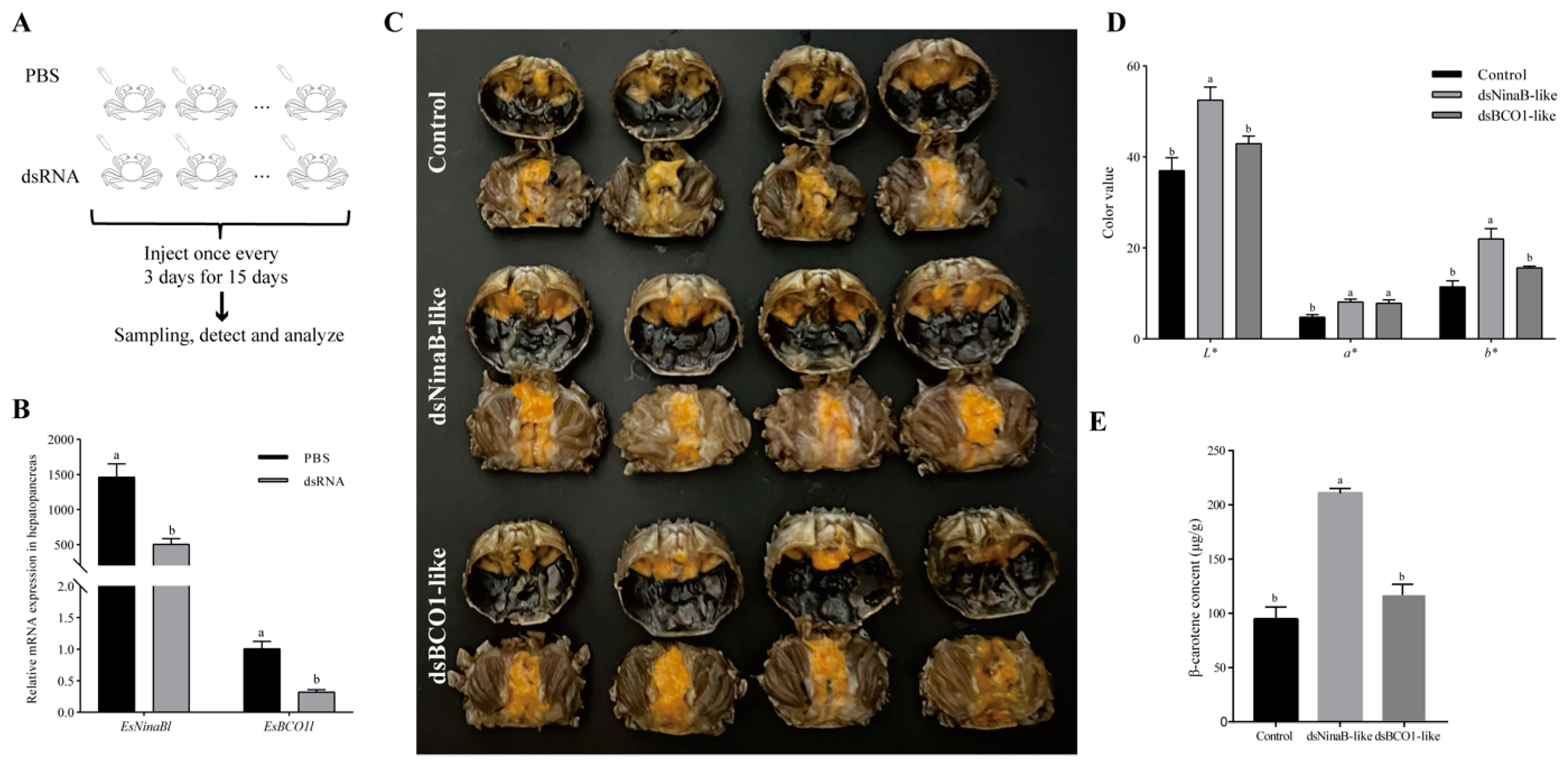
It was observed that, after 15 days of interference, there was a significant reduction in the expression of EsNinaBl upon EsNinaBl knockdown (p < 0.05). Likewise, a parallel occurrence was observed in the dsEsBCO1-like group (Figure 4B). Moreover, for the crabs injected with the dsRNA of EsNinaBl and EsBCO1l, the colors of the hepatopancreas changed significantly compared to those of the control group (Figure 4C). Figure 4D shows the color parameters of the hepatopancreas of juvenile crabs after dsEsNinaB-like and dsEsBCO1-like injections. The juveniles administered dsEsNinaB-like injections had significantly higher light (L*), red (a*), and yellow (b*) values than the controls (p < 0.05). Meanwhile, the color values of the dsEsBCO1-like group only exhibited a significant difference in their a* values compared to the control (p < 0.05). Notably, the L* and b* values of the dsEsNinaB-like group were significantly higher than those of the dsEsBCO1-like group (p < 0.05). Considering the apparent change in the color parameters of the hepatopancreas in the dsEsNinaB-like group, we determined their carotenoid content using HPLC analysis. There was a twofold increase in β-carotene in the dsEsNinaB-like group compared to the control and dsEsBCO1-like groups (Figure 4E).
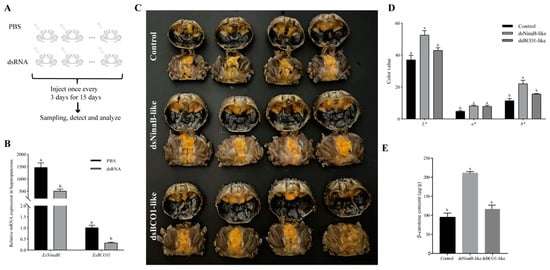
Figure 4.
