


Secukinumab and Dead Sea Climatotherapy Impact Resolved Psoriasis Skin Differently Potentially Affecting Disease Memory
 , ,
, ,  , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. Dead Sea Climatotherapy and Secukinumab Treatment Reduced Epidermal Thickness, Proliferation, CD1a+, CD3+, CD4+, CD8+, CD11c+, CD45RO+, and MPO+ Cells
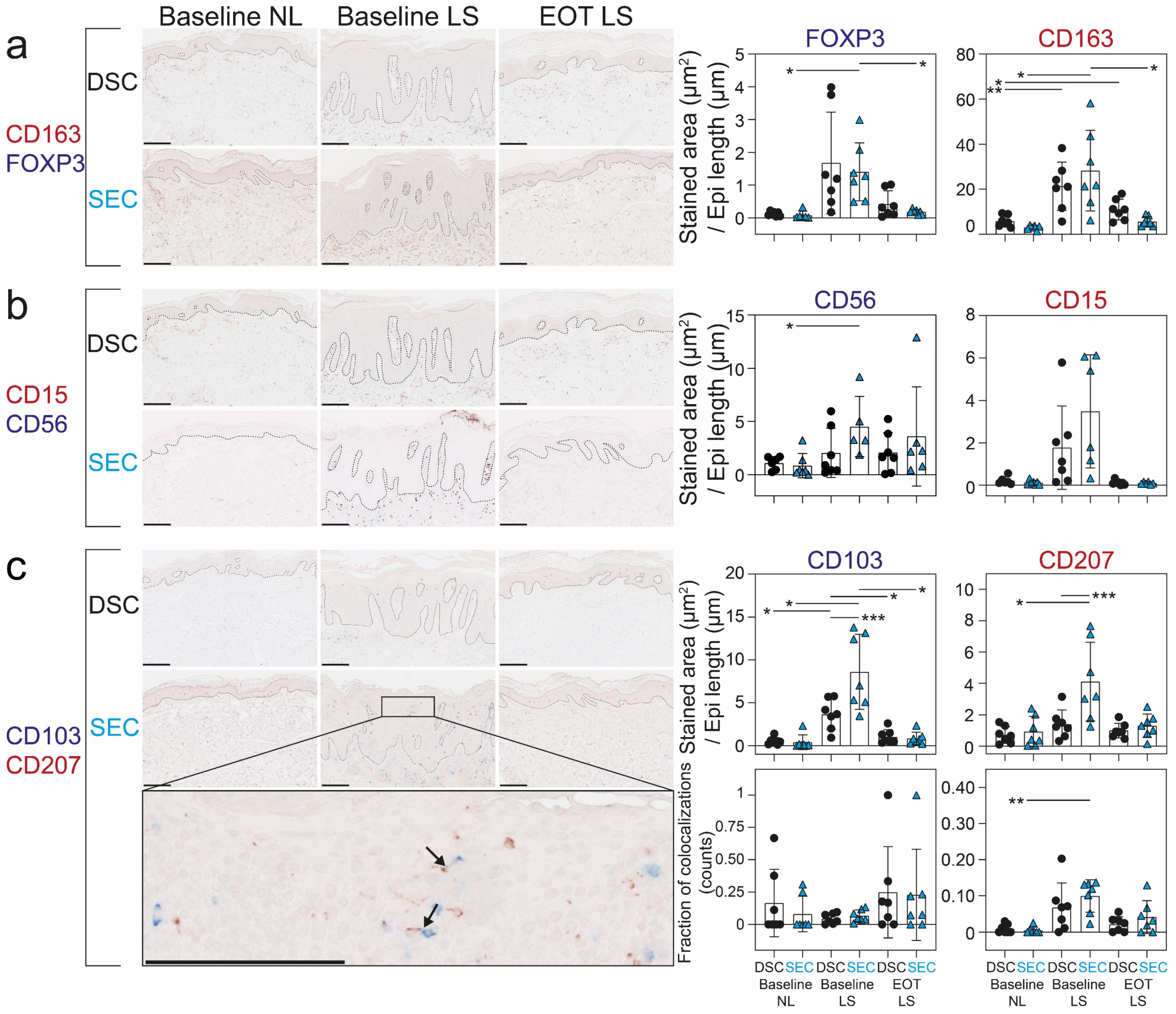
2.2. Dead Sea Climatotherapy and Secukinumab Treatment Differentially Reduced CD15+, CD56+, CD103+, CD163+, CD207+, and FOXP3+ Cells
2.3. CD8+CD49a+ T-Cell Counts Correlated with Disease Severity But the Proportion of CD8+CD49a+ T-Cells Out of CD8+ and CD49+ T-Cells Did Not Change during Treatment
2.4. Dead Sea Climatotherapy and Secukinumab Treatment Altered the Transcriptome in Patients Responding to Treatment
2.5. A Molecular Scar Was Still Present in Completely Resolved Psoriasis Skin after Dead Sea Climatotherapy and Secukinumab Treatment
2.6. Dead Sea Climatotherapy and Secukinumab Treatment Differentially Altered the Transcriptome in Clinically Resolved Psoriasis Skin
2.7. Digital Spatial Profiling Successfully Assessed the Proteome of CD45+ Cells, MelanA+ Cells, and the Microenvironment in the Skin
2.8. Digital Spatial Profiling of the Proteome in Selected Areas of the Skin Showed Distinct Differences in Resolved Skin from Dead Sea Climatotherapy and Secukinumab Treatment
3. Discussion
4. Materials and Methods
4.1. Study Populations
4.2. Statistical Analyses and Methods
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- de Oliveira, M.d.F.S.P.; de Oliveira Rocha, B.; Duarte, G.V. Psoriasis: Classical and emerging comorbidities. An. Bras. Dermatol. 2015, 90, 9–20. [Google Scholar] [CrossRef]
- Griffiths, C.E.M.; Armstrong, A.W.; Gudjonsson, J.E.; Barker, J.N.W.N. Psoriasis. Lancet 2021, 397, 1301–1315. [Google Scholar] [CrossRef] [PubMed]
- Parisi, R.; Iskandar, I.Y.K.K.; Kontopantelis, E.; Augustin, M.; Griffiths, C.E.M.M.; Ashcroft, D.M. National, regional, and worldwide epidemiology of psoriasis: Systematic analysis and modelling study. BMJ 2020, 369, m1590. [Google Scholar] [CrossRef] [PubMed]
- Reid, C.; Griffiths, C. Psoriasis and Treatment: Past, Present and Future Aspects. Acta Derm. Venereol. 2020, 100, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Mrowietz, U.; Steinz, K.; Gerdes, S. Psoriasis: To treat or to manage? Exp. Dermatol. 2014, 23, 705–709. [Google Scholar] [CrossRef] [PubMed]
- Bertelsen, T.; Ljungberg, C.; Litman, T.; Huppertz, C.; Hennze, R.; Rønholt, K.; Iversen, L.; Johansen, C. IκBζ is a key player in the antipsoriatic effects of secukinumab. J. Allergy Clin. Immunol. 2020, 145, 379–390. [Google Scholar] [CrossRef] [PubMed]
- Emmanuel, T.; Lybæk, D.; Johansen, C.; Iversen, L. Effect of Dead Sea Climatotherapy on Psoriasis; A Prospective Cohort Study. Front. Med. 2020, 7, 83. [Google Scholar] [CrossRef] [PubMed]
- Hodak, E.; Gottlieb, A.B.; Segal, T.; Politi, Y.; Maron, L.; Sulkes, J.; David, M. Climatotherapy at the Dead Sea is a remittive therapy for psoriasis: Combined effects on epidermal and immunologic activation. J. Am. Acad. Dermatol. 2003, 49, 451–457. [Google Scholar] [CrossRef] [PubMed]
