

Genome-Wide Transcriptomic and Metabolomic Analyses Unveiling the Defence Mechanisms of Populus tremula against Sucking and Chewing Insect Herbivores
 , , , ,
, , , ,  and
and
Abstract
:1. Introduction
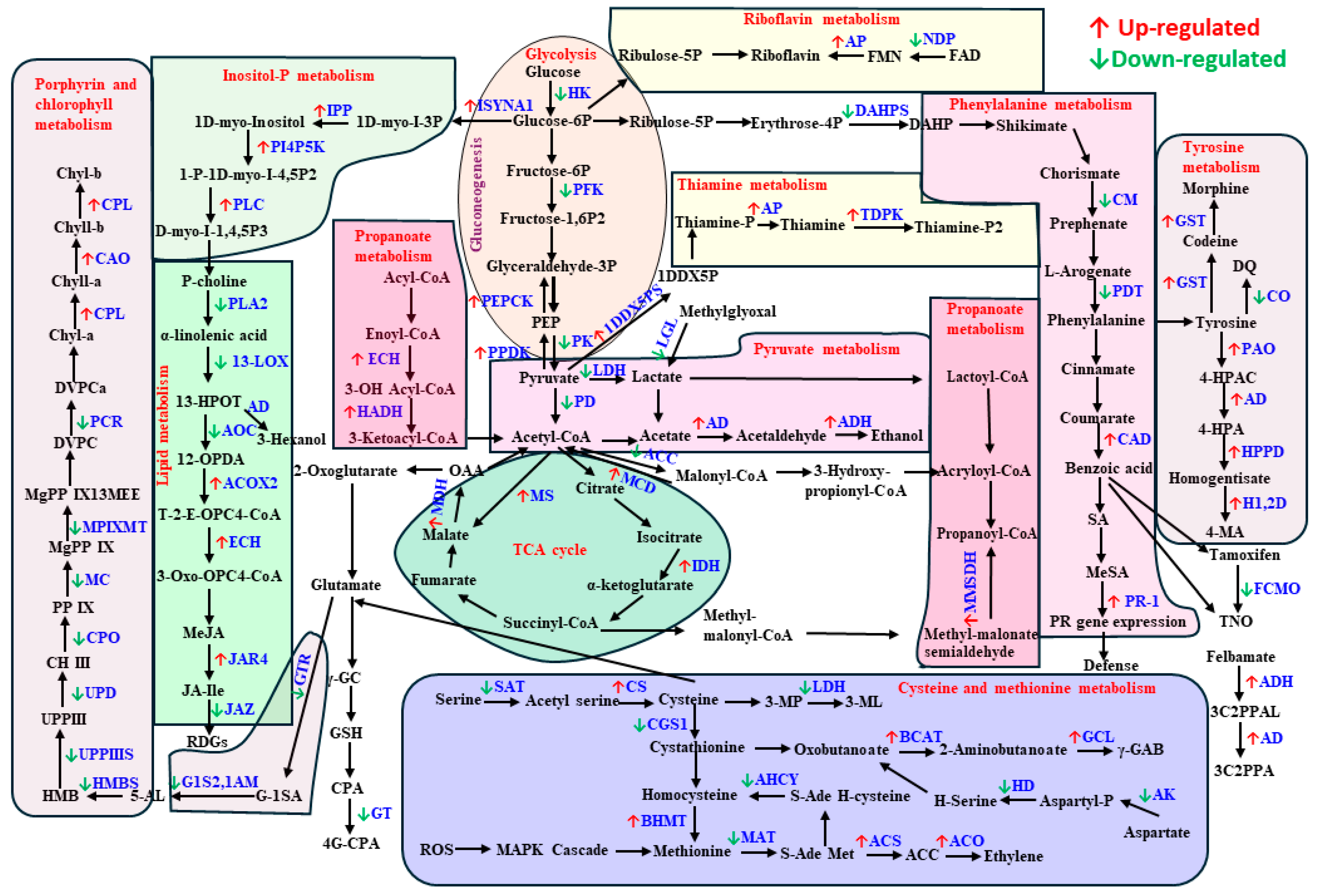