Effects of dsRNA injection on gene expression (B), appearance characteristics (C), color parameters (D), and β-carotene contents (E) in the hepatopancreas of E. sinensis. (A) The diagram of this RNA interference experiment. The data are shown as the means ± SEM (n = 5). The columns with different letters on the top indicate that they are significantly different for the same parameter (p < 0.05).
2.5. Carotenoid Intake Influences the Expression of NinaBl and BCO1l
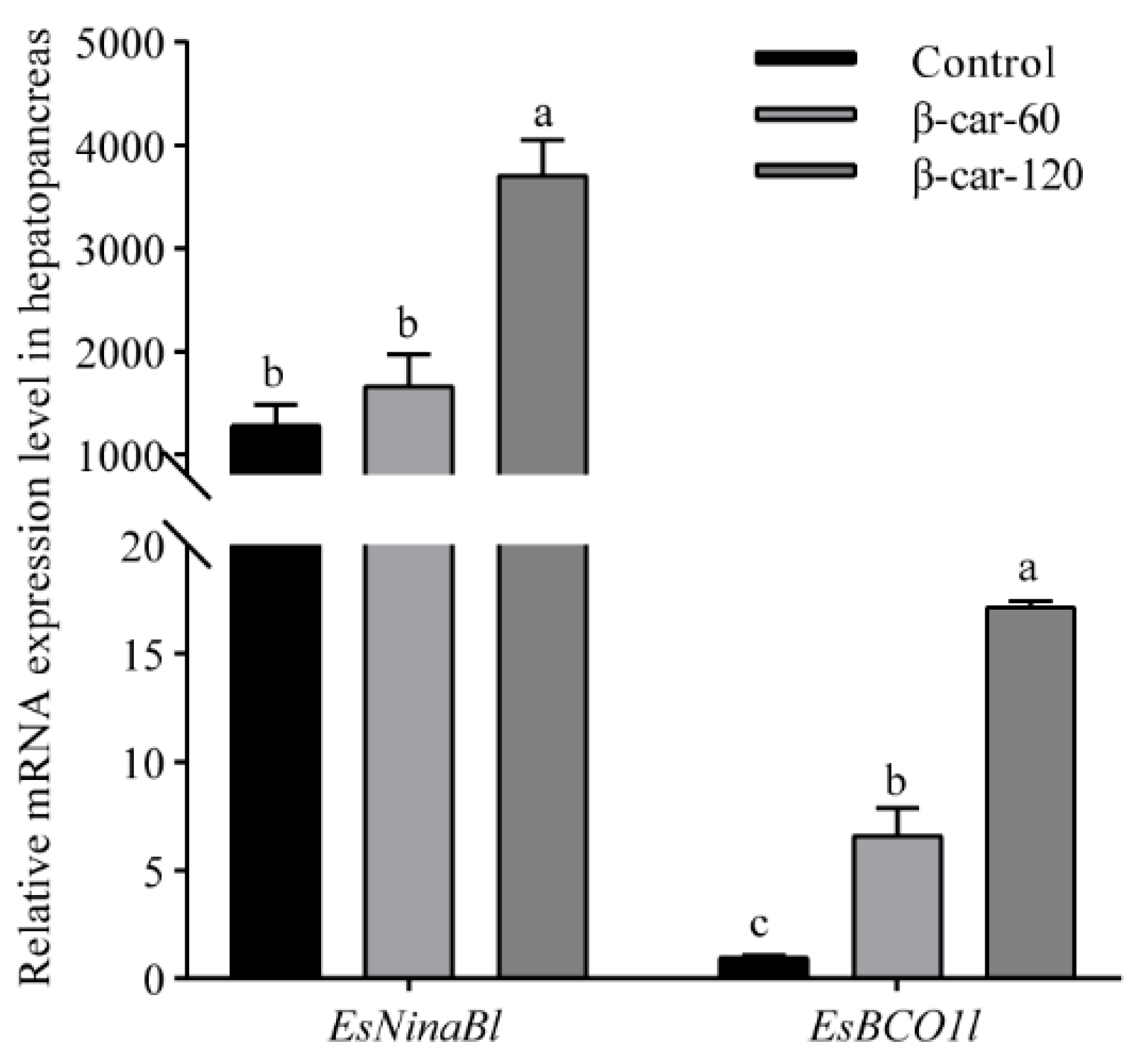
As shown in Figure 5, a significant increase in the mRNA expression of EsNinaBl and EsBCO1l with the increase in dietary β-carotene supplementation was observed. For EsNinaBl, the addition of 60 ppm of β-carotene did not result in a significant increase in its expression; meanwhile, there was a remarkable fivefold increase in the expression of EsBCO1l (p < 0.05). When the β-carotene supplementation reached 120 ppm, a substantial elevation in the expression of EsNinaBl was observed, showing a three- to fourfold increase compared to the control and β-car-60 groups. Similarly, the expression of EsBCO1l was upregulated significantly, showing values that were approximately 15 times higher than the control and around 2.5 times higher than the β-car-60 group.
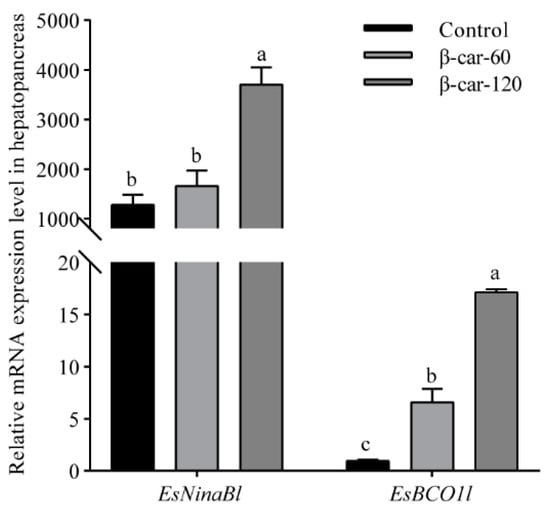
Figure 5.
