

Targeting NF-κB Signaling: Selected Small Molecules Downregulate Pro-Inflammatory Cytokines in Both Food Allergen and LPS-Induced Inflammation
 , ,
, ,
Abstract

1. Introduction
2. Results
2.1. Both LPS and Allergen Act d 1 Trigger Inflammation in THP-1 Differentiated Macrophages under Optimized Treatment Conditions
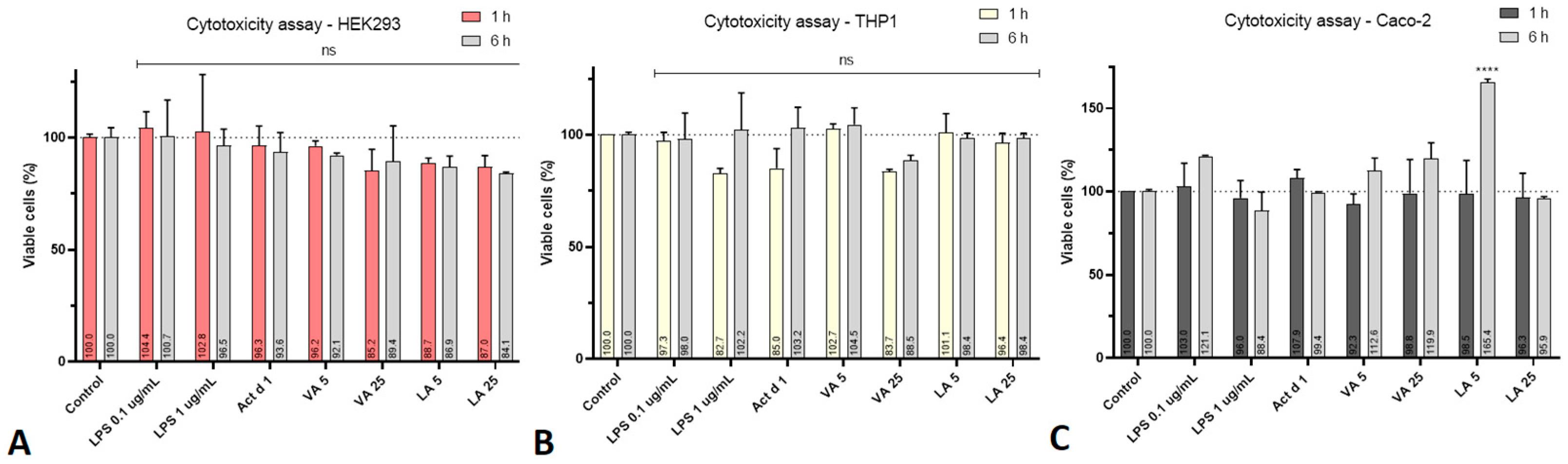
2.2. VA and LA Have No Impact on Cell Viability or Pro-Inflammatory Effect
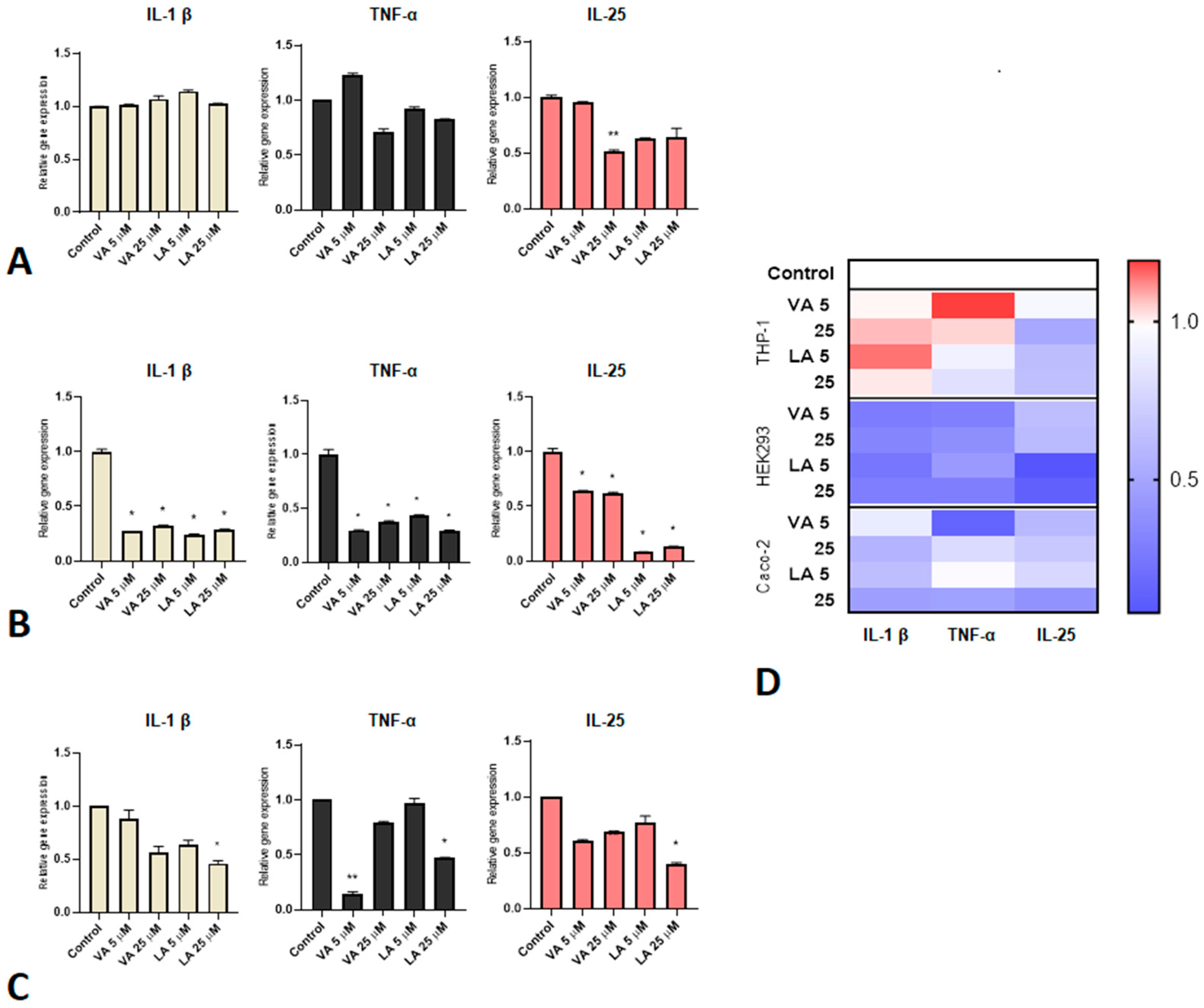
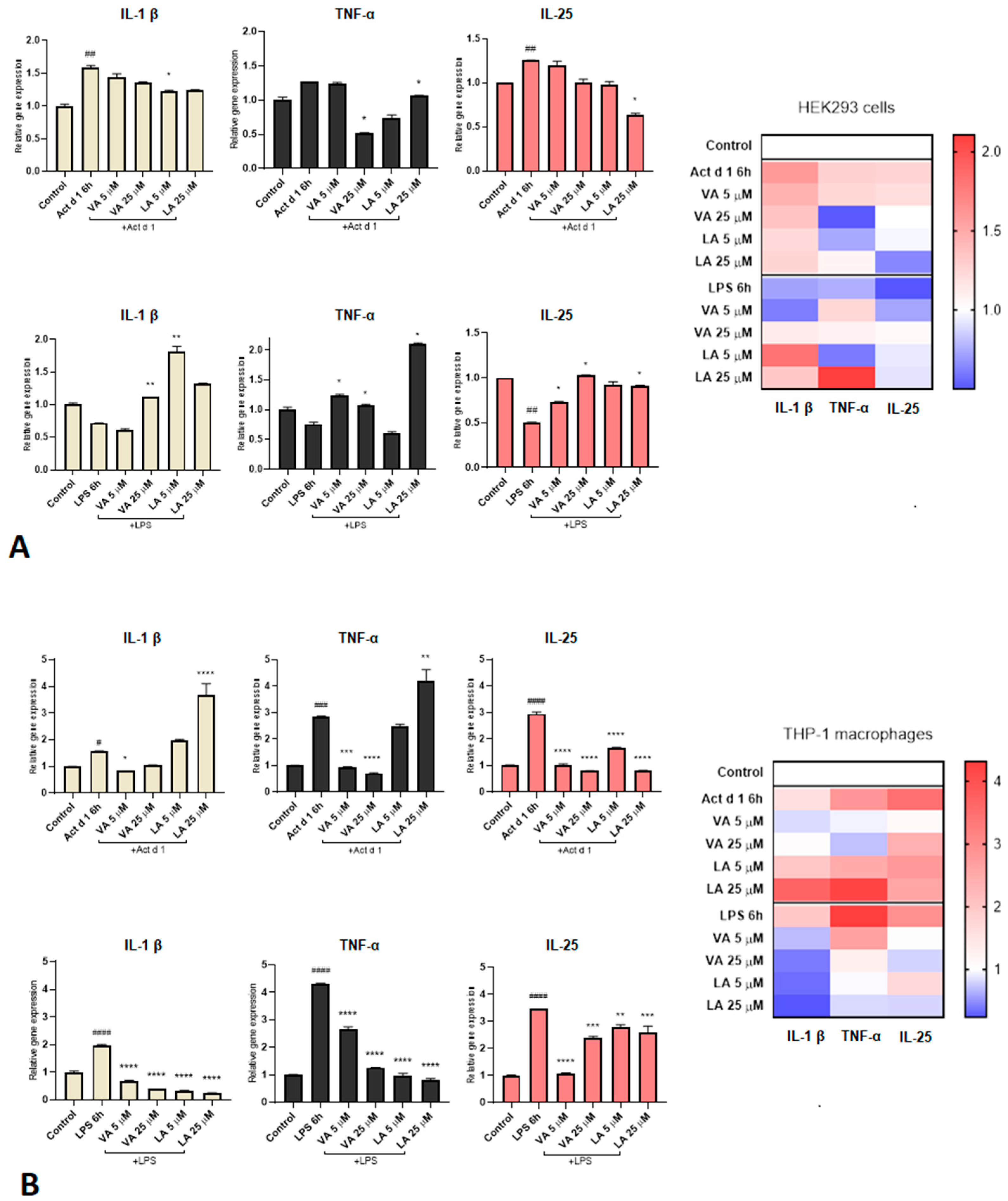
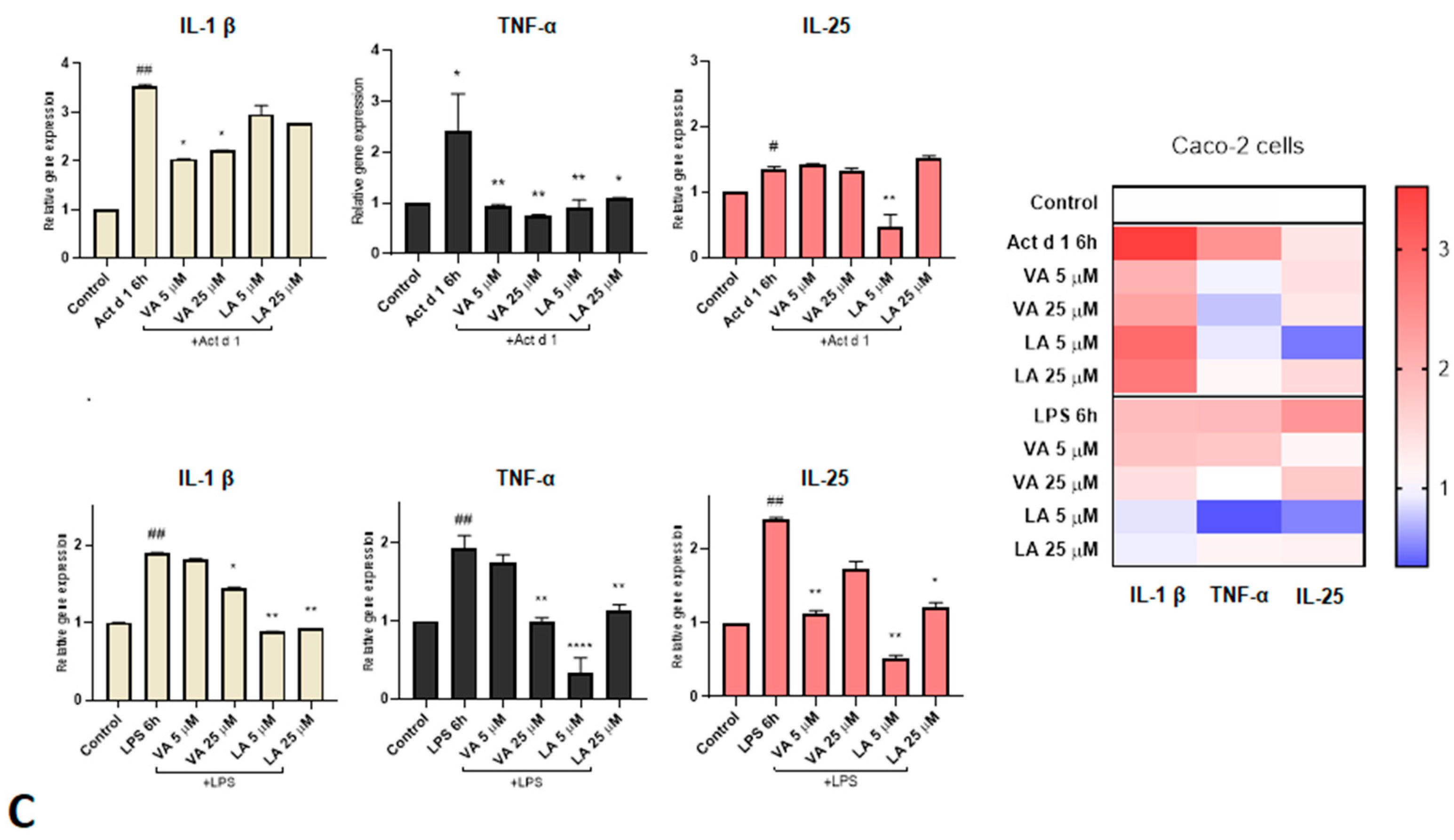
2.3. Small Molecules Reduce the Expression of Pro-Inflammatory Cytokines in Inflammation Responses
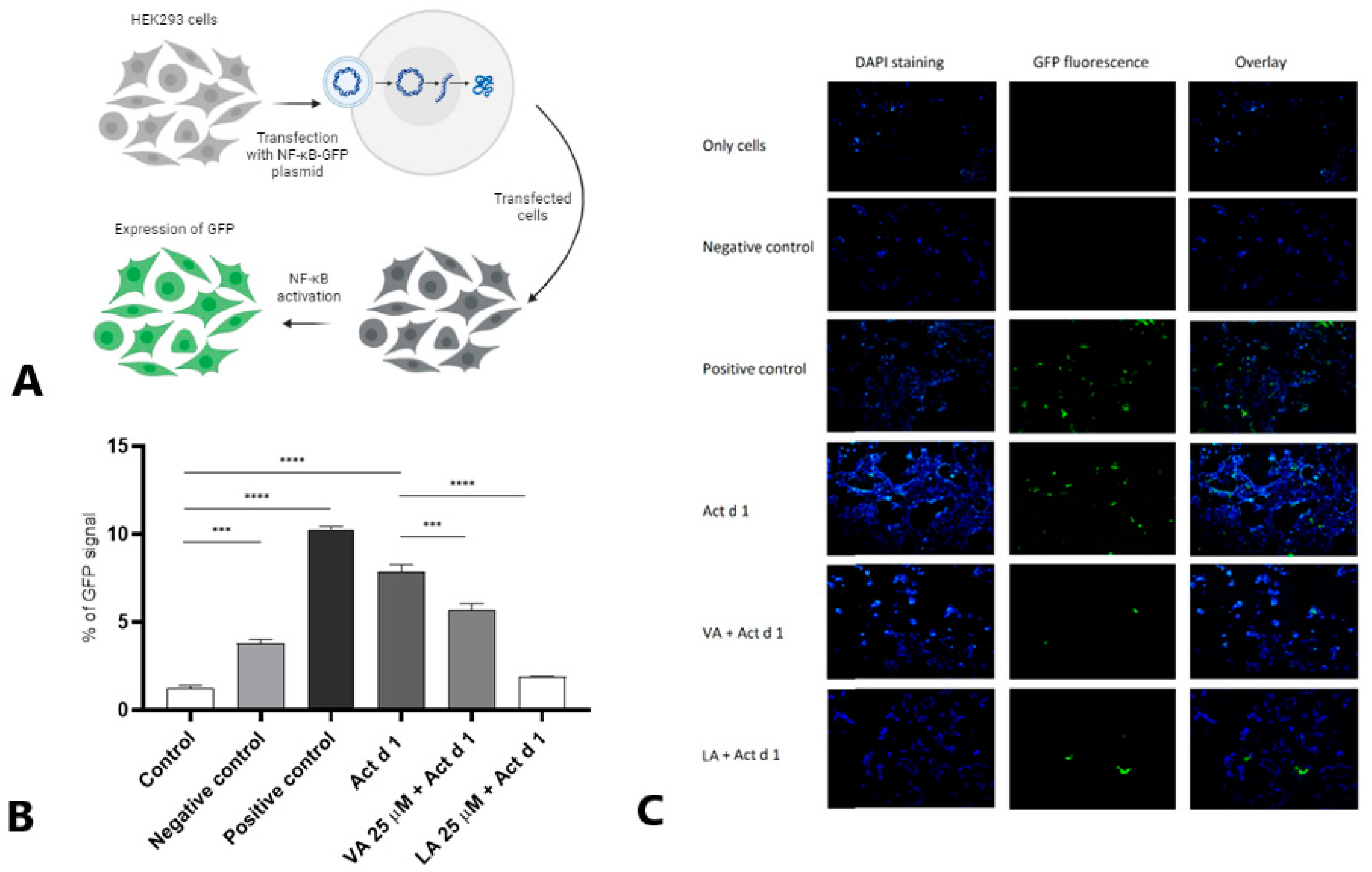
2.4. Small Molecules Attenuate NF-ĸB Activation in HEK293 Cells
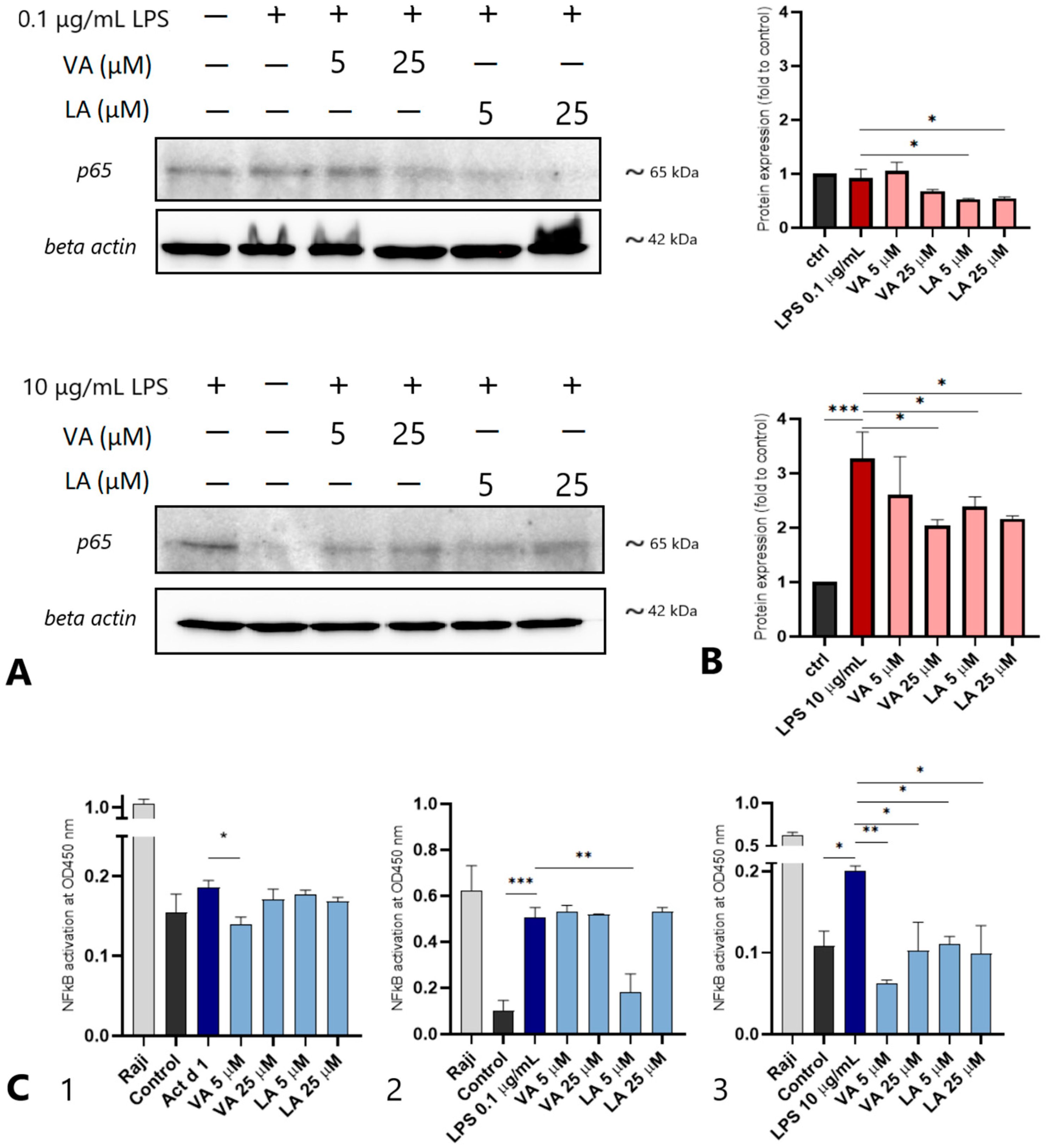
2.5. Small Molecules Reduce the NF-ĸB p65 Levels in THP-1-Derived Macrophages
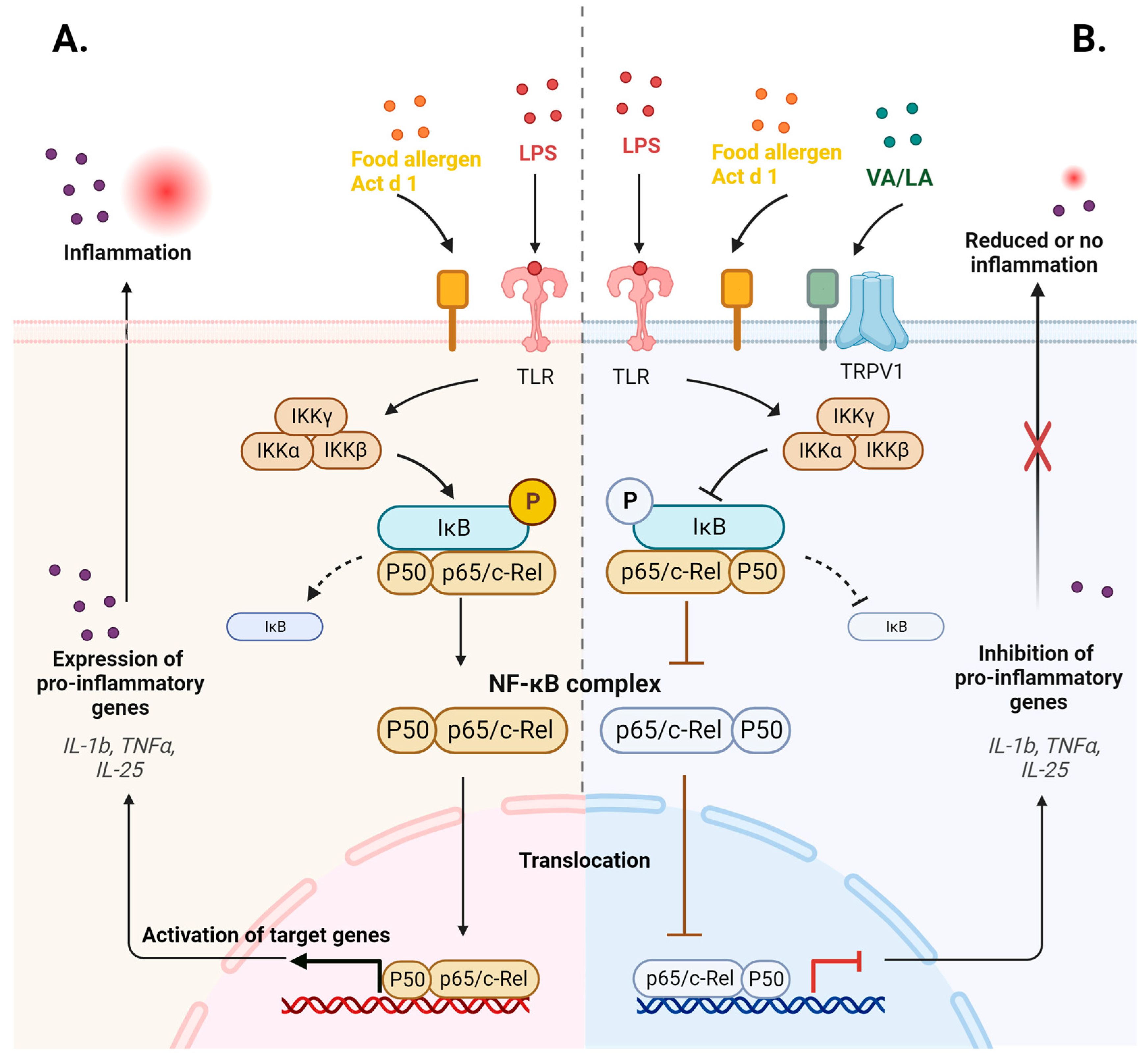
3. Discussion
4. Materials and Methods
4.1. Act d 1 Purification and Small Molecules’ Preparation
4.2. THP-1 Monocytes and Their Differentiation into Macrophages
4.3. HEK293 and Caco-2 Cell Culture
4.4. Cell Treatment
4.5. Flow Cytometric Analysis of THP-1-Derived Macrophages
4.6. Isolation of RNA and Gene Expression Analysis
4.7. CCK-8 Cytotoxicity Assay
4.8. NF-κB-GFP Reporter Assay
4.9. Western Blot
4.10. NF-κB DNA Binding Assay
4.11. Quantification and Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Afonina, I.S.; Zhong, Z.; Karin, M.; Beyaert, R. Limiting Inflammation—The Negative Regulation of NF-ΚB and the NLRP3 Inflammasome. Nat. Immunol. 2017, 18, 861–869. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Hayden, M.S. New Regulators of NF-ΚB in Inflammation. Nat. Rev. Immunol. 2008, 8, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S.C. NF-ΚB Signaling in Inflammation. Signal Transduct. Target. Ther. 2017, 2, 17023. