
Genetic Alterations of SMYD4 in Solid Tumors Using Integrative Multi-Platform Analysis

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Mutation Profile of SMYD Genes
2.2. Copy Number Alterations in the SMYD Family Genes
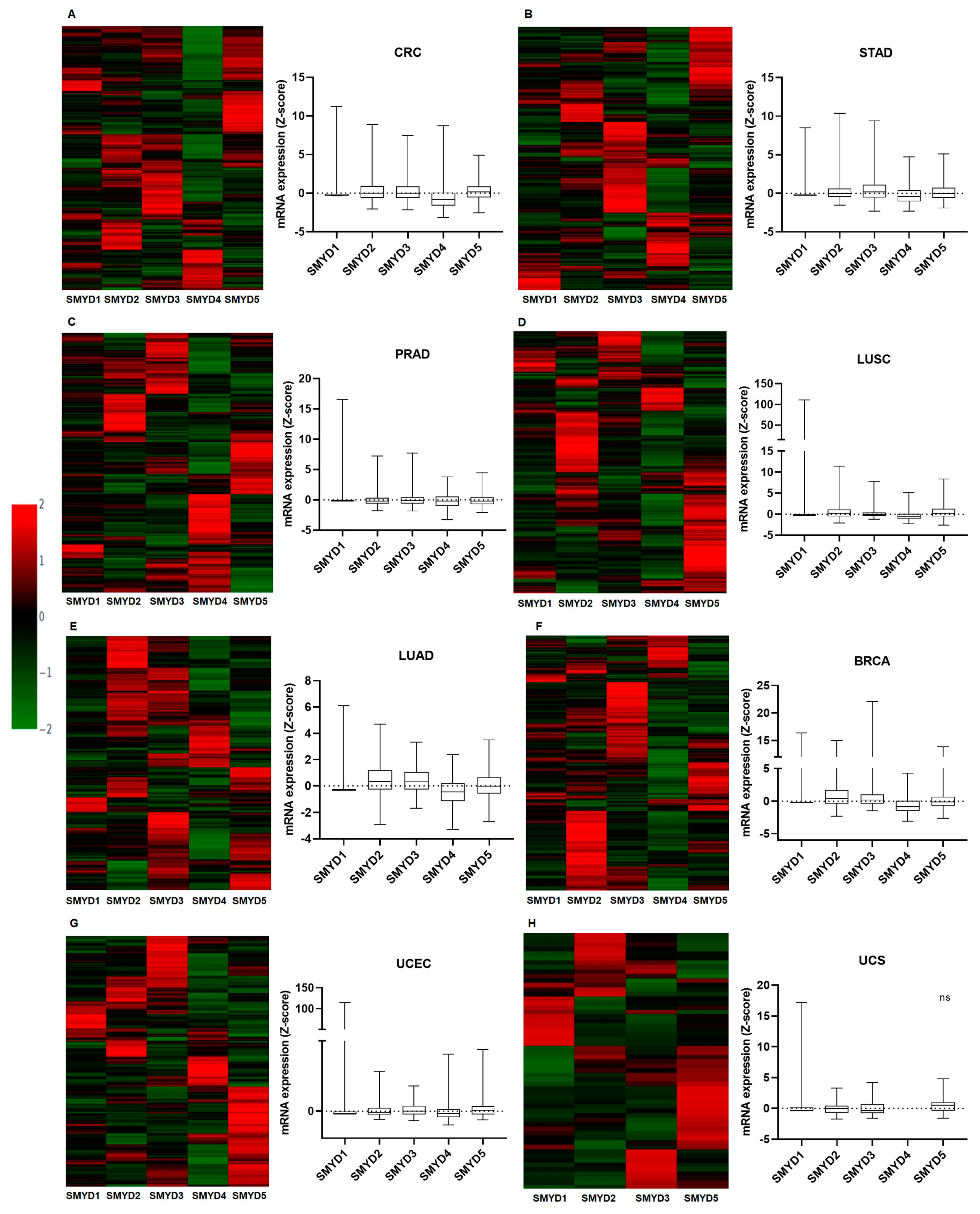
2.3. mRNA Expression Profile
2.4. Correlation Analysis of SMYD4 Expression with Other SMYD Genes
2.5. Comparison of SMYD4 Expression between Normal Samples and Tumor Samples at Different Tumor Stages
2.6. Correlation Analyses between SMYD4 CNAs and mRNA Expression
2.7. Overall Patient Survival Based on SMYD4 Expression
3. Discussion
4. Materials and Methods
4.1. Gene Database Analysis
4.2. Heatmap
4.3. Survival Curve
4.4. Statistics
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Albert, M.; Helin, K. Histone methyltransferases in cancer. Semin. Cell Dev. Biol. 2010, 21, 209–220. [Google Scholar] [CrossRef]
- Leinhart, K.; Brown, M. SET/MYND Lysine Methyltransferases Regulate Gene Transcription and Protein Activity. Genes 2011, 2, 210–218. [Google Scholar] [CrossRef]
- Liu, D.; Wang, X.; Shi, E.; Wang, L.; Nie, M.; Li, L.; Jiang, Q.; Kong, P.; Shi, S.; Wang, C.; et al. Comprehensive Analysis of the Value of SMYD Family Members in the Prognosis and Immune Infiltration of Malignant Digestive System Tumors. Front. Genet. 2021, 12, 699910. [Google Scholar] [CrossRef]
- Hu, L.; Zhu, Y.T.; Qi, C.; Zhu, Y.J. Identification of Smyd4 as a potential tumor suppressor gene involved in breast cancer development. Cancer Res. 2009, 69, 4067–4072. [Google Scholar] [CrossRef]
- Han, S.; Zou, H.; Lee, J.W.; Han, J.; Kim, H.C.; Cheol, J.J.; Kim, L.S.; Kim, H. miR-1307-3p Stimulates Breast Cancer Development and Progression by Targeting SMYD4. J. Cancer 2019, 10, 441–448. [Google Scholar] [CrossRef]
- Aziz, N.; Hong, Y.H.; Kim, H.G.; Kim, J.H.; Cho, J.Y. Tumor-suppressive functions of protein lysine methyltransferases. Exp. Mol. Med. 2023, 55, 2475–2497. [Google Scholar] [CrossRef]
- Liu, S.; Cheng, K.; Zhang, H.; Kong, R.; Wang, S.; Mao, C.; Liu, S. Methylation Status of the Nanog Promoter Determines the Switch between Cancer Cells and Cancer Stem Cells. Adv. Sci. 2020, 7, 1903035. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA Cancer J Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Kastenhuber, E.R.; Lowe, S.W. Putting p53 in Context. Cell 2017, 170, 1062–1078. [Google Scholar] [CrossRef]