Effects of dietary β-carotene supplementation on gene expression levels of EsNinaBl and EsBCO1l in the hepatopancreas of E. sinensis. The amount of EsNinaBl and EsBCO1l mRNA was normalized to the β-actin transcript level. The data are shown as the means ± SEM (n = 5). The columns with different letters on the top indicate that they are significantly different for the same parameter (p < 0.05).
3. Discussion
Crustaceans accumulate carotenoids through dietary intake, and carotenoid supplementation can provide significant health benefits, such as coloration and disease resistance [30]. Once carotenoids enter the body, they can be broken down into a series of crucial compounds, with carotenoid oxygenase and carotenoid isomerase playing essential roles in this process. BCO1 catalyzes the symmetrical cleavage of the C15–C15′ double bonds of carotenoids that exhibit provitamin A activity, such as β-carotene; it then converts them into colorless retinol [6,10]. NinaB, initially discovered in the fruit fly Drosophila melanogaster, catalyzes the trans–cis isomerization of the C11–C12 double bonds and the oxidative cleavage of the C15–C15′ double bonds in the main chain of carotenoids; visual chromophores are thus directly produced [7]. Research on the honeycomb moth Galleria mellonella has confirmed that NinaB possesses a dual function, performing both oxidative cleavage and isomerization [31]. It is noteworthy that both NinaB and BCO1 are capable of cleaving the C15–C15′ double bonds in the main chain of carotenoids; however, it remains unclear how they differ in this process. Both genes exhibit a typical RPE65 domain and belong to the CCO family. The evolutionary analysis showed that BCO1s and NinaBs are divided into two clades with high bootstrap support, indicating a relatively stable evolutionary relationship between them. In E. carinicauda, NinaB-X1 was also not classified as the same branch of BCO1, similar to the results of this study [18]. Compared to NinaB in insects and BCO1 in crustaceans, NinaB in crustaceans lacks motif 7, suggesting a potential evolutionary association. Furthermore, the structural domain analysis revealed a consistent, singular, and conserved RPE65 domain in both BCO1 and NinaB across all species. Similarly, a previous analysis of the CCO family in P. trituberculatus, E. sinensis, Penaeus chinensis, and Hyalella azteca indicated that these CCO genes have highly similar structures [29].
The activity and expression of carotenoid cleavage enzymes were found in several tissues of various vertebrate species [32]. However, research on these enzymes in crustaceans is currently limited. An investigation of the BCO genes in E. carinicauda revealed that BCO6 exhibits a high level of expression in the heart, while other BCO genes demonstrate elevated levels of expression in the hepatopancreas and stomach [17]. For EsBCO1l, we found that it exhibited high levels of expression in the epidermis layer, eyestalk, and hepatopancreas. Liu et al. found that the CCO3 (i.e., BCO1) gene was also highly expressed in the hepatopancreas of E. sinensis [29]. Moreover, EsNinaBl was highly expressed in the hepatopancreas in this study. These findings suggest that the hepatopancreas may be a crucial tissue in carotenoid digestion and cleavage in crustaceans. Additionally, studies on bivalves such as triangle sail mussel Hyriopsis ctumingii and pearl oyster Pinctada martensii have shown that BCO1 is predominantly expressed in the hepatopancreas, where the total carotenoid content is significantly higher than in other tissues [23,33]. This indicates that the hepatopancreas might be a crucial organ in carotenoid metabolism. Interestingly, in our study, EsBCO1l and EsNinaBl exhibited high expression levels in the eyestalk, which aligns with a previous study that reported the elevated expression of carotenoid isomerooxygenase-like 1 and 2 in the eyestalks of E. carinicauda [17]. In Drosophila melanogaster, NinaB has been identified as a key component in the production of visual pigments [31]. Therefore, we hypothesize that BCO1 and NinaB may have additional biological functions in other tissues.
Carotenoids play a critical role in the reproduction and growth of crustaceans [34,35]. Molting, a pivotal biological process in crustaceans, necessitates the substantial input of energy for water absorption and the swelling of the exoskeleton, which is intricately linked to both growth and gonad development [36,37]. The hepatopancreas serves as the primary energy source for the molting process, contributing to energy homeostasis and ensuring survival post-molting [36,38,39]. In comparison to other molting stages, the expression level of EsBCO1l was significantly higher in the AB stage; this may be due to the fact that the crabs had just completed molting and the new exoskeleton had not fully hardened; more carotenoid metabolites were thus required to promote growth [40]. Unlike EsBCO1l, stage C exhibited a substantial increase in the expression of EsNinaBl in the hepatopancreas. Although carotenoids are not directly required in stage C, we speculate that crustaceans undergoing exoskeleton shedding and the formation of a new exoskeleton may benefit from the antioxidant and immune-modulating functions of carotenoids [41]. With regard to the pathogen challenge of E. carinicauda, the roles of NinaB-X1 and BCO2 in immune defense have been confirmed; this is attributed to the accumulation of carotenoids [18,42]. In addition, our bioinformatic analysis revealed that EsBCO1l and EsNinaBl have RPE65 domains, which exert the oxidoreductase effects of a single donor with the incorporation of molecular oxygen or two atoms of oxygen [43]. Therefore, EsBCO1l and EsNinaBl in the hepatopancreas may be involved in immune function, helping to alleviate oxidative stress during the molting process via the accumulation of carotenoids. Consequently, both NinaB and BCO play pivotal roles in the vital physiological process of molting in crabs.