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Zhang, L.; Zheng, B.; Zhang, X.; Zhang, H.; Zhao, A.; Yu, J.; Yang, Z.; Wen, J. CircACTA2 Inhibits NLRP3 Inflammasome-Mediated Inflammation via Interacting with NF-ΚB in Vascular Smooth Muscle Cells. Cell. Mol. Life Sci. 2023, 80, 229. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Gu, X.; Zhang, M.; Jiang, Q.; Xu, T. Di (2-Ethylhexyl) Phthalate and Polystyrene Microplastics Co-Exposure Caused Oxidative Stress to Activate NF-ΚB/NLRP3 Pathway Aggravated Pyroptosis and Inflammation in Mouse Kidney. Sci Total Environ. 2024, 926, 171817. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.C. Non-Canonical NF-ΚB Signaling Pathway. Cell Res. 2010, 21, 71–85. [Google Scholar] [CrossRef]
- Schuliga, M. NF-KappaB Signaling in Chronic Inflammatory Airway Disease. Biomolecules 2015, 5, 1266–1283. [Google Scholar] [CrossRef] [PubMed]
- Trivedi, S.; Labuz, D.; Deering-Rice, C.E.; Kim, C.U.; Christensen, H.; Aamodt, S.; Huecksteadt, T.; Sanders, K.; Warren, K.J. IL-33 Induces NF-ΚB Activation in ILC2 That Can Be Suppressed by in Vivo and Ex Vivo 17β-Estradiol. Front. Allergy 2022, 3, 143. [Google Scholar] [CrossRef] [PubMed]
- Ballegaard, A.-S.R.; Bøgh, K.L. Intestinal Protein Uptake and IgE-Mediated Food Allergy. Food Res. Int. 2023, 163, 112150. [Google Scholar] [CrossRef]
- Nešić, A.; Čavić, M.; Popović, M.; Zlatanova, M.; Pieters, R.; Smit, J.; Gavrović-Jankulović, M. The Kiwifruit Allergen Act d 1 Activates NF-ΚB Signaling and Affects MRNA Expression of TJ Proteins and Innate pro-Allergenic Cytokines. Biomolecules 2019, 9, 816. [Google Scholar] [CrossRef]
- Grozdanović, M.M.; Drakulić, B.J.; Gavrović-Jankulović, M. Conformational Mobility of Active and E-64-Inhibited Actinidin. Biochim. Biophys. Acta Gen. Subj. 2013, 1830, 4790–4799. [Google Scholar] [CrossRef]