- Albertson, D.G.; Collins, C.; McCormick, F.; Gray, J.W. Chromosome aberrations in solid tumors. Nat. Genet. 2003, 34, 369–376. [Google Scholar] [CrossRef]
- Mermel, C.H.; Schumacher, S.E.; Hill, B.; Meyerson, M.L.; Beroukhim, R.; Getz, G. GISTIC2.0 facilitates sensitive and confident localization of the targets of focal somatic copy-number alteration in human cancers. Genome Biol. 2011, 12, R41. [Google Scholar] [CrossRef]
- Lánczky, A.; Győrffy, B. Web-Based Survival Analysis Tool Tailored for Medical Research (KMplot): Development and Implementation. J. Med. Internet Res. 2021, 23, e27633. [Google Scholar] [CrossRef]
- Jenal, M.; Britschgi, C.; Fey, M.F.; Tschan, M.P. Inactivation of the hypermethylated in cancer 1 tumour suppressor--not just a question of promoter hypermethylation? Swiss Med. Wkly. 2010, 140, w13106. [Google Scholar] [CrossRef]
- Wu, G.J.; Shan, X.N.; Li, M.F.; Shi, S.L.; Zheng, Q.P.; Yu, L.; Zhao, S.Y. Preliminary study on the loss of heterozygosity at 17p13 in gastric and colorectal cancers. World J. Gastroenterol. 1997, 3, 160–162. [Google Scholar] [CrossRef]
- Tsuchiya, E.; Tanigami, A.; Ishikawa, Y.; Nishida, K.; Hayashi, M.; Tokuchi, Y.; Hashimoto, T.; Okumura, S.; Tsuchiya, S.; Nakagawa, K. Three New Regions on Chromosome 17p13.3 Distal to p53 with Possible Tumor Suppressor Gene Involvement in Lung Cancer. Jpn. J. Cancer Res. 2000, 91, 589–596. [Google Scholar] [CrossRef]
- Park, S.Y.; Kang, Y.S.; Kim, B.G.; Lee, S.H.; Lee, E.D.; Lee, K.H.; Park, K.B.; Lee, J.H. Loss of heterozygosity on the short arm of chromosome 17 in uterine cervical carcinomas. Cancer Genet. Cytogenet. 1995, 79, 74–78. [Google Scholar] [CrossRef]
- Bhattacharya, A.; Bense, R.D.; Urzúa-Traslaviña, C.G.; de Vries, E.G.; van Vugt, M.A.; Fehrmann, R.S. Transcriptional effects of copy number alterations in a large set of human cancers. Nat. Commun. 2020, 11, 715. [Google Scholar] [CrossRef]
- Xiao, D.; Wang, H.; Hao, L.; Guo, X.; Ma, X.; Qian, Y.; Chen, H.; Ma, J.; Zhang, J.; Sheng, W.; et al. The roles of SMYD4 in epigenetic regulation of cardiac development in zebrafish. PLoS Genet. 2018, 14, e1007578. [Google Scholar] [CrossRef]
- Chen, K.; Chen, Z.; Wu, D.; Zhang, L.; Lin, X.; Su, J.; Rodriguez, B.; Xi, Y.; Xia, Z.; Chen, X.; et al. Broad H3K4me3 is associated with increased transcription elongation and enhancer activity at tumor-suppressor genes. Nat. Genet. 2015, 47, 1149–1157. [Google Scholar] [CrossRef]