In this study, it was found that the hepatopancreas is the main tissue used to store and metabolize carotenoids [35]. Given that carotenoid cleavage oxygenase families (CCOs) are capable of cleaving carotenoids enzymatically [4], it is therefore reasonable to hypothesize that EsNinaBl and EsBCO1l, which belong to the CCO family, play a role in the carotenoid cleavage process that takes place in the hepatopancreas of crabs. In this study, we constructed the recombinant plasmids pGEX-EsNinaBl and pGEX-EsBCO1l in vitro. These plasmids were co-expressed by inserting them into E. coli, which can produce β-carotene. The noticeable decrease in yellow coloration and the reduction in the β-carotene content indicated that EsNinaBl possesses a stronger ability to cleave β-carotene compared to EsBCO1l. Similar findings were reported in a study on scallops, where BCO was found to cause a lightening of the color of E. coli-producing carotenoids [24].
To further investigate the potential effects of pGEX-EsNinaBl and pGEX-EsBCO1l, we conducted in vivo RNA interference on juvenile crabs. After 15 days of silencing NinaBl, a slight increase in L* and b* was observed in the dsNinaB-like group compared to the control. Similarly, the deletion of the NinaB-X1 and BCO genes in E. carinicauda resulted in a significant change in the hepatopancreas to a red color [18,42]. Additionally, in comparison to the dsBCO1-like group, the dsNinaB-like group showed significantly higher L* and b* values. This suggests that knocking down the NinaBl gene has a more pronounced impact on the coloration of the hepatopancreas in crabs compared to BCO1l. This finding was consistent with the in vitro results. Moreover, we observed a substantial increase in the β-carotene content in the hepatopancreas tissues of the dsNinaB-like experimental group; these results were approximately double those of the control group and the dsBCO1-like group. Earlier studies reported that knocking down BCO-like 6 in the hepatopancreas tissues of E. carinicauda not only induced a change in coloration but also led to a fivefold increase in the β-carotene content [17]. Therefore, the integration of in vitro and in vivo experiments indicates that NinaB exhibits a more pronounced ability to cleave β-carotene.
Numerous studies have demonstrated that the carotenoid content and color of crustaceans are enhanced significantly when they are fed diets with carotenoid supplementation [30,44,45]. In animals, dietary β-carotene serves as the substrate for the production of vitamin A, which is then converted into the chromophore [46]. The critical step in the conversion of β-carotene to vitamin A is the centric cleavage performed by BCO; in Drosophila, this is encoded by the ninaB gene [14]. Therefore, we conducted a β-carotene feeding experiment to investigate whether different intakes of dietary β-carotene would affect the expression of EsNinaBl and EsBCO1l in vivo. The results revealed a significant increase in the expression levels of EsNinaBl and EsBCO1l with the escalating addition of β-carotene. In particular, there was a notable increase in the expression level of EsBCO1l in β-car-120, which was almost 15 times greater than that of the control. We proposed that crabs experience the negative feedback regulation of retinoid homeostasis, in which nuclear receptor transcription factors regulate the gene expressions of carotenoid cleavage oxygenases to control the conversion of carotenoids and the production of retinoids [47]. This regulation may prevent the excessive production of vitamin A when the supply of provitamin A carotenoids is abundant and promote its production in deficient conditions. BCO1, a key enzyme involved in the formation of vitamin A, catalyzes the centric cleavage of β-carotene to yield retinaldehyde [48,49]. To sum up, it is reasonable to speculate that EsNinaB and EsBCO1 are functionally co-expressed; the two gene products may collaborate primarily in crab hepatopancreas to perform carotenoid cleavage.
4. Materials and Methods
4.1. Animals
Juvenile and adult crabs were sourced from the outer ponds of Chongming Research Station at Shanghai Ocean University, China, and maintained in an indoor facility with a circulating freshwater system. The crabs were fed daily; residual feed and feces were removed nightly. Hepatopancreas tissues were harvested at various molting stages, including AB (postmolt), C (intermolt), D (premolt), and E (ecdysis). Additionally, tissues from the gills, eyestalks, muscles, midgut, hindgut, stomach, and heart were collected from adult crabs and preserved at −80 °C for subsequent comparative expression analyses. All sampling procedures adhered to the ethical guidelines established by the Committee on Experimental Animal Management at Shanghai Ocean University. The approval code is SHOU-DW-2018-033, and the approval date is 26 January 2018.
4.2. RNA Extraction and cDNA Synthesis
The total RNA was extracted from the hepatopancreas of the crabs using RNAex Pro Reagent (AGBio, Changsha, China) following the manufacturer’s protocol. The quality of RNA was assessed using NanoDrop 2000 (Thermo Fisher Scientific, Waltham, MA, USA) and 1% agarose gel electrophoresis. Subsequently, cDNA was synthesized from the extracted RNA using the PrimeScript RT Reagent Kit (AGBio, Changsha, China), adhering to the manufacturer’s instructions.
4.3. Gene Bioinformatic Analysis
Based on the genome data of Eriocheir sinensis (accession no. ASM2467909), we identified the coding sequences (CDS) for NinaB (XM_050837057.1) and BCO1 (XM_050877727.1). We subsequently confirmed the full-length cDNA sequences of NinaB-like (EsNinaBl) and BCO1-like (EsBCO1l) in E. sinensis. The protein domains were predicted using Interpro (https://www.ebi.ac.uk/interpro/, accessed on 22 February 2024), and a motif analysis was performed using the MEME Suite (https://memesuite.org/meme/doc/meme.html, accessed on 22 February 2024). Multiple sequence alignments were carried out using MEGA11.0, and phylogenetic trees were constructed using the Maximum Likelihood method, with the visualization provided by TBtools-II software version 2.069.
4.4. In vivo Verification of EsNinaBl and EsBCO1l Functions
Gene fragments of EsNinaBl/EsBCO1l were inserted into the L4440 vector to construct interference vectors. These recombinant plasmids were transformed into HT115 competent cells (MKBio, Shanghai, China) and induced with IPTG to produce dsRNA that would target the specific fragments. The dsRNA was purified and diluted to a concentration of 1 μg/μL using nuclease-free 1 × PBS.