- Grozdanovic, M.M.; Ostojic, S.; Aleksic, I.; Andjelkovic, U.; Petersen, A.; Gavrovic-Jankulovic, M. Active Actinidin Retains Function upon Gastro-Intestinal Digestion and Is More Thermostable than the E-64-Inhibited Counterpart. J. Sci. Food Agric. 2014, 94, 3046–3052. [Google Scholar] [CrossRef] [PubMed]
- Cavic, M.; Grozdanovic, M.M.; Bajic, A.; Jankovic, R.; Andjus, P.R.; Gavrovic-Jankulovic, M. The Effect of Kiwifruit (Actinidia Deliciosa) Cysteine Protease Actinidin on the Occludin Tight Junction Network in T84 Intestinal Epithelial Cells. Food Chem. Toxicol. 2014, 72, 61–68. [Google Scholar] [CrossRef]
- Peterson, L.W.; Artis, D. Intestinal Epithelial Cells: Regulators of Barrier Function and Immune Homeostasis. Nat. Rev. Immunol. 2014, 14, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Chanput, W.; Mes, J.J.; Wichers, H.J. THP-1 Cell Line: An in Vitro Cell Model for Immune Modulation Approach. Int. Immunopharmacol. 2014, 23, 37–45. [Google Scholar] [CrossRef]
- Teodorowicz, M.; Perdijk, O.; Verhoek, I.; Govers, C.; Savelkoul, H.F.J.; Tang, Y.; Wichers, H.; Broersen, K. Optimized Triton X-114 Assisted Lipopolysaccharide (LPS) Removal Method Reveals the Immunomodulatory Effect of Food Proteins. PLoS ONE 2017, 12, e0173778. [Google Scholar] [CrossRef]
- Tandon, A.; Harioudh, M.K.; Ishrat, N.; Tripathi, A.K.; Srivastava, S.; Ghosh, J.K. An MD2-Derived Peptide Promotes LPS Aggregation, Facilitates Its Internalization in THP-1 Cells, and Inhibits LPS-Induced pro-Inflammatory Responses. Cell. Mol. Life Sci. 2018, 75, 2431–2446. [Google Scholar] [CrossRef] [PubMed]
- Deme, P.; Narasimhulu, C.A.; Parthasarathy, S. Identification and Evaluation of Anti-Inflammatory Properties of Aqueous Components Extracted from Sesame (Sesamum indicum) Oil. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2018, 1087–1088, 61–69. [Google Scholar] [CrossRef]