- Kudish, A.I.; Evseev, E.G.; Cohen, G.; Harari, M. Ultraviolet Measurements and Photoclimatotherapy for Psoriasis at the Dead Sea: 25 Years of Experience. Int. J. Environ. Res. Public Health 2022, 19, 12364. [Google Scholar] [CrossRef]
- Even-Paz, Z.; Gumon, R.; Kipnis, V.; Abels, D.; Efron, D. Dead Sea sun versus Dead Sea water in the treatment of psoriasis. J. Dermatol. Treat. 1996, 7, 83–86. [Google Scholar] [CrossRef]
- Svendsen, M.T.; Jeyabalan, J.; Andersen, K.E.; Andersen, F.; Johannessen, H. Worldwide utilization of topical remedies in treatment of psoriasis: A systematic review. J. Dermatol. Treat. 2017, 28, 374–383. [Google Scholar] [CrossRef] [PubMed]
- Jensen, K.K.; Serup, J.; Alsing, K.K. Psoriasis and seasonal variation: A systematic review on reports from Northern and Central Europe—Little overall variation but distinctive subsets with improvement in summer or wintertime. Ski. Res. Technol. 2022, 28, 180–186. [Google Scholar] [CrossRef] [PubMed]
- Breuckmann, F.; Von Kobyletzki, G.; Avermaete, A.; Radenhausen, M.; Höxtermann, S.; Pieck, C.; Schöneborn, P.; Gambichler, T.; Freitag, M.; Hoffmann, K.; et al. Mechanisms of apoptosis: UVA1-induced immediate and UVB-induced delayed apoptosis in human T cells in vitro. J. Eur. Acad. Dermatol. Venereol. 2003, 17, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Addison, R.; Weatherhead, S.C.; Pawitri, A.; Smith, G.R.; Rider, A.; Grantham, H.J.; Cockell, S.J.; Reynolds, N.J. Therapeutic wavelengths of ultraviolet B radiation activate apoptotic, circadian rhythm, redox signalling and key canonical pathways in psoriatic epidermis. Redox Biol. 2021, 41, 101924. [Google Scholar] [CrossRef] [PubMed]
- Frieder, J.; Kivelevitch, D.; Menter, A. Secukinumab: A review of the anti-IL-17A biologic for the treatment of psoriasis. Ther. Adv. Chronic. Dis. 2018, 9, 5–21. [Google Scholar] [CrossRef]
- Emmanuel, T.; Lybaek, D.; Johansen, C.; Iversen, L. Non-random Plaque-site Recurrence of Psoriasis in Patients Treated with Dead Sea Climatotherapy. Acta Derm. Venereol. 2019, 99, 909–910. [Google Scholar] [CrossRef] [PubMed]
- Cheuk, S.; Wikén, M.; Blomqvist, L.; Nylén, S.; Talme, T.; Ståhle, M.; Eidsmo, L. Epidermal Th22 and Tc17 Cells Form a Localized Disease Memory in Clinically Healed Psoriasis. J. Immunol. 2014, 192, 3111–3120. [Google Scholar] [CrossRef] [PubMed]
- Puig, L.; Costanzo, A.; Muñoz-Elías, E.J.; Jazra, M.; Wegner, S.; Paul, C.F.; Conrad, C. The biological basis of disease recurrence in psoriasis: A historical perspective and current models. Br. J. Dermatol. 2022, 186, 773–781. [Google Scholar] [CrossRef] [PubMed]
- Emmanuel, T.; Mistegård, J.; Bregnhøj, A.; Johansen, C.; Iversen, L. Tissue-Resident Memory T Cells in Skin Diseases: A Systematic Review. Int. J. Mol. Sci. 2021, 22, 9004. [Google Scholar] [CrossRef]
- Pham, J.P.; Wark, K.J.L.; Woods, J.; Frew, J.W. Resident cutaneous memory T-cells: A clinical review of their role in chronic inflammatory dermatoses and potential as therapeutic targets. Br. J. Dermatol. 2023, 189, 656–663. [Google Scholar] [CrossRef]
- Christo, S.N.; Evrard, M.; Park, S.L.; Gandolfo, L.C.; Burn, T.N.; Fonseca, R.; Newman, D.M.; Alexandre, Y.O.; Collins, N.; Zamudio, N.M.; et al. Discrete tissue microenvironments instruct diversity in resident memory T cell function and plasticity. Nat. Immunol. 2021, 22, 1140–1151. [Google Scholar] [CrossRef] [PubMed]