Juvenile crabs weighing approximately 10 g and in molting stage C were subjected to the dsRNA-mediated gene silencing of EsNinaBl and EsBCO1l. As outlined in Section 4.1, the crabs were maintained in an indoor container with a circulating freshwater system and were fed daily. Each group contained three replicates, with 10 crabs in each. Each crab received 1 μL of dsRNA or 1 × PBS (as a negative control) per gram of body weight; this was injected into the base of the third walking leg every three days over a period of fifteen days. At the end of the experiment, the survival rates were 60% for the dsRNA group and 80% for the PBS group. Subsequently, hepatopancreas tissues were harvested in order to analyze the expression of EsNinaBl and EsBCO1l and to detect carotenoids.
4.5. In Vitro Verification of EsNinaBl and EsBCO1l Function in E. coli
The pAC-BETAipi plasmid (Miaolingbio, Wuhan, China), which contains β-carotene synthesis genes, was transformed into E. coli BL(DE3) cells (Tiangen, Beijing, China). The open reading frames (ORFs) of EsNinaBl and EsBCO1l were amplified and cloned into the pGEX-4T vector (Miaolingbio, Wuhan, China) using T4 DNA ligase (Sangon, Shanghai, China). These recombinant plasmids were then introduced into E. coli strains harboring the pAC-BETAipi plasmid. The colonies that co-expressed β-carotene synthesis genes along with EsBCO1l/EsNinaBl were induced with 1 mM of IPTG. The phenotypic changes in the E. coli colonies were assessed, and those with only the pAC-BETAipi plasmid, those with both pAC-BETAipi and the recombinant pGEX-4T-EsBCO1l/EsNinaBl plasmids, and those with the pGEX-4T vector alone were compared.
4.6. Carotenoid Feeding
Healthy crabs, averaging 100 g in weight, were sourced from Chongming Research Station, Shanghai Ocean University, China. The crabs were individually cultured in boxes with shelters within a freshwater recirculating system and acclimated for one week using a carotenoid-free commercial crab feed.
In the formal experiment, 21 crabs were cultured in controlled tanks at 24 °C and divided into three groups of seven: one control and two β-carotene diet groups. β-carotene, sourced from Zhejiang NHU Co., Ltd. (Shaoxing, China), was administered at concentrations of 60 ppm and 120 ppm by incorporating it into the diet. The control group received a base diet devoid of any carotenoids. The details of the diet are provided in Table 1. The crabs were fed between 1.5 and 3.0% of their body weight daily at 18:00, with waste and uneaten feed removed by the following morning. The 28-day feeding trial concluded with the random selection of five crabs from each group for the collection of hepatopancreas tissue, which was immediately frozen in liquid nitrogen and stored at −80 °C for analysis.

Table 1.
The formulation, proximate composition, and actual carotenoid concentration of experimental diets.
4.7. Quantitative PCR
The gene expression levels were quantified using CFX384TM Real-Time System (Bio-Rad, Hercules, CA, USA). The reactions utilized 1 μL of cDNA with the SYBR Green Premix Pro Taq HS qPCR Kit (AGBio, Wuhan, China) in a 10 μL total reaction volume; this comprised 5 μL of 2 × SYBR Green Pro Taq HS Premix, 0.2 μL of each primer, and 3.6 μL of nuclease-free water. All qRT-PCR analyses were conducted with five biological replicates, and each sample was run in triplicate. β-actin served as the reference gene for normalization. The primer sequences that were used are listed in Table 2. The relative gene expression was calculated using the 2−ΔΔCt method [50].
Table 2.
Information on the primers used in this study.
4.8. Carotenoid Extraction and HPLC Analysis
The total carotenoids were extracted from tissues using acetone and then filtered. The optical density (OD) of the extracts was measured using a UV–Visible spectrophotometer (T6 New Century, Beijing Purkinje General Instrument Co., Ltd., Beijing, China) to estimate the β-carotene levels. The β-carotene content was quantified using an Agilent 1260 high-performance liquid chromatography (HPLC) system (Agilent Technologies Inc., Santa Clara, CA, USA) following the methodology described by Peng et al. [51]. The Agilent 1260 HPLC system utilized a YMC™ Carotenoid C30 column (4.6 × 150 mm, 3 μm particle size, YMC Co., Ltd., Kyoto, Japan). The gradient mobile phases consisted of phase A (methyl alcohol/methyl tert-butyl ether/formic acid at a ratio of 3:2:0.01, v/v/v) and phase B (alcohol/triethylamine at a ratio of 100:0.04, v/v). Details of the procedures used to perform the carotenoid analysis in crab tissues can be found in the methodology described by Long et al. [28].
4.9. Statistical Analysis
Statistical analyses were performed using one-way analysis of variance (ANOVA), followed by Tukey’s test, with SPSS 22.0 software. The results were deemed statistically significant at p-values less than 0.05. Values are presented as the means ± standard error (SE). All graphs were generated using GraphPad Prism version 7.03.
5. Conclusions
To the best of our knowledge, this study is the first to explore the functions of NinaB and BCO1 during carotenoid cleavage using comprehensive approaches. Both EsNinaBl and EsBCO1l are highly expressed in the hepatopancreas, but they have different expression patterns during the molting cycle of E. sinensis. The assessment of the catalytic activity of the recombinant protein in E. coli showed that EsNinaBl had a stronger catalytic efficiency than EsBCO1l for carotenoid cleavage. The knockdown of EsNinaBl or EsBCO1l led to an increase in β-carotene deposition in the hepatopancreas of juvenile E. sinensis, and EsNinaBl exhibited a significantly higher ability to degrade β-carotene than EsBCO1l. Moreover, β-carotene feeding led to a significant increase in the expression of both genes in the hepatopancreas. Taken together, these findings suggest that NinaB and BCO1 collaborate during carotenoid cleavage in E. sinensis. The present results offer initial insights into the importance of NinaB and BCO in carotenoid cleavage in crabs and provide valuable information regarding the regulation of the carotenoid metabolism in crustaceans.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/ijms25115592/s1.