- Rosa, A.; Atzeri, A.; Deiana, M.; Melis, M.P.; Incani, A.; Corona, G.; Loru, D.; Appendino, G.; Dessi, M.A. Protective Effect of Vanilloids against Tert-Butyl Hydroperoxide-Induced Oxidative Stress in Vero Cells Culture. J. Agric. Food Chem. 2008, 56, 3546–3553. [Google Scholar] [CrossRef]
- Jung, H.J.; Song, Y.S.; Lim, C.J.; Park, E.H. Anti-Angiogenic, Anti-Inflammatory and Anti-Nociceptive Activities of Vanillyl Alcohol. Arch. Pharm. Res. 2008, 31, 1275–1279. [Google Scholar] [CrossRef]
- Zhao, D.; Jiang, Y.; Sun, J.; Li, H.; Huang, M.; Sun, X.; Zhao, M. Elucidation of The Anti-Inflammatory Effect of Vanillin In Lps-Activated THP-1 Cells. J. Food Sci. 2019, 84, 1920–1928. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.W.; Koppula, S.; Kim, J.W.; Lim, H.W.; Hwang, J.W.; Kim, I.S.; Park, P.J.; Choi, D.K. Modulation of LPS-Stimulated Neuroinflammation in BV-2 Microglia by Gastrodia Elata: 4-Hydroxybenzyl Alcohol Is the Bioactive Candidate. J. Ethnopharmacol. 2012, 139, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.C.; Kim, S.J.; Kim, D.S.; Jeon, Y.D.; Park, S.J.; Lee, H.S.; Um, J.Y.; Hong, S.H. Vanillic Acid Inhibits Inflammatory Mediators by Suppressing NF-ΚB in Lipopolysaccharide-Stimulated Mouse Peritoneal Macrophages. Immunopharmacol. Immunotoxicol. 2011, 33, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Dayrit, F.M. The Properties of Lauric Acid and Their Significance in Coconut Oil. J. Am. Oil Chem. Soc. 2015, 92, 1–15. [Google Scholar] [CrossRef]
- Trbojević Ivić, J.; Milosavić, N.; Dimitrijević, A.; Gavrović Jankulović, M.; Bezbradica, D.; Kolarski, D.; Veličković, D. Synthesis of Medium-Chain Length Capsinoids from Coconut Oil Catalyzed by Candida Rugosa Lipases. Food Chem. 2017, 218, 505–508. [Google Scholar] [CrossRef] [PubMed]