- Christo, S.N.; Evrard, M.; Park, S.L.; Gandolfo, L.C.; Burn, T.N.; Fonseca, R.; Newman, D.M.; Alexandre, Y.O.; Collins, N.; Zamudio, N.M.; et al. Survival of tissue-resident memory T cells requires exogenous lipid uptake and metabolism. Nature 2017, 543, 252–256. [Google Scholar] [CrossRef] [PubMed]
- Richer, M.J.; Pewe, L.L.; Hancox, L.S.; Hartwig, S.M.; Varga, S.M.; Harty, J.T. Inflammatory IL-15 is required for optimal memory T cell responses. J. Clin. Investig. 2015, 125, 3477–3490. [Google Scholar] [CrossRef]
- Adachi, T.; Kobayashi, T.; Sugihara, E.; Yamada, T.; Ikuta, K.; Pittaluga, S.; Saya, H.; Amagai, M.; Nagao, K. Hair follicle–derived IL-7 and IL-15 mediate skin-resident memory T cell homeostasis and lymphoma. Nat. Med. 2015, 21, 1272–1279. [Google Scholar] [CrossRef] [PubMed]
- Rønholt, K.; Iversen, L. Old and New Biological Therapies for Psoriasis. Int. J. Mol. Sci. 2017, 18, 2297. [Google Scholar] [CrossRef] [PubMed]
- Korman, N.J.; Malatestinic, W.; Goldblum, O.M.; Murage, M.J.; Renda, L.; Lin, C.-Y.; Lucas, J.; Middleton, C.; Lobosco, S. Assessment of the benefit of achieving complete versus almost complete skin clearance in psoriasis: A patient’s perspective. J. Dermatol. Treat. 2022, 33, 733–739. [Google Scholar] [CrossRef]
- Armstrong, A.W.; Patil, D.; Levi, E.; McGuiness, C.B.; Wang, X.; Wang, Y.; Chen, C.-C.; Nguyen, E.; Yamauchi, P.S. Real-World Satisfaction with Secukinumab in Clearing the Skin of Patients with Plaque Psoriasis through 24 Months of Follow-Up: Results from US Dermatology Electronic Medical Records. Dermatol. Ther. 2021, 11, 1733–1749. [Google Scholar] [CrossRef]
- Reich, K.; Hansen, J.B.; Puig, L.; Konstantinou, M.P.; Warren, R.B. Complete clearance and Psoriasis Area and Severity Index response for brodalumab and ustekinumab by previous treatment history in AMAGINE-2 and AMAGINE-3. J. Eur. Acad. Dermatol. Venereol. 2021, 35, 2034–2044. [Google Scholar] [CrossRef]
- Ben Abdallah, H.; Johansen, C.; Iversen, L. Key Signaling Pathways in Psoriasis: Recent Insights from Antipsoriatic Therapeutics. Psoriasis Targets Ther. 2021, 11, 83–97. [Google Scholar] [CrossRef]
- Iversen, L.; Eidsmo, L.; Austad, J.; de Rie, M.; Osmancevic, A.; Skov, L.; Talme, T.; Bachmann, I.; van de Kerkhof, P.; Stahle, M.; et al. Secukinumab treatment in new-onset psoriasis: Aiming to understand the potential for disease modification—Rationale and design of the randomized, multicenter STEPIn study. J. Eur. Acad. Dermatol. Venereol. 2018, 32, 1930–1939. [Google Scholar] [CrossRef]
- Iversen, L.; Conrad, C.; Eidsmo, L.; Costanzo, A.; Narbutt, J.; Pinter, A.; Kingo, K.; Diaz, R.R.; Kolbinger, F.; Nanna, M.; et al. Secukinumab demonstrates superiority over narrow-band ultraviolet B phototherapy in new-onset moderate to severe plaque psoriasis patients: Week 52 results from the STEPIn study. J. Eur. Acad. Dermatol. Venereol. 2023, 37, 1004–1016. [Google Scholar] [CrossRef] [PubMed]
- Eyerich, K.; Weisenseel, P.; Pinter, A.; Schäkel, K.; Asadullah, K.; Wegner, S.; Muñoz-Elias, E.J.; Bartz, H.; Taut, F.J.H.; Reich, K. IL-23 blockade with guselkumab potentially modifies psoriasis pathogenesis: Rationale and study protocol of a phase 3b, randomised, double-blind, multicentre study in participants with moderate-to-severe plaque-type psoriasis (GUIDE). BMJ Open 2021, 11, e049822. [Google Scholar] [CrossRef] [PubMed]