Author Contributions
X.W. and M.Z. contributed to the conception and design of the study. M.Z. performed all experiments, performed the statistical analysis, and wrote the original draft. J.X. validated the gene sequences, conducted the bioinformatics analysis, performed the quantitative PCR, and extracted the carotenoids. Z.Y. and B.Z. assisted in the RNA interference and carotenoid feeding experiments. Y.W. assisted in the carotenoid extraction. X.C. and X.W. revised the manuscript and the research platform of this study. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by a general project from the Natural Science Foundation of China (No. 31873041), a Key R&D Program (No. 2022YFD2400702) from the Ministry of Science and Technology of China, and an agriculture R&D project from the Shanghai Municipal Science and Technology Commission (No. 23N71900100). X.G.W. was supported by the Leading Talent Project (No. DYRC202100215) in Yellow River Delta from the Dongying Municipal Government of Shandong Province and the Academic Fellow program in the School of Biological Sciences, Universiti Sains Malaysia during this study.
Institutional Review Board Statement
All sampling procedures adhered to the ethical guidelines established by the Committee on Experimental Animal Management at Shanghai Ocean University. The approval code is SHOU-DW-2018-033, and the approval date is 26 January 2018.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data are contained within the article and Supplementary Materials.
Acknowledgments
The authors would like to thank Gaofeng Qiu for kindly providing us the L4440 vector, and Le Yin, Di Kong, Tao Peng, Ya’nan Gao, Yufan Wang for their contributions to the crab cultures and sample collection for this study. Special thanks are also extended to Zhiyan Wang and Saira for their grammar checks on the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Stahl, W.; Sies, H. Bioactivity and protective effects of natural carotenoids. Biochim. Biophys. Acta-Mol. Basis Dis. 2005, 1740, 101–107. [Google Scholar] [CrossRef] [PubMed]
- Šesták, Z.; Britton, G.; Liaaen-Jensen, S.; Pfander, H. Carotenoids. Handbook. Photosynthetica 2004, 42, 186. [Google Scholar] [CrossRef]
- Maoka, T. Carotenoids in marine animals. Mar. Drugs 2011, 9, 278–293. [Google Scholar] [CrossRef] [PubMed]
- Poliakov, E.; Soucy, J.; Gentleman, S.; Rogozin, I.B.; Redmond, T.M. Phylogenetic analysis of the metazoan carotenoid oxygenase superfamily: A new ancestral gene assemblage of BCO-like (BCOL) proteins. Sci. Rep. 2017, 7, 13192. [Google Scholar] [CrossRef] [PubMed]
- Giuliano, G.; Al-Babili, S.; von Lintig, J. Carotenoid oxygenases: Cleave it or leave it. Trends Plant Sci. 2003, 8, 145–149. [Google Scholar] [CrossRef] [PubMed]
- Amengual, J.; Widjaja-Adhi, M.A.K.; Rodriguez-Santiago, S.; Hessel, S.; Golczak, M.; Palczewski, K.; von Lintig, J. Two carotenoid oxygenases contribute to mammalian provitamin A metabolism. J. Biol. Chem. 2013, 288, 34081–34096. [Google Scholar] [CrossRef] [PubMed]
- Oberhauser, V.; Voolstra, O.; Bangert, A.; von Lintig, J.; Vogt, K. NinaB combines carotenoid oxygenase and retinoid isomerase activity in a single polypeptide. Proc. Natl. Acad. Sci. USA 2008, 105, 19000–19005. [Google Scholar] [CrossRef] [PubMed]
- von Lintig, J.; Vogt, K. Vitamin A formation in animals: Molecular identification and functional characterization of carotene cleaving enzymes. J. Nutr. 2004, 134, 251–256. [Google Scholar] [CrossRef] [PubMed]
- Redmond, T.M.; Gentleman, S.; Duncan, T.; Yu, S.; Wiggert, B.; Gantt, E.; Cunningham, F.X. Identification, expression, and substrate specificity of a mammalian β-carotene 15,15′-dioxygenase. J. Biol. Chem. 2001, 276, 6560–6565. [Google Scholar] [CrossRef]
- Hessel, S.; Eichinger, A.; Isken, A.; Amengual, J.; Hunzelmann, S.; Hoeller, U.; Elste, V.; Hunziker, W.; Goralczyk, R.; Oberhauser, V.; et al. CMO1 deficiency abolishes vitamin A production from β-carotene and alters lipid metabolism in mice. J. Biol. Chem. 2007, 282, 33553–33561. [Google Scholar] [CrossRef]
- Redmond, T.M.; Poliakov, E.; Yu, S.; Tsai, J.Y.; Lu, Z.J.; Gentleman, S. Mutation of key residues of RPE65 abolishes its enzymatic role as isomerohydrolase in the visual cycle. Proc. Natl. Acad. Sci. USA 2005, 102, 13658–13663. [Google Scholar] [CrossRef] [PubMed]
- Lampert, J.M.; Holzschuh, J.; Hessel, S.; Driever, W.; Vogt, K.; von Lintig, J. Provitamin A conversion to retinal via the β, β-carotene-15,15′-oxygenase (bcox) is essential for pattern formation and differentiation during zebrafish embryogenesis. Development 2003, 130, 2173–2186. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Li, S.; Moghrabi, W.N.; Philp, A.R.; Travis, G.H.; Jin, M.; Li, S.; Moghrabi, W.N.; Philp, A.R.; Travis, G.H. Mutational analysis to determine key residues essential for activity and membrane association of RPE65 isomerohydrolase. Algebr. Geom. Topol. 2006, 12, 805–827. [Google Scholar]