- Zhang, Y.; Yan, L.; Yao, W.; Chen, K.; Xu, H.; Ye, Z. Integrated Analysis of Genetic Abnormalities of the Histone Lysine Methyltransferases in Prostate Cancer. Med. Sci. Monit. 2019, 25, 193–239. [Google Scholar] [CrossRef]
- Lin, T.; Ding, Y.Q.; Li, J.M. Overexpression of Nanog protein is associated with poor prognosis in gastric adenocarcinoma. Med. Oncol. 2012, 29, 878–885. [Google Scholar] [CrossRef]
- Ma, X.; Wang, B.; Wang, X.; Luo, Y.; Fan, W. NANOGP8 is the key regulator of stemness, EMT, Wnt pathway, chemoresistance, and other malignant phenotypes in gastric cancer cells. PLoS ONE 2018, 13, e0192436. [Google Scholar] [CrossRef]
- Zhou, Z.; Chen, Z.; Zhou, Q.; Meng, S.; Shi, J.; Mui, S.; Jiang, H.; Lin, J.; He, G.; Li, W.; et al. SMYD4 monomethylates PRMT5 and forms a positive feedback loop to promote hepatocellular carcinoma progression. Cancer Sci. 2024, 115, 1587–1601. [Google Scholar] [CrossRef]
- Hamamoto, R.; Furukawa, Y.; Morita, M.; Iimura, Y.; Silva, F.P.; Li, M.; Yagyu, R.; Nakamura, Y. SMYD3 encodes a histone methyltransferase involved in the proliferation of cancer cells. Nat. Cell Biol. 2004, 6, 731–740. [Google Scholar] [CrossRef]
- Liu, N.; Sun, S.; Yang, X. Prognostic significance of stromal SMYD3 expression in colorectal cancer of TNM stage I-III. Int J Clin Exp Pathol. 2017, 10, 8901–8907. [Google Scholar]
- Bernard, B.J.; Nigam, N.; Burkitt, K.; Saloura, V. SMYD3: A regulator of epigenetic and signaling pathways in cancer. Clin. Epigenetics 2021, 13, 45. [Google Scholar] [CrossRef]
- Bottino, C.; Peserico, A.; Simone, C.; Caretti, G. SMYD3: An Oncogenic Driver Targeting Epigenetic Regulation and Signaling Pathways. Cancers 2020, 12, 142. [Google Scholar] [CrossRef]
- Ikram, S.; Rege, A.; Negesse, M.Y.; Casanova, A.G.; Reynoird, N.; Green, E.M. The SMYD3-MAP3K2 Signaling Axis Promotes Tumor Aggressiveness and Metastasis in Prostate Cancer. Sci. Adv. 2023, 9, eadi5921. Available online: https://www.science.org/doi/10.1126/sciadv.adi5921 (accessed on 28 April 2024). [CrossRef]
- Hamamoto, R.; Silva, F.P.; Tsuge, M.; Nishidate, T.; Katagiri, T.; Nakamura, Y.; Furukawa, Y. Enhanced SMYD3 expression is essential for the growth of breast cancer cells. Cancer Sci. 2006, 97, 113–118. [Google Scholar] [CrossRef]