- Weatherill, A.R.; Lee, J.Y.; Zhao, L.; Lemay, D.G.; Youn, H.S.; Hwang, D.H. Saturated and Polyunsaturated Fatty Acids Reciprocally Modulate Dendritic Cell Functions Mediated through TLR4. J. Immunol. 2005, 174, 5390–5397. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Shimizu, K.; Kondo, R. Anti-Androgenic Activity of Fatty Acids. Chem. Biodivers. 2009, 6, 503–512. [Google Scholar] [CrossRef] [PubMed]
- Khan, H.U.; Aamir, K.; Jusuf, P.R.; Sethi, G.; Sisinthy, S.P.; Ghildyal, R.; Arya, A. Lauric Acid Ameliorates Lipopolysaccharide (LPS)-Induced Liver Inflammation by Mediating TLR4/MyD88 Pathway in Sprague Dawley (SD) Rats. Life Sci. 2021, 265, 118750. [Google Scholar] [CrossRef] [PubMed]
- Nešić, A.; Stam, A.; Čavić, M.; Ten Klooster, J.P.; Pieters, R.; Smit, J.; Gavrović-Jankulović, M. Activation of Epithelial Cells by the Major Kiwifruit Allergen Act d 1 in Human and Mouse-Derived Intestinal Model. J. Funct. Foods 2019, 62, 103556. [Google Scholar] [CrossRef]
- Huang, H.; Fletcher, A.; Niu, Y.; Wang, T.T.Y.; Yu, L. Characterization of Lipopolysaccharide-Stimulated Cytokine Expression in Macrophages and Monocytes. Inflamm. Res. 2012, 61, 1329–1338. [Google Scholar] [CrossRef]
- Glaser, K.B.; Falduto, M.; Metzger, R.; Pederson, T.; Pease, L.; Shiosaki, K.; Morgan, D.W. Expression, Release, and Regulation of Human TNFa from Stable Transfectants of HEK-293 Cells. Inflamm. Res. 1997, 46 (Suppl. S2), S127–S128. [Google Scholar] [CrossRef] [PubMed]
- Burgueño, J.F.; Abreu, M.T. Epithelial Toll-like Receptors and Their Role in Gut Homeostasis and Disease. Nat. Rev. Gastroenterol. Hepatol. 2020, 17, 263–278. [Google Scholar] [CrossRef] [PubMed]
- Saha, D.; Koli, S.; Patgaonkar, M.; Reddy, K.V.R. Expression of Hemoglobin-α and β Subunits in Human Vaginal Epithelial Cells and Their Functional Significance. PLoS ONE 2017, 12, e0171084. [Google Scholar] [CrossRef] [PubMed]
- Chanput, W.; Mes, J.J.; Savelkoul, H.F.J.; Wichers, H.J. Characterization of Polarized THP-1 Macrophages and Polarizing Ability of LPS and Food Compounds. Food Funct. 2013, 4, 266–276. [Google Scholar] [CrossRef]
- Genin, M.; Clement, F.; Fattaccioli, A.; Raes, M.; Michiels, C. M1 and M2 Macrophages Derived from THP-1 Cells Differentially Modulate the Response of Cancer Cells to Etoposide. BMC Cancer 2015, 15, 577. [Google Scholar] [CrossRef]
- Ezzat, M.I.; Hassan, M.; Abdelhalim, M.A.; El-Desoky, A.M.; Mohamed, S.O.; Ezzat, S.M. Immunomodulatory Effect of Noni Fruit and Its Isolates: Insights into Cell-Mediated Immune Response and Inhibition of LPS-Induced THP-1 Macrophage Inflammation. Food Funct. 2021, 12, 3170–3179. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, J.; Peng, X.; Lv, B.; Wang, P.; Zhao, X.; Yu, B. Geraniin Inhibits LPS-Induced THP-1 Macrophages Switching to M1 Phenotype via SOCS1/NF-ΚB Pathway. Inflammation 2016, 39, 1421–1433. [Google Scholar] [CrossRef] [PubMed]
- Moser, B.A.; Escalante-Buendia, Y.; Steinhardt, R.C.; Rosenberger, M.G.; Cassaidy, B.J.; Naorem, N.; Chon, A.C.; Nguyen, M.H.; Tran, N.T.; Esser-Kahn, A.P. Small Molecule NF-ΚB Inhibitors as Immune Potentiators for Enhancement of Vaccine Adjuvants. Front. Immunol. 2020, 11, 511513. [Google Scholar] [CrossRef]
- Bezerra-Filho, C.S.M.; Barboza, J.N.; Souza, M.T.S.; Sabry, P.; Ismail, N.S.M.; de Sousa, D.P. Therapeutic Potential of Vanillin and Its Main Metabolites to Regulate the Inflammatory Response and Oxidative Stress. Mini-Rev. Med. Chem. 2019, 19, 1681–1693. [Google Scholar] [CrossRef]
- Huang, W.C.; Tsai, T.H.; Chuang, L.T.; Li, Y.Y.; Zouboulis, C.C.; Tsai, P.J. Anti-Bacterial and Anti-Inflammatory Properties of Capric Acid against Propionibacterium Acnes: A Comparative Study with Lauric Acid. J. Dermatol. Sci. 2014, 73, 232–240. [Google Scholar] [CrossRef]
- Bujak, J.K.; Kosmala, D.; Szopa, I.M.; Majchrzak, K.; Bednarczyk, P. Inflammation, Cancer and Immunity—Implication of TRPV1 Channel. Front. Oncol. 2019, 9, 1087. [Google Scholar] [CrossRef] [PubMed]
- Andrei, C.; Zanfirescu, A.; Nițulescu, G.M.; Olaru, O.T.; Negreș, S. Natural Active Ingredients and TRPV1 Modulation: Focus on Key Chemical Moieties Involved in Ligand–Target Interaction. Plants 2023, 12, 339. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wang, P.; Wang, J.; Xu, J.; Liu, C.; Qiao, H.; Gan, L.; Duan, E.; Zhang, Y.; Wang, M.; et al. Zinc Laurate Protects against Intestinal Barrier Dysfunction and Inflammation Induced by ETEC in a Mice Model. Nutrients 2023, 15, 54. [Google Scholar] [CrossRef] [PubMed]
- Chruszcz, M.; Chew, F.T.; Hoffmann-Sommergruber, K.; Hurlburt, B.K.; Mueller, G.A.; Pomés, A.; Rouvinen, J.; Villalba, M.; Wöhrl, B.M.; Breiteneder, H. Allergens and Their Associated Small Molecule Ligands—Their Dual Role in Sensitization. Allergy Eur. J. Allergy Clin. Immunol. 2021, 76, 2367–2382. [Google Scholar] [CrossRef] [PubMed]
- Park, M.H.; Hong, J.T. Roles of NF-ΚB in Cancer and Inflammatory Diseases and Their Therapeutic Approaches. Cells 2016, 5, 15. [Google Scholar] [CrossRef] [PubMed]
- Grozdanovic, M.M.; Čavić, M.; Nešić, A.; Andjelković, U.; Akbari, P.; Smit, J.J.; Gavrović-Jankulović, M. Kiwifruit Cysteine Protease Actinidin Compromises the Intestinal Barrier by Disrupting Tight Junctions. Biochim. Biophys. Acta Gen. Subj. 2016, 1860, 516–526. [Google Scholar] [CrossRef] [PubMed]
- Drapeau, G.R. [38] Protease from Staphyloccus Aureus. Methods Enzym. 1976, 45, 469–475. [Google Scholar] [CrossRef] [PubMed]
- Palacio, J.R.; Markert, U.R.; Martínez, P. Anti-Inflammatory Properties of N-Acetylcysteine on Lipopolysaccharide- Activated Macrophages. Inflamm. Res. 2011, 60, 695–704. [Google Scholar] [CrossRef]
- Chanput, W.; Mes, J.; Vreeburg, R.A.M.; Savelkoul, H.F.J.; Wichers, H.J. Transcription Profiles of LPS-Stimulated THP-1 Monocytes and Macrophages: A Tool to Study Inflammation Modulating Effects of Food-Derived Compounds. Food Funct. 2010, 1, 254–261. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Reference | Human Primer Sequence (5′-3′) | AT (°C) |
|---|---|---|---|
| GAPDH | NM_009386.2 | For: CGAGGCATCATCCCAAATAAGAAC Rev: TCCAGAAGTCTGCCCGATCAC | 65 |
| IL-1β | NM_000576.2 | For: AACCTCTTCGAGGCACAAGG Rev: GGCGAGCTCAGGTACTTCTG | 49 |
| IL-25 | NM_022789.3 | For: CCAGGTGGTTGCATTCTTGG Rev: TGGCTGTAGGTGTGGGTTCC | 49 |
| TNF-α | NM_000594.3 | For: GACAAGCCTGTAGCCCATGT Rev: CTCTGATGGCACCACCAACT | 49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zlatanova, M.; Nešić, A.; Trbojević-Ivić, J.; Četić, D.; Gavrović-Jankulović, M. Targeting NF-κB Signaling: Selected Small Molecules Downregulate Pro-Inflammatory Cytokines in Both Food Allergen and LPS-Induced Inflammation. Int. J. Mol. Sci. 2024, 25, 5798. https://doi.org/10.3390/ijms25115798
Zlatanova M, Nešić A, Trbojević-Ivić J, Četić D, Gavrović-Jankulović M. Targeting NF-κB Signaling: Selected Small Molecules Downregulate Pro-Inflammatory Cytokines in Both Food Allergen and LPS-Induced Inflammation. International Journal of Molecular Sciences. 2024; 25(11):5798. https://doi.org/10.3390/ijms25115798
Chicago/Turabian StyleZlatanova, Milena, Andrijana Nešić, Jovana Trbojević-Ivić, Danilo Četić, and Marija Gavrović-Jankulović. 2024. "Targeting NF-κB Signaling: Selected Small Molecules Downregulate Pro-Inflammatory Cytokines in Both Food Allergen and LPS-Induced Inflammation" International Journal of Molecular Sciences 25, no. 11: 5798. https://doi.org/10.3390/ijms25115798
APA StyleZlatanova, M., Nešić, A., Trbojević-Ivić, J., Četić, D., & Gavrović-Jankulović, M. (2024). Targeting NF-κB Signaling: Selected Small Molecules Downregulate Pro-Inflammatory Cytokines in Both Food Allergen and LPS-Induced Inflammation. International Journal of Molecular Sciences, 25(11), 5798. https://doi.org/10.3390/ijms25115798