- von Lintig, J.; Dreher, A.; Kiefer, C.; Wernet, M.F.; Vogt, K. Analysis of the blind Drosophila mutant ninaB identifies the gene encoding the key enzyme for vitamin A formation in vivo. Proc. Natl. Acad. Sci. USA 2001, 98, 1130–1135. [Google Scholar] [CrossRef] [PubMed]
- Chai, C.L.; Xu, X.; Sun, W.Z.; Zhang, F.; Ye, C.; Ding, G.S.; Li, J.T.; Zhong, G.X.; Xiao, W.; Liu, B.B.; et al. Characterization of the novel role of NinaB orthologs from Bombyx mori and Tribolium castaneum. Insect Biochem. Mol. Biol. 2019, 109, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Dong, X.L.; Pan, C.X.; Zhang, M.J. A novel gene Bombyx mori carotenoid oxygenases and retinal isomerase (BmCORI) related to β-carotene depletion. Biochem. Genet. 2020, 58, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Yu, Y.; Zhang, C.S.; Li, S.H.; Zhang, X.J.; Li, F.H. Characterization and function analysis of the beta-carotene oxygenase-like genes in carotenoids metabolism of the ridgetail white prawn Exopalaemon carinicauda. Front. Physiol. 2020, 11, 745. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.Y.; Yan, C.C.; Liu, M.F.; Liu, Y.J.; Wang, W.Z.; Cheng, W.Z.; Yang, F.S.; Zhang, J.Q. CRISPR/Cas9-mediated deletion of one carotenoid isomerooxygenase gene (EcNinaB-X1) from Exopalaemon carinicauda. Fish Shellfish. Immunol. 2020, 97, 421–431. [Google Scholar] [CrossRef]
- During, A.; Albaugh, G.; Smith, J.C. Characterization of β-carotene 15,15′-dioxygenase activity in TC7 clone of human intestinal cell line Caco-2. Biochem. Biophys. Res. Commun. 1998, 249, 467–474. [Google Scholar] [CrossRef]
- Cai, X.; Conley, S.M.; Naash, M.I. RPE65: Role in the visual cycle, human retinal disease, and gene therapy. Ophthalmic Genet. 2009, 30, 57–62. [Google Scholar] [CrossRef]
- Marlhens, F.; Bareil, C.; Griffoin, J.M.; Zrenner, E.; Amalric, P.; Eliaou, C.; Liu, S.Y.; Harris, E.; Redmond, T.M.; Arnaud, B.; et al. Mutations in RPE65 cause Leber’s congenital amaurosis. Nat. Genet. 1997, 17, 139–141. [Google Scholar] [CrossRef]
- Wang, P.; Liu, B.; Zhang, D.L.; Belew, M.Y.; Tissenbaum, H.A.; Cheng, J.X. Imaging lipid metabolism in live Caenorhabditis elegans using fingerprint vibrations. Angew. Chem. -Int. Ed. 2014, 53, 11787–11792. [Google Scholar] [CrossRef] [PubMed]
- Lei, C.; Li, J.H.; Zheng, Z.; Du, X.D.; Deng, Y.W. Molecular cloning, expression pattern of β-carotene 15,15-dioxygenase gene and association analysis with total carotenoid content in pearl oyster Pinctada fucata martensii. Comp. Biochem. Physiol. B-Biochem. Mol. Biol. 2019, 229, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, S.Y.; Xun, X.G.; Zhang, M.R.; Wang, S.; Li, H.D.; Zhao, L.; Fu, Q.; Wang, H.Z.; Li, T.T.; et al. A carotenoid oxygenase is responsible for muscle coloration in scallop. Biochim. Biophys. Acta-Mol. Cell Biol. Lipids 2019, 1864, 966–975. [Google Scholar] [CrossRef] [PubMed]
- Yin, M.Y.; Fu, X.Y.; Wang, X. C, Key lipid molecules in hepatopancreas of Eriocheir sinensis: Identification and thermal oxidative degradation characteristics. J. Food Biochem. 2021, 45, e13734. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Tao, N.-P.; Wu, X.G.; Wang, X.C. Metabolomics of the hepatopancreas in Chinese mitten crabs (Eriocheir sinensis). Food Res. Int. 2022, 152, 110914. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.Q.; Zu, L.; Cheng, Y.X.; Wade, N.M.; Liu, J.G.; Wu, X.G. Carapace color affects carotenoid composition and nutritional quality of the Chinese mitten crab, Eriochier sinensis. LWT-Food Sci. Technol. 2020, 126, 109286. [Google Scholar] [CrossRef]
- Long, X.W.; Wu, X.G.; Zhao, L.; Liu, J.G.; Cheng, Y.X. Effects of dietary supplementation with Haematococcus pluvialis cell powder on coloration, ovarian development and antioxidation capacity of adult female Chinese mitten crab, Eriocheir sinensis. Aquaculture 2017, 473, 545–553. [Google Scholar] [CrossRef]
- Liu, Y.F.; Li, M.J.; Zhang, M.; Yang, Z.L.; Chen, X.W.; Wu, X.G. Evolution and expression analysis of carotenoid cleavage oxygenase gene family in Chinese mitten crab Eriocheir sinensis. Int. J. Biol. Macromol. 2024, 257, 128475. [Google Scholar] [CrossRef]
- Jiang, X.D.; Zu, L.; Wang, Z.Y.; Cheng, Y.X.; Yang, Y.H.; Wu, X.G. Micro-algal astaxanthin could improve the antioxidant capability, immunity and ammonia resistance of juvenile Chinese mitten crab, Eriocheir sinensis. Fish Shellfish. Immunol. 2020, 102, 499–510. [Google Scholar] [CrossRef]