- Fenizia, C.; Bottino, C.; Corbetta, S.; Fittipaldi, R.; Floris, P.; Gaudenzi, G.; Carra, S.; Cotelli, F.; Vitale, G.; Caretti, G. SMYD3 promotes the epithelial-mesenchymal transition in breast cancer. Nucleic Acids Res. 2019, 47, 1278–1293. [Google Scholar] [CrossRef]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [PubMed]
- Chandrashekar, D.S.; Karthikeyan, S.K.; Korla, P.K.; Patel, H.; Shovon, A.R.; Athar, M.; Netto, G.J.; Qin, Z.S.; Kumar, S.; Manne, U.; et al. UALCAN: An update to the integrated cancer data analysis platform. Neoplasia 2022, 25, 18–27. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Tumor | SMYD1 | SMYD2 | SMYD3 | SMYD4 | SMYD5 |
|---|---|---|---|---|---|
| CRC | 1.4% | 0.9% | 1.0% | 0.5% | 2.3% |
| STAD | 1.2% | 0.9% | 1.9% | 0,8% | 1.1% |
| BRCA | 0.3% | 0.2% | 0.4% | 0.4% | 0.3% |
| PRAD | 0.2% | 0.1% | 0.2% | 0.1% | 0.1% |
| Lung Cancer | |||||
| LUAD | 2.3% | 0.8% | 0.7% | 0.4% | 0.5% |
| LUSC | 2.3% | 0.8% | 1.2% | 0.8% | 0.4% |
| Uterine Cancer | |||||
| UCEC | 3.3% | 2.1% | 2.1% | 2.1% | 2.4% |
| UCS | 5.1% | 0.0% | 0.0% | 2.5% | 1.3% |
| Tumors | Pearson | p Value | Spearman | p Value | Patients (n) |
|---|---|---|---|---|---|
| CRC | 0.568 | <0.0001 | 0.337 | <0.0001 | 255 |
| STAD | 0.593 | <0.0001 | 0.465 | <0.0001 | 190 |
| PRAD | 0.468 | <0.0001 | 0.296 | 0.0015 | 98 |
| LUSC | 0.615 | <0.0001 | 0.500 | <0.0001 | 362 |
| LUAD | 0.488 | <0.0001 | 0.354 | 0.0003 | 89 |
| BRCA | 0.532 | <0.0001 | 0.472 | <0.0001 | 695 |
| UCEC | 0.483 | <0.0001 | 0.538 | <0.0001 | 74 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olivera Santana, B.L.; de Loyola, M.B.; Gualberto, A.C.M.; Pittella-Silva, F. Genetic Alterations of SMYD4 in Solid Tumors Using Integrative Multi-Platform Analysis. Int. J. Mol. Sci. 2024, 25, 6097. https://doi.org/10.3390/ijms25116097
Olivera Santana BL, de Loyola MB, Gualberto ACM, Pittella-Silva F. Genetic Alterations of SMYD4 in Solid Tumors Using Integrative Multi-Platform Analysis. International Journal of Molecular Sciences. 2024; 25(11):6097. https://doi.org/10.3390/ijms25116097
Chicago/Turabian StyleOlivera Santana, Brunna Letícia, Mariana Braccialli de Loyola, Ana Cristina Moura Gualberto, and Fabio Pittella-Silva. 2024. "Genetic Alterations of SMYD4 in Solid Tumors Using Integrative Multi-Platform Analysis" International Journal of Molecular Sciences 25, no. 11: 6097. https://doi.org/10.3390/ijms25116097