- Cheuk, S.; Schlums, H.; Sérézal, I.G.; Martini, E.; Chiang, S.C.; Marquardt, N.; Gibbs, A.; Detlofsson, E.; Introini, A.; Forkel, M.; et al. CD49a Expression Defines Tissue-Resident CD8+ T Cells Poised for Cytotoxic Function in Human Skin. Immunity 2017, 46, 287–300. [Google Scholar] [CrossRef]
- Conrad, C.; Boyman, O.; Tonel, G.; Tun-Kyi, A.; Laggner, U.; de Fougerolles, A.; Kotelianski, V.; Gardner, H.; O Nestle, F. α1β1 integrin is crucial for accumulation of epidermal T cells and the development of psoriasis. Nat. Med. 2007, 13, 836–842. [Google Scholar] [CrossRef]
- Suárez-Fariñas, M.; Fuentes-Duculan, J.; Lowes, M.A.; Krueger, J.G. Resolved psoriasis lesions retain expression of a subset of disease-related genes. J. Investig. Dermatol. 2011, 131, 391–400. [Google Scholar] [CrossRef]
- Schwingen, J.; Kaplan, M.; Kurschus, F.C. Review—Current Concepts in Inflammatory Skin Diseases Evolved by Transcriptome Analysis: In-Depth Analysis of Atopic Dermatitis and Psoriasis. Int. J. Mol. Sci. 2020, 21, 699. [Google Scholar] [CrossRef]
- Vacharanukrauh, P.; Meephansan, J.; Ponnikorn, S.; Tangtanatakul, P.; Soonthornchai, W.; Wongpiyabovorn, J.; Ingkaninanda, P.; Morita, A. Transcriptome profiling in psoriasis: NB-UVB treatment-associated transcriptional changes and modulation of autoinflammation in perilesional skin in early-phase disease. J. Dermatol. Sci. 2022, 107, 123–132. [Google Scholar] [CrossRef]
- Gudjonsson, J.E.; Ding, J.; Johnston, A.; Tejasvi, T.; Guzman, A.M.; Nair, R.P.; Voorhees, J.J.; Abecasis, G.R.; Elder, J.T. Assessment of the Psoriatic Transcriptome in a Large Sample: Additional Regulated Genes and Comparisons with In Vitro Models. J. Investig. Dermatol. 2010, 130, 1829–1840. [Google Scholar] [CrossRef] [PubMed]
- Su, W.; Zhao, Y.; Wei, Y.; Zhang, X.; Ji, J.; Yang, S. Exploring the Pathogenesis of Psoriasis Complicated with Atherosclerosis via Microarray Data Analysis. Front. Immunol. 2021, 12, 2045. [Google Scholar] [CrossRef]
- Rioux, G.; Ridha, Z.; Simard, M.; Turgeon, F.; Guérin, S.L.; Pouliot, R. Transcriptome Profiling Analyses in Psoriasis: A Dynamic Contribution of Keratinocytes to the Pathogenesis. Genes 2020, 11, 1155. [Google Scholar] [CrossRef]
- Li, J.; Yu, P. Genome-wide transcriptome analysis identifies alternative splicing regulatory network and key splicing factors in mouse and human psoriasis. Sci. Rep. 2018, 8, 4124. [Google Scholar] [CrossRef] [PubMed]
- Quaranta, M.; Knapp, B.; Garzorz, N.; Mattii, M.; Pullabhatla, V.; Pennino, D.; Andres, C.; Traidl-Hoffmann, C.; Cavani, A.; Theis, F.J.; et al. Intraindividual genome expression analysis reveals a specific molecular signature of psoriasis and eczema. Sci. Transl. Med. 2014, 6, 244ra90. [Google Scholar] [CrossRef] [PubMed]
- Wilsmann-Theis, D.; Wagenpfeil, J.; Holzinger, D.; Roth, J.; Koch, S.; Schnautz, S.; Bieber, T.; Wenzel, J. Among the S100 proteins, S100A12 is the most significant marker for psoriasis disease activity. J. Eur. Acad. Dermatol. Venereol. 2016, 30, 1165–1170. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Fariñas, M.; Li, K.; Fuentes-Duculan, J.; Hayden, K.; Brodmerkel, C.; Krueger, J.G. Expanding the Psoriasis Disease Profile: Interrogation of the Skin and Serum of Patients with Moderate-to-Severe Psoriasis. J. Investig. Dermatol. 2012, 132, 2552–2564. [Google Scholar] [CrossRef] [PubMed]
- Suárez-Fariñas, M.; Lowes, M.A.; Zaba, L.C.; Krueger, J.G. Evaluation of the Psoriasis Transcriptome across Different Studies by Gene Set Enrichment Analysis (GSEA). PLoS ONE 2010, 5, e10247. [Google Scholar] [CrossRef] [PubMed]
- Mitsui, H.; Suárez-Fariñas, M.; Belkin, D.A.; Levenkova, N.; Fuentes-Duculan, J.; Coats, I.; Fujita, H.; Krueger, J.G. Combined Use of Laser Capture Microdissection and cDNA Microarray Analysis Identifies Locally Expressed Disease-Related Genes in Focal Regions of Psoriasis Vulgaris Skin Lesions. J. Investig. Dermatol. 2012, 132, 1615–1626. [Google Scholar] [CrossRef]
- Arakawa, A.; Siewert, K.; Stöhr, J.; Besgen, P.; Kim, S.-M.; Rühl, G.; Nickel, J.; Vollmer, S.; Thomas, P.; Krebs, S.; et al. Melanocyte antigen triggers autoimmunity in human psoriasis. J. Exp. Med. 2015, 212, 2203–2212. [Google Scholar] [CrossRef] [PubMed]
- Merritt, C.R.; Ong, G.T.; Church, S.E.; Barker, K.; Danaher, P.; Geiss, G.; Hoang, M.; Jung, J.; Liang, Y.; McKay-Fleisch, J.; et al. Multiplex digital spatial profiling of proteins and RNA in fixed tissue. Nat. Biotechnol. 2020, 38, 586–599. [Google Scholar] [CrossRef] [PubMed]
- Esaki, H.; Ewald, D.A.; Ungar, B.; Rozenblit, M.; Zheng, X.; Xu, H.; Estrada, Y.D.; Peng, X.; Mitsui, H.; Litman, T.; et al. Identification of novel immune and barrier genes in atopic dermatitis by means of laser capture microdissection. J. Allergy Clin. Immunol. 2015, 135, 153–163. [Google Scholar] [CrossRef]
- Ignatov, B.; Sortebech, D.; Emmanuel, T.; Zhuravleva, E.; Eidsmo, L. Method for high-plex analysis of immune cells in human skin using the GeoMx system. Scand. J. Immunol. 2023, 99, e13326. [Google Scholar] [CrossRef]
- Benezeder, T.; Wolf, P. Resolution of plaque-type psoriasis: What is left behind (and reinitiates the disease). Semin. Immunopathol. 2019, 41, 633. [Google Scholar] [CrossRef] [PubMed]
- Pearse, A.D.; Gaskell, S.A.; Marks, R. Epidermal Changes in Human Skin Following Irradiation with Either UVB or UVA. J. Investig. Dermatol. 1987, 88, 83–87. [Google Scholar] [CrossRef] [PubMed]
- Emmanuel, T.; Petersen, A.; Houborg, H.I.; Rønsholdt, A.B.; Lybæk, D.; Steiniche, T.; Bregnhøj, A.; Iversen, L.; Johansen, C. Climatotherapy at the Dead Sea for psoriasis is a highly effective anti-inflammatory treatment in the short term: An immunohistochemical study. Exp. Dermatol. 2022, 31, 1136–1144. [Google Scholar] [CrossRef] [PubMed]
- de Winter, S.; Pavel, S.; Vink, A.A.; Roza, L. Solar-Simulated Skin Adaptation and its Effect on Subsequent UV-Induced Epidermal DNA Damage. J. Investig. Dermatol. 2001, 117, 678–682. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Hu, Y.; Yongqing, T.; Kim, J.; Hughes, V.A.; Le Nours, J.; Marquez, E.A.; Purcell, A.W.; Wan, Q.; Sugita, M.; et al. CD1a on Langerhans cells controls inflammatory skin disease. Nat. Immunol. 2016, 17, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Krueger, J.G.; Wharton, K.A., Jr.; Schlitt, T.; Suprun, M.; Torene, R.I.; Jiang, X.; Wang, C.Q.; Fuentes-Duculan, J.; Hartmann, N.; Peters, T.; et al. IL-17A inhibition by secukinumab induces early clinical, histopathologic, and molecular resolution of psoriasis. J. Allergy Clin. Immunol. 2019, 144, 750–763. [Google Scholar] [CrossRef] [PubMed]
- Zaba, L.C.; Cardinale, I.; Gilleaudeau, P.; Sullivan-Whalen, M.; Suárez-Fariñas, M.; Fuentes-Duculan, J.; Novitskaya, I.; Khatcherian, A.; Bluth, M.J.; Lowes, M.A.; et al. Amelioration of epidermal hyperplasia by TNF inhibition is associated with reduced Th17 responses. J. Exp. Med. 2007, 204, 3183–3194. [Google Scholar] [CrossRef] [PubMed]
- Tsoi, L.C.; Xing, X.; Xing, E.; Wasikowski, R.; Shao, S.; Zeng, C.; Plazyo, O.; Kirma, J.; Jiang, Y.; Billi, A.C.; et al. Non-invasive tape-stripping with high-resolution RNA profiling effectively captures a pre-inflammatory state in nonlesional psoriatic skin. J. Investig. Dermatol. 2021, 142, 1587–1596.e2. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Trepicchio, W.L.; Oestreicher, J.L.; Pittman, D.; Wang, F.; Chamian, F.; Dhodapkar, M.; Krueger, J.G. Increased Expression of Interleukin 23 p19 and p40 in Lesional Skin of Patients with Psoriasis Vulgaris. J. Exp. Med. 2004, 199, 125–130. [Google Scholar] [CrossRef]
- Zhou, X.; Krueger, J.G.; Kao, M.-C.J.; Lee, E.; Du, F.; Menter, A.; Wong, W.H.; Bowcock, A.M. Novel mechanisms of T-cell and dendritic cell activation revealed by profiling of psoriasis on the 63,100-element oligonucleotide array. Physiol. Genom. 2003, 13, 69–78. [Google Scholar] [CrossRef]
- Hochberg, M.; Zeligson, S.; Amariglio, N.; Rechavi, G.; Ingber, A.; Enk, C.D.D. Genomic-scale analysis of psoriatic skin reveals differentially expressed insulin-like growth factor-binding protein-7 after phototherapy. Br. J. Dermatol. 2007, 156, 289–300. [Google Scholar] [CrossRef] [PubMed]
- D’Erme, A.M.; Wilsmann-Theis, D.; Wagenpfeil, J.; Hölzel, M.; Ferring-Schmitt, S.; Sternberg, S.; Wittmann, M.; Peters, B.; Bosio, A.; Bieber, T.; et al. IL-36γ (IL-1F9) Is a Biomarker for Psoriasis Skin Lesions. J. Investig. Dermatol. 2015, 135, 1025–1032. [Google Scholar] [CrossRef] [PubMed]
- Keermann, M.; Kõks, S.; Reimann, E.; Prans, E.; Abram, K.; Kingo, K. Transcriptional landscape of psoriasis identifies the involvement of IL36 and IL36RN. BMC Genom. 2015, 16, 322. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Yi, X.; Ding, Y. Combined Transcriptomic Analysis Revealed AKR1B10 Played an Important Role in Psoriasis through the Dysregulated Lipid Pathway and Overproliferation of Keratinocyte. Biomed. Res. Int. 2017, 2017, 8717369. [Google Scholar] [CrossRef] [PubMed]
- Langley, R.G.; Elewski, B.E.; Lebwohl, M.; Reich, K.; Griffiths, C.E.; Papp, K.; Puig, L.; Nakagawa, H.; Spelman, L.; Sigurgeirsson, B.; et al. Secukinumab in Plaque Psoriasis—Results of Two Phase 3 Trials. N. Engl. J. Med. 2014, 371, 326–338. [Google Scholar] [CrossRef] [PubMed]
- Leonardi, C.L.; Kimball, A.B.; Papp, K.A.; Yeilding, N.; Guzzo, C.; Wang, Y.; Li, S.; Dooley, L.T.; Gordon, K.B. Efficacy and safety of ustekinumab, a human interleukin-12/23 monoclonal antibody, in patients with psoriasis: 76-week results from a randomised, double-blind, placebo-controlled trial (PHOENIX 1). Lancet 2008, 371, 1665–1674. [Google Scholar] [CrossRef]
- Gordon, K.B.; Strober, B.; Lebwohl, M.; Augustin, M.; Blauvelt, A.; Poulin, Y.; Papp, K.A.; Sofen, H.; Puig, L.; Foley, P.; et al. Efficacy and safety of risankizumab in moderate-to-severe plaque psoriasis (UltIMMa-1 and UltIMMa-2): Results from two double-blind, randomised, placebo-controlled and ustekinumab-controlled phase 3 trials. Lancet 2018, 392, 650–661. [Google Scholar] [CrossRef] [PubMed]
- Thomas, S.E.; van den Reek, J.M.P.A.; Seyger, M.M.B.; de Jong, E.M.G.J. How to define a ‘super-responder’ to biologics in psoriasis studies? Br. J. Dermatol. 2023. [CrossRef] [PubMed]
- Reich, K.; Armstrong, A.W.; Langley, R.G.; Flavin, S.; Randazzo, B.; Li, S.; Hsu, M.-C.; Branigan, P.; Blauvelt, A. Guselkumab versus secukinumab for the treatment of moderate-to-severe psoriasis (ECLIPSE): Results from a phase 3, randomised controlled trial. Lancet 2019, 394, 831–839. [Google Scholar] [CrossRef]
- Cook, C.P.; Taylor, M.; Liu, Y.; Schmidt, R.; Sedgewick, A.; Kim, E.; Hailer, A.; North, J.P.; Harirchian, P.; Wang, H.; et al. A single-cell transcriptional gradient in human cutaneous memory T cells restricts Th17/Tc17 identity. Cell Rep. Med. 2022, 3, 100715. [Google Scholar] [CrossRef]
- Zitti, B.; Hoffer, E.; Zheng, W.; Pandey, R.V.; Schlums, H.; Casoni, G.P.; Fusi, I.; Nguyen, L.; Kärner, J.; Kokkinou, E.; et al. Human skin-resident CD8+ T cells require RUNX2 and RUNX3 for induction of cytotoxicity and expression of the integrin CD49a. Immunity 2023, 56, 1285–1302.e7. [Google Scholar] [CrossRef] [PubMed]
- Strobl, J.; Pandey, R.V.; Krausgruber, T.; Bayer, N.; Kleissl, L.; Reininger, B.; Vieyra-Garcia, P.; Wolf, P.; Jentus, M.-M.; Mitterbauer, M.; et al. Long-term skin-resident memory T cells proliferate in situ and are involved in human graft-versus-host disease. Sci. Transl. Med. 2020, 12, eabb7028. [Google Scholar] [CrossRef]
- Vu, T.T.; Koguchi-Yoshioka, H.; Watanabe, R. Skin-Resident Memory T Cells: Pathogenesis and Implication for the Treatment of Psoriasis. J. Clin. Med. 2021, 10, 3822. [Google Scholar] [CrossRef]
- Wienke, J.; Veldkamp, S.R.; Struijf, E.M.; Yengej, F.A.Y.; van der Wal, M.M.; van Royen-Kerkhof, A.; van Wijk, F. T cell interaction with activated endothelial cells primes for tissue-residency. Front. Immunol. 2022, 13, 5133. [Google Scholar] [CrossRef] [PubMed]
- Kasprowicz-Furmańczyk, M.; Czerwińska, J.; Placek, W.; Owczarczyk-Saczonek, A. Assessment of the Tissue Resident Memory Cells in Lesional Skin of Patients with Psoriasis and in Healthy Skin of Healthy Volunteers. Int. J. Environ. Res. Public Health 2021, 18, 11251. [Google Scholar] [CrossRef] [PubMed]
- Kurihara, K.; Fujiyama, T.; Phadungsaksawasdi, P.; Ito, T.; Tokura, Y. Significance of IL-17A-producing CD8+CD103+ skin resident memory T cells in psoriasis lesion and their possible relationship to clinical course. J. Dermatol. Sci. 2019, 95, 21–27. [Google Scholar] [CrossRef]
- Fenix, K.; Wijesundara, D.K.; Cowin, A.J.; Grubor-Bauk, B.; Kopecki, Z. Immunological Memory in Imiquimod-Induced Murine Model of Psoriasiform Dermatitis. Int. J. Mol. Sci. 2020, 21, 7228. [Google Scholar] [CrossRef] [PubMed]
- Munn, D.H.; Shafizadeh, E.; Attwood, J.T.; Bondarev, I.; Pashine, A.; Mellor, A.L. Inhibition of T Cell Proliferation by Macrophage Tryptophan Catabolism. J. Exp. Med. 1999, 189, 1363–1372. [Google Scholar] [CrossRef] [PubMed]
- He, S.; Bhatt, R.; Brown, C.; Brown, E.A.; Buhr, D.L.; Chantranuvatana, K.; Danaher, P.; Dunaway, D.; Garrison, R.G.; Geiss, G.; et al. High-plex imaging of RNA and proteins at subcellular resolution in fixed tissue by spatial molecular imaging. Nat. Biotechnol. 2022, 40, 1794–1806. [Google Scholar] [CrossRef]
- Ben Abdallah, H.; Emmanuel, T.; Bregnhøj, A.; Johansen, C.; Iversen, L. Early intervention and disease memory in psoriasis: A literature review. JEADV Clin. Pract. 2022, 1, 307–316. [Google Scholar] [CrossRef]
- Bellinato, F.; Chiricozzi, A.; Piaserico, S.; Targher, G.; Gisondi, P. Could Targeted Pharmacotherapies Exert a “Disease Modification Effect” in Patients with Chronic Plaque Psoriasis? Int. J. Mol. Sci. 2022, 23, 12849. [Google Scholar] [CrossRef] [PubMed]
- Lebwohl, M.; Iversen, L.; Eidsmo, L.; Krueger, J.G.; Suárez-Fariñas, M.; Tomalin, L.; Kolbinger, F.; You, R.; Milutinovic, M. Investigation of plaque psoriasis relapse after secukinumab withdrawal in patients from two Phase 3 studies. Clin. Exp. Dermatol. 2023, llad329. [Google Scholar] [CrossRef] [PubMed]
- Bankhead, P.; Loughrey, M.B.; Fernández, J.A.; Dombrowski, Y.; McArt, D.G.; Dunne, P.D.; McQuaid, S.; Gray, R.T.; Murray, L.J.; Coleman, H.G.; et al. QuPath: Open source software for digital pathology image analysis. Sci. Rep. 2017, 7, 16878. [Google Scholar] [CrossRef]
- Emmanuel, T.; Brent, M.B.; Iversen, L.; Johansen, C. Quantification of Immunohistochemically Stained Cells in Skin Biopsies. Dermatopathology 2022, 9, 82–93. [Google Scholar] [CrossRef] [PubMed]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Emmanuel, T.; Ignatov, B.; Bertelsen, T.; Litman, T.; Nielsen, M.M.; Brent, M.B.; Touborg, T.; Rønsholdt, A.B.; Petersen, A.; Boye, M.; et al. Secukinumab and Dead Sea Climatotherapy Impact Resolved Psoriasis Skin Differently Potentially Affecting Disease Memory. Int. J. Mol. Sci. 2024, 25, 6086. https://doi.org/10.3390/ijms25116086
Emmanuel T, Ignatov B, Bertelsen T, Litman T, Nielsen MM, Brent MB, Touborg T, Rønsholdt AB, Petersen A, Boye M, et al. Secukinumab and Dead Sea Climatotherapy Impact Resolved Psoriasis Skin Differently Potentially Affecting Disease Memory. International Journal of Molecular Sciences. 2024; 25(11):6086. https://doi.org/10.3390/ijms25116086
Chicago/Turabian StyleEmmanuel, Thomas, Borislav Ignatov, Trine Bertelsen, Thomas Litman, Morten Muhlig Nielsen, Mikkel Bo Brent, Toke Touborg, Anders Benjamin Rønsholdt, Annita Petersen, Mette Boye, and et al. 2024. "Secukinumab and Dead Sea Climatotherapy Impact Resolved Psoriasis Skin Differently Potentially Affecting Disease Memory" International Journal of Molecular Sciences 25, no. 11: 6086. https://doi.org/10.3390/ijms25116086
APA StyleEmmanuel, T., Ignatov, B., Bertelsen, T., Litman, T., Nielsen, M. M., Brent, M. B., Touborg, T., Rønsholdt, A. B., Petersen, A., Boye, M., Kaaber, I., Sortebech, D., Lybæk, D., Steiniche, T., Bregnhøj, A., Eidsmo, L., Iversen, L., & Johansen, C. (2024). Secukinumab and Dead Sea Climatotherapy Impact Resolved Psoriasis Skin Differently Potentially Affecting Disease Memory. International Journal of Molecular Sciences, 25(11), 6086. https://doi.org/10.3390/ijms25116086