- Voolstra, O.; Oberhauser, V.; Sumser, E.; Meyer, N.E.; Maguire, M.E.; Huber, A.; von Lintig, J. NinaB is essential for Drosophila vision but induces retinal degeneration in opsin-deficient photoreceptors. J. Biol. Chem. 2010, 285, 2130–2139. [Google Scholar] [CrossRef] [PubMed]
- Lietz, G.; Lange, J.; Rimbach, G. Molecular and dietary regulation of beta, beta-carotene 15,15’-monooxygenase 1 (BCMO1). Arch. Biochem. Biophys. 2010, 502, 8–16. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.B.; Nie, Z.J.; Cao, Z.M.; Hua, D.; Gu, R.B.; Xu, P. Carotenoid levels in soft tissues of triangle pearl mussel Hyriopsis cumingii with different color nacreous layer. J. Dalian Ocean. Univ. 2012, 27, 265–268. [Google Scholar]
- Wouters, R.; Lavens, P.; Nieto, J.; Sorgeloos, P. Penaeid shrimp broodstock nutrition: An updated review on research and development. Aquaculture 2001, 202, 1–21. [Google Scholar] [CrossRef]
- Wade, N.M.; Gabaudan, J.; Glencross, B.D. A review of carotenoid utilisation and function in crustacean aquaculture. Rev. Aquac. 2017, 9, 141–156. [Google Scholar] [CrossRef]
- Huang, S.; Wang, J.; Yue, W.C.; Chen, J.; Gaughan, S.; Lu, W.Q.; Lu, G.Q.; Wang, C.H. Transcriptomic variation of hepatopancreas reveals the energy metabolism and biological processes associated with molting in Chinese mitten crab, Eriocheir sinensis. Sci. Rep. 2015, 5, 14015. [Google Scholar] [CrossRef] [PubMed]
- Panganiban, G.; Sebring, A.; Nagy, L.; Carroll, S. The development of crustacean limbs and the evolution of arthropods. Science 1995, 270, 1363–1366. [Google Scholar] [CrossRef] [PubMed]
- Fang, F.; Yuan, Y.; Jin, M.; Shi, B.; Zhu, T.T.; Luo, J.X.; Lu, J.J.; Wang, X.X.; Jiao, L.F.; Zhou, Q.C. Hepatopancreas transcriptome analysis reveals the molecular responses to different dietary n-3 PUFA lipid sources in the swimming crab Portunus trituberculatus. Aquaculture 2021, 543, 737016. [Google Scholar] [CrossRef]
- Wang, W.; Wu, X.G.; Liu, Z.J.; Zheng, H.J.; Cheng, Y.X. Insights into hepatopancreatic functions for nutrition metabolism and ovarian development in the crab Portunus trituberculatus: Gene discovery in the comparative transcriptome of different hepatopancreas stages. PLoS ONE 2014, 9, e84921. [Google Scholar] [CrossRef]
- McGrane, M.M. Vitamin A regulation of gene expression: Molecular mechanism of a prototype gene. J. Nutr. Biochem. 2007, 18, 497–508. [Google Scholar] [CrossRef]
- Yun, J.; Finkel, T. Mitohormesis. Cell Metab. 2014, 19, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.Y.; Liu, M.F.; Yan, C.C.; Yang, H.; Wu, Z.X.; Liu, Y.J.; Su, N.K.; Hou, J.L.; Zhang, J.H.; Yang, F.S.; et al. CRISPR/Cas9-mediated deletion of β, β-carotene 9′, 10′-oxygenase gene (EcBCO2) from Exopalaemon carinicauda. Int. J. Biol. Macromol. 2020, 151, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Ribaya-Mercado, J.D. Influence of dietary fat on beta-carotene absorption and bioconversion into vitamin A. Nutr. Rev. 2002, 60, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Boonyaratpalin, M.; Thongrod, S.; Supamattaya, K.; Britton, G.; Schlipalius, L.E. Effects of β-carotene source, Dunaliella salina, and astaxanthin on pigmentation, growth, survival and health of Penaeus monodon. Aquac. Res. 2001, 32, 182–190. [Google Scholar] [CrossRef]
- Supamattaya, K.; Kiriratnikom, S.; Boonyaratpalin, M.; Borowitzka, L. Effect of a Dunaliella extract on growth performance, health condition, immune response and disease resistance in black tiger shrimp (Penaeus monodon). Aquaculture 2005, 248, 207–216. [Google Scholar] [CrossRef]
- Green, A.S.; Fascetti, A.J. Meeting the vitamin A requirement: The efficacy and importance of β-carotene in animal species. Sci. World J. 2016, 2016, 7393620. [Google Scholar] [CrossRef]
- Wan, S.; Li, Q.; Yu, H.; Liu, S.K.; Kong, L.F. Transcriptome analysis based on dietary beta-carotene supplement reveals genes potentially involved in carotenoid metabolism in Crassostrea gigas. Gene 2022, 818, 146226. [Google Scholar] [CrossRef] [PubMed]
- Parker, R.S. Carotenoids .4. Absorption, metabolism, and transport of carotenoids. FASEB J. 1996, 10, 542–551. [Google Scholar] [CrossRef] [PubMed]
- Vanvliet, T.; Schreurs, W.H.P.; Vandenberg, H. Intestinal beta-carotene absorption and cleavage in men-response of beta-carotene and retinyl esters in the triglyceride-rich lipoprotein fraction after a single oral dose of beta-carotene. Am. J. Clin. Nutr. 1995, 62, 110–116. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Peng, J.; Xiang, W.Z.; Tang, Q.M.; Sun, N.; Chen, F.; Yuan, J.P. Comparative analysis of astaxanthin and its esters in the mutant E1 of Haematococcus pluvialis and other green algae by HPLC with a C30 column. Sci. China Ser. C-Life Sci. 2008, 51, 1108–1115. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).