
Natural Variation in the Promoter of GmSPL9d Affects Branch Number in Soybean


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
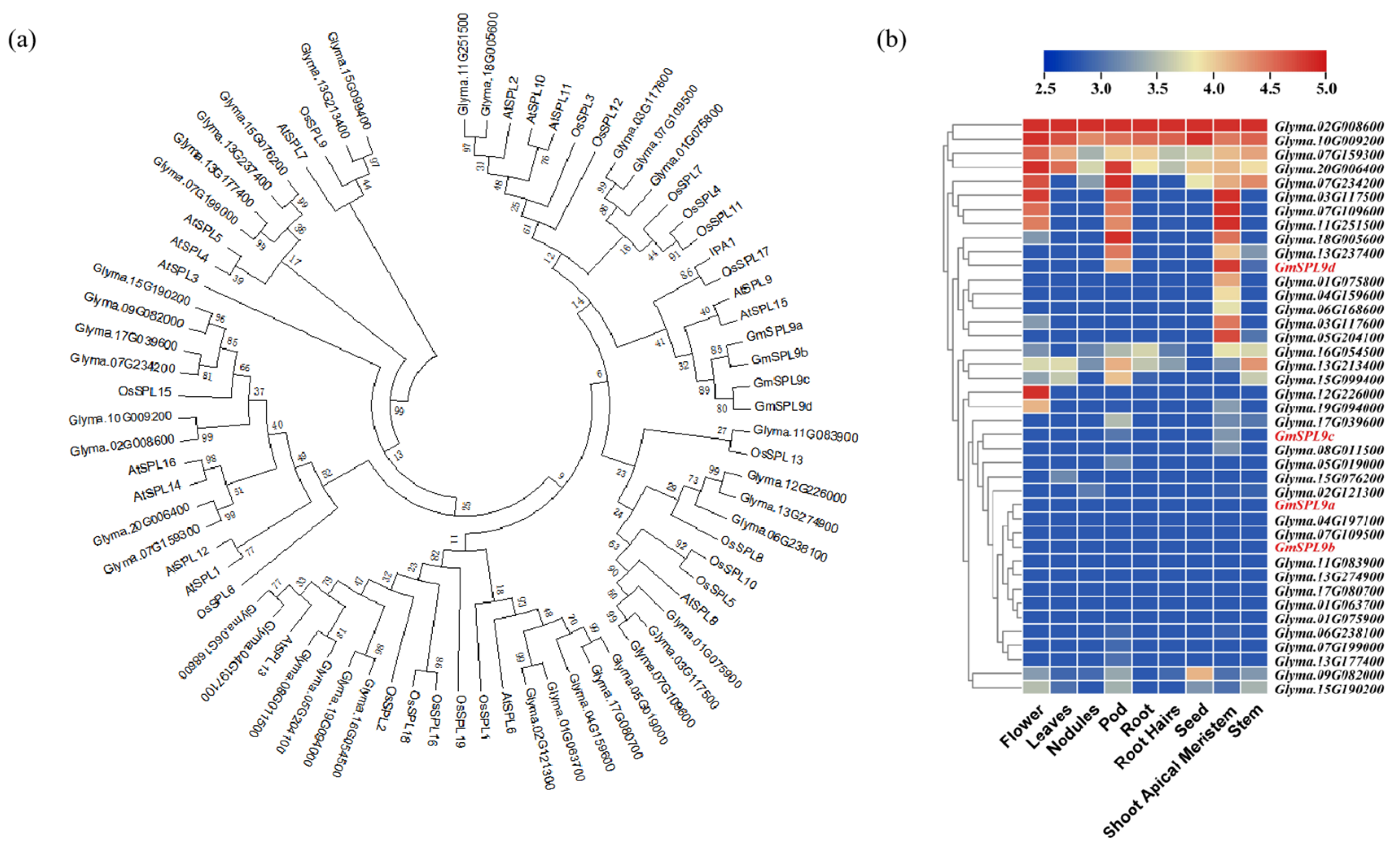
2.1. Identification of Soybean SPL Gene Family Members
2.2. GmSPL9d Expression Pattern and Subcellular Localization
2.3. Haplotype Analysis of GmSPL9d
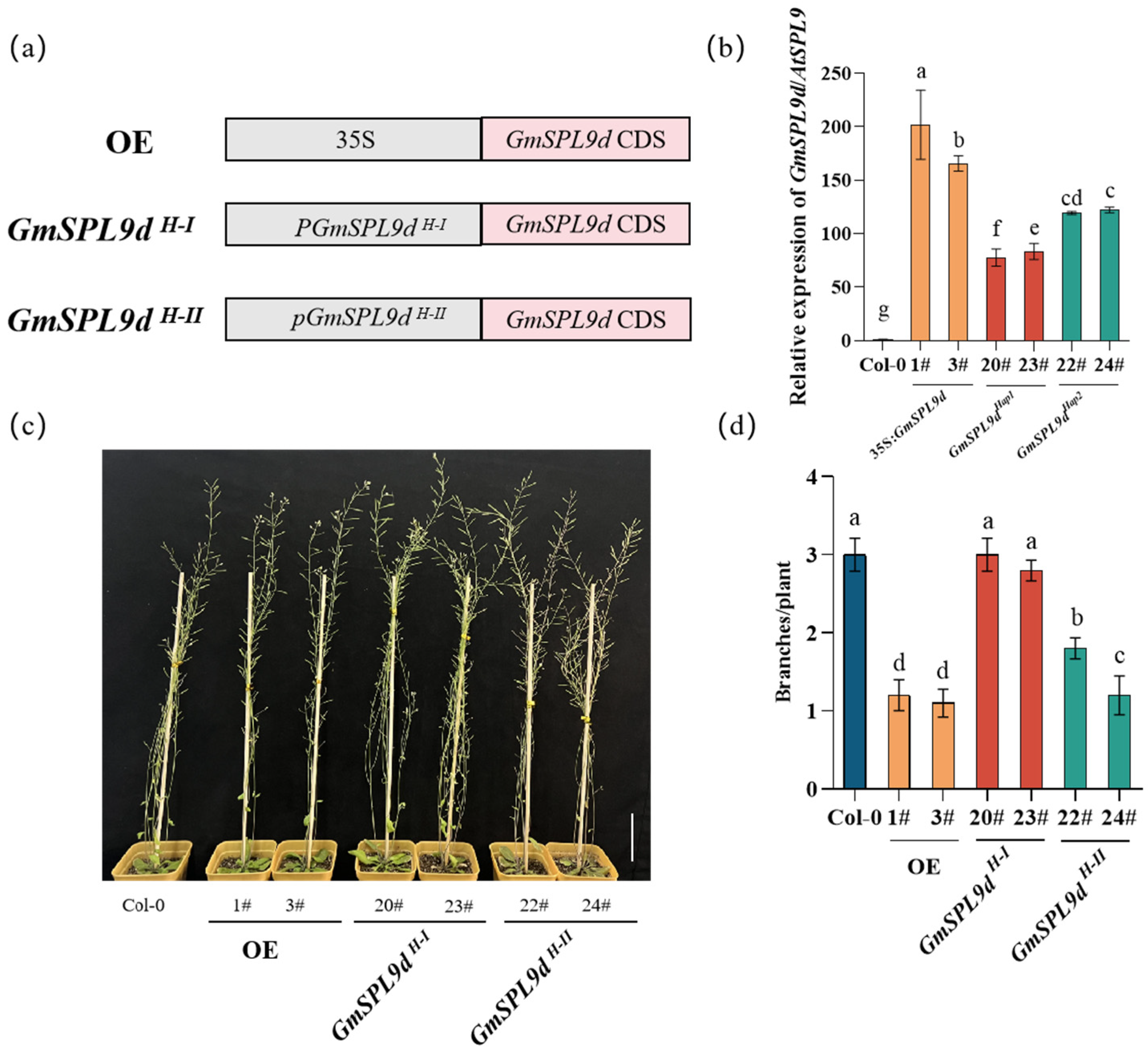
2.4. The Activity of the GmSPL9d Promoter Affected the Branch Number in Soybean
2.5. GmSPL9d H-II Haplotype Inhibited Arabidopsis Branching
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. RNA Extraction and qRT-PCR
4.3. Identification and Bioinformatics Analysis of GmSPL9d
4.4. Subcellular Localization
4.5. Histochemical Analysis of GUS Activity
4.6. Transient Transcription Activity Assay
4.7. Arabidopsis Genetic Transformation
4.8. Genetic Diversity Analysis
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, Y.H.; Zhou, G.Y.; Ma, J.X.; Jiang, W.K.; Jin, L.G.; Zhang, Z.H.; Guo, Y.; Zhang, J.B.; Sui, Y.; Zheng, L.T.; et al. De novo assembly of soybean wild relatives for pan-genome analysis of diversity and agronomic traits. Nat. Biotechnol. 2014, 32, 1045. [Google Scholar] [CrossRef] [PubMed]
- Miao, L.; Yang, S.N.; Zhang, K.; He, J.B.; Wu, C.H.; Ren, Y.H.; Gai, J.Y.; Li, Y. Natural variation and selection in GmSWEET39 affect soybean seed oil content. New Phytol. 2020, 225, 1651–1666. [Google Scholar] [CrossRef]
- Agudamu; Yoshihira, T.; Shiraiwa, T. Branch development responses to planting density and yield stability in soybean cultivars. Plant. Prod. Sci. 2016, 19, 331–339. [Google Scholar] [CrossRef]
- Xu, C.L.; Li, R.D.; Song, W.W.; Wu, T.T.; Sun, S.; Han, T.F.; Wu, C.X. High Density and Uniform Plant Distribution Improve Soybean Yield by Regulating Population Uniformity and Canopy Light Interception. Agronomy 2021, 11, 18. [Google Scholar] [CrossRef]
- Li, S.C.; Sun, Z.H.; Sang, Q.; Qin, C.; Kong, L.P.; Huang, X.; Liu, H.; Su, T.; Li, H.Y.; He, M.L.; et al. Soybean reduced internode 1 determines internode length and improves grain yield at dense planting. Nat. Commun. 2023, 14, 13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Q.; Gao, L.; Ke, M.Y.; Gao, Z.; Tu, T.L.; Huang, L.M.; Chen, J.M.; Guan, Y.F.; Huang, X.; Chen, X. GmPIN1-mediated auxin asymmetry regulates leaf petiole angle and plant architecture in soybean. J. Integr. Plant Biol. 2022, 64, 1325–1338. [Google Scholar] [CrossRef]
- Shim, S.; Ha, J.; Kim, M.Y.; Choi, M.S.; Kang, S.T.; Jeong, S.C.; Moon, J.K.; Lee, S.H. GmBRC1 is a Candidate Gene for Branching in Soybean (Glycine max (L.) Merrill). Int. J. Mol. Sci. 2019, 20, 15. [Google Scholar] [CrossRef] [PubMed]
- Liang, Q.J.; Chen, L.Y.; Yang, X.; Yang, H.; Liu, S.L.; Kou, K.; Fan, L.; Zhang, Z.F.; Duan, Z.B.; Yuan, Y.Q.; et al. Natural variation of Dt2 determines branching in soybean. Nat. Commun. 2022, 13, 9. [Google Scholar] [CrossRef]
- Wang, J.W.; Czech, B.; Weigel, D. miR156-Regulated SPL Transcription Factors Define an Endogenous Flowering Pathway in Arabidopsis thaliana. Cell 2009, 138, 738–749. [Google Scholar] [CrossRef]
- Jiao, Y.Q.; Wang, Y.H.; Xue, D.W.; Wang, J.; Yan, M.X.; Liu, G.F.; Dong, G.J.; Zeng, D.L.; Lu, Z.F.; Zhu, X.D.; et al. Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice. Nature Genet. 2010, 42, 541–544. [Google Scholar] [CrossRef]
- Miura, K.; Ikeda, M.; Matsubara, A.; Song, X.J.; Ito, M.; Asano, K.; Matsuoka, M.; Kitano, H.; Ashikari, M. OsSPL14 promotes panicle branching and higher grain productivity in rice. Nature Genet. 2010, 42, 545–549. [Google Scholar] [CrossRef]
- Yamasaki, K.; Kigawa, T.; Inoue, M.; Tateno, M.; Yamasaki, T.; Yabuki, T.; Aoki, M.; Seki, E.; Matsuda, T.; Nunokawa, E.; et al. A novel zinc-binding motif revealed by solution structures of DNA-binding domains of Arabidopsis SBP-family transcription factors. J. Mol. Biol. 2004, 337, 49–63. [Google Scholar] [CrossRef]
- Manning, K.; Tör, M.; Poole, M.; Hong, Y.; Thompson, A.J.; King, G.J.; Giovannoni, J.J.; Seymour, G.B. A naturally occurring epigenetic mutation in a gene encoding an SBP-box transcription factor inhibits tomato fruit ripening. Nature Genet. 2006, 38, 948–952. [Google Scholar] [CrossRef]
- Xu, M.L.; Hu, T.Q.; Zhao, J.F.; Park, M.Y.; Earley, K.W.; Wu, G.; Yang, L.; Poethig, R.S. Developmental Functions of miR156-Regulated SQUAMOSA PROMOTER BINDING PROTEIN-LIKE (SPL) Genes in Arabidopsis thaliana. PLoS Genet. 2016, 12, 29. [Google Scholar] [CrossRef]
- Shikata, M.; Koyama, T.; Mitsuda, N.; Ohme-Takagi, M. Arabidopsis SBP-Box Genes SPL10, SPL11 and SPL2 Control Morphological Change in Association with Shoot Maturation in the Reproductive Phase. Plant Cell Physiol. 2009, 50, 2133–2145. [Google Scholar] [CrossRef]
- Zhang, Q.Q.; Wang, J.G.; Wang, L.Y.; Wang, J.F.; Wang, Q.; Yu, P.; Bai, M.Y.; Fan, M. Gibberellin repression of axillary bud formation in Arabidopsis by modulation of DELLA-SPL9 complex activity. J. Integr. Plant Biol. 2020, 62, 421–432. [Google Scholar] [CrossRef] [PubMed]
- Xie, Y.R.; Liu, Y.; Ma, M.D.; Zhou, Q.; Zhao, Y.P.; Zhao, B.B.; Wang, B.B.; Wei, H.B.; Wang, H.Y. Arabidopsis FHY3 and FAR1 integrate light and strigolactone signaling to regulate branching. Nat. Commun. 2020, 11, 13. [Google Scholar] [CrossRef]
- Sun, Z.X.; Su, C.; Yun, J.X.; Jiang, Q.; Wang, L.X.; Wang, Y.N.; Cao, D.; Zhao, F.; Zhao, Q.S.; Zhang, M.C.; et al. Genetic improvement of the shoot architecture and yield in soya bean plants via the manipulation of GmmiR156b. Plant Biotechnol. J. 2019, 17, 50–62. [Google Scholar] [CrossRef]
- Bao, A.L.; Chen, H.F.; Chen, L.M.; Chen, S.L.; Hao, Q.N.; Guo, W.; Qiu, D.Z.; Shan, Z.H.; Yang, Z.L.; Yuan, S.L.; et al. CRISPR/Cas9-mediated targeted mutagenesis of GmSPL9 genes alters plant architecture in soybean. Bmc Plant Biol. 2019, 19, 12. [Google Scholar] [CrossRef] [PubMed]
- Arazi, T.; Talmor-Neiman, M.; Stav, R.; Riese, M.; Huijser, P.; Baulcombe, D.C. Cloning and characterization of micro-RNAs from moss. Plant J. 2005, 43, 837–848. [Google Scholar] [CrossRef] [PubMed]
- Eriksson, M.; Moseley, J.L.; Tottey, S.; del Campo, J.A.; Quinn, J.; Kim, Y.; Merchant, S. Genetic dissection of nutritional copper signaling in chlamydomonas distinguishes regulatory and target genes. Genetics 2004, 168, 795–807. [Google Scholar] [CrossRef] [PubMed]
- Kropat, J.; Tottey, S.; Birkenbihl, R.P.; Depège, N.; Huijser, P.; Merchant, S. A regulator of nutritional copper signaling in Chlamydomonas is an SBP domain protein that recognizes the GTAC core of copper response element. Proc. Natl. Acad. Sci. USA 2005, 102, 18730–18735. [Google Scholar] [CrossRef] [PubMed]
- Lännenpää, M.; Jänönen, I.; Hölttä-Vuori, M.; Gardemeister, M.; Porali, I.; Sopanen, T. A new SBP-box gene BpSPL1 in silver birch (Betula pendula). Physiol. Plant. 2004, 120, 491–500. [Google Scholar] [CrossRef] [PubMed]
- Moreno, M.A.; Harper, L.C.; Krueger, R.W.; Dellaporta, S.L.; Freeling, M. liguleless1 encodes a nuclear-localized protein required for induction of ligules and auricles during maize leaf organogenesis. Genes Dev. 1997, 11, 616–628. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.B.; Zhang, Z.L.; Liu, D.M.; Zhang, K.; Li, A.L.; Mao, L. SQUAMOSA Promoter-Binding Protein-Like Transcription Factors: Star Players for Plant Growth and Development. J. Integr. Plant Biol. 2010, 52, 946–951. [Google Scholar] [CrossRef] [PubMed]
- Schmutz, J.; Cannon, S.B.; Schlueter, J.; Ma, J.X.; Mitros, T.; Nelson, W.; Hyten, D.L.; Song, Q.J.; Thelen, J.J.; Cheng, J.L.; et al. Genome sequence of the palaeopolyploid soybean (vol 463, pg 178, 2010). Nature 2010, 465, 120. [Google Scholar] [CrossRef]
- Haerizadeh, F.; Wong, C.E.; Singh, M.B.; Bhalla, P.L. Genome-wide analysis of gene expression in soybean shoot apical meristem. Plant Mol. Biol. 2009, 69, 711–727. [Google Scholar] [CrossRef] [PubMed]
- Begum, N.; Xiao, Y.T.; Wang, L.; Li, D.M.; Irshad, A.; Zhao, T.J. Arbuscular mycorrhizal fungus Rhizophagus irregularis alleviates drought stress in soybean with overexpressing the GmSPL9d gene by promoting photosynthetic apparatus and regulating the antioxidant system. Microbiol. Res. 2023, 273, 14. [Google Scholar] [CrossRef]
- Yun, J.X.; Sun, Z.X.; Jiang, Q.; Wang, Y.N.; Wang, C.; Luo, Y.Q.; Zhang, F.R.; Li, X. The miR156b-GmSPL9d module modulates nodulation by targeting multiple core nodulation genes in soybean. New Phytol. 2022, 233, 1881–1899. [Google Scholar] [CrossRef] [PubMed]
- Springer, N.; de León, N.; Grotewold, E. Challenges of Translating Gene Regulatory Information into Agronomic Improvements. Trends Plant Sci. 2019, 24, 1075–1082. [Google Scholar] [CrossRef]
- Swinnen, G.; Goossens, A.; Pauwels, L. Lessons from Domestication: Targeting Cis-Regulatory Elements for Crop Improvement. Trends in Plant Science 2016, 21, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Duan, P.G.; Xu, J.S.; Zeng, D.L.; Zhang, B.L.; Geng, M.F.; Zhang, G.Z.; Huang, K.; Huang, L.J.; Xu, R.; Ge, S.; et al. Natural Variation in the Promoter of GSE5 Contributes to Grain Size Diversity in Rice. Mol. Plant 2017, 10, 685–694. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.F.; Chen, J.; Zheng, X.M.; Wu, F.Q.; Lin, Q.B.; Heng, Y.Q.; Tian, P.; Cheng, Z.J.; Yu, X.W.; Zhou, K.N.; et al. GW5 acts in the brassinosteroid signalling pathway to regulate grain width and weight in rice. Nat. Plants 2017, 3, 7. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Ye, H.; Vuong, T.D.; Zhou, L.J.; Do, T.D.; Chhapekar, S.S.; Zhao, W.Q.; Li, B.; Jin, T.; Gu, J.B.; et al. A novel natural variation in the promoter of GmCHX1 regulates conditional gene expression to improve salt tolerance in soybean. J. Exp. Bot. 2024, 75, 1051–1062. [Google Scholar] [CrossRef] [PubMed]
- Greb, T.; Clarenz, O.; Schäfer, E.; Müller, D.; Herrero, R.; Schmitz, G.; Theres, K. Molecular analysis of the LATERAL SUPPRESSOR gene in Arabidopsis reveals a conserved control mechanism for axillary meristem formation. Genes Dev. 2003, 17, 1175–1187. [Google Scholar] [CrossRef] [PubMed]
- Shi, B.H.; Zhang, C.; Tian, C.H.; Wang, J.; Wang, Q.; Xu, T.F.; Xu, Y.; Ohno, C.; Sablowski, R.; Heisler, M.G.; et al. Two-Step Regulation of a Meristematic Cell Population Acting in Shoot Branching in Arabidopsis. PLoS Genet. 2016, 12, 20. [Google Scholar] [CrossRef] [PubMed]
- Tucker, M.R.; Laux, T. Connecting the paths in plant stem cell regulation. Trends Cell Biol. 2007, 17, 403–410. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.Q.; Yuan, C.Q.; Cong, T.C.; Zhang, Q.X. The Secrets of Meristems Initiation: Axillary Meristem Initiation and Floral Meristem Initiation. Plants-Basel 2023, 12, 18. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.; Sun, Y.Y.; Shan, Z.; He, J.B.; Wang, N.; Gai, J.Y.; Li, Y. Natural variation in the promoter of GsERD15B affects salt tolerance in soybean. Plant Biotechnol. J. 2021, 19, 1155–1169. [Google Scholar] [CrossRef]
- Ahmad, M.Z.; Rehman, N.U.; Yu, S.W.; Zhou, Y.Z.; ul Haq, B.; Wang, J.J.; Li, P.H.; Zeng, Z.X.; Zhao, J. GmMAX2-D14 and -KAI interaction-mediated SL and KAR signaling play essential roles in soybean root nodulation. Plant J. 2020, 101, 334–351. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Yang, H.F.; Wang, X.C.; Qiu, Y.J.; Tian, L.H.; Qi, X.Q.; Qu, L. Cytochrome P450 family member CYP96B5 hydroxylates alkanes to primary alcohols and is involved in rice leaf cuticular wax synthesis. New Phytol. 2020, 225, 2094–2107. [Google Scholar] [CrossRef] [PubMed]
- Mistry, J.; Finn, R.D.; Eddy, S.R.; Bateman, A.; Punta, M. Challenges in homology search: HMMER3 and convergent evolution of coiled-coil regions. Nucleic Acids Res. 2013, 41, 10. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef]
- Clough; Steven, J.; Bent; Andrew, F. Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 1998, 16, 735–743. [Google Scholar] [CrossRef] [PubMed]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, D.; Zheng, H.; Li, J.; Wan, M.; Shu, K.; Wang, W.; Hu, X.; Hu, Y.; Qiu, L.; Wang, X. Natural Variation in the Promoter of GmSPL9d Affects Branch Number in Soybean. Int. J. Mol. Sci. 2024, 25, 5991. https://doi.org/10.3390/ijms25115991
Zhao D, Zheng H, Li J, Wan M, Shu K, Wang W, Hu X, Hu Y, Qiu L, Wang X. Natural Variation in the Promoter of GmSPL9d Affects Branch Number in Soybean. International Journal of Molecular Sciences. 2024; 25(11):5991. https://doi.org/10.3390/ijms25115991
Chicago/Turabian StyleZhao, Duo, Haowei Zheng, Jiajia Li, Mingyue Wan, Kuo Shu, Wenhui Wang, Xiaoyu Hu, Yu Hu, Lijuan Qiu, and Xiaobo Wang. 2024. "Natural Variation in the Promoter of GmSPL9d Affects Branch Number in Soybean" International Journal of Molecular Sciences 25, no. 11: 5991. https://doi.org/10.3390/ijms25115991
APA StyleZhao, D., Zheng, H., Li, J., Wan, M., Shu, K., Wang, W., Hu, X., Hu, Y., Qiu, L., & Wang, X. (2024). Natural Variation in the Promoter of GmSPL9d Affects Branch Number in Soybean. International Journal of Molecular Sciences, 25(11), 5991. https://doi.org/10.3390/ijms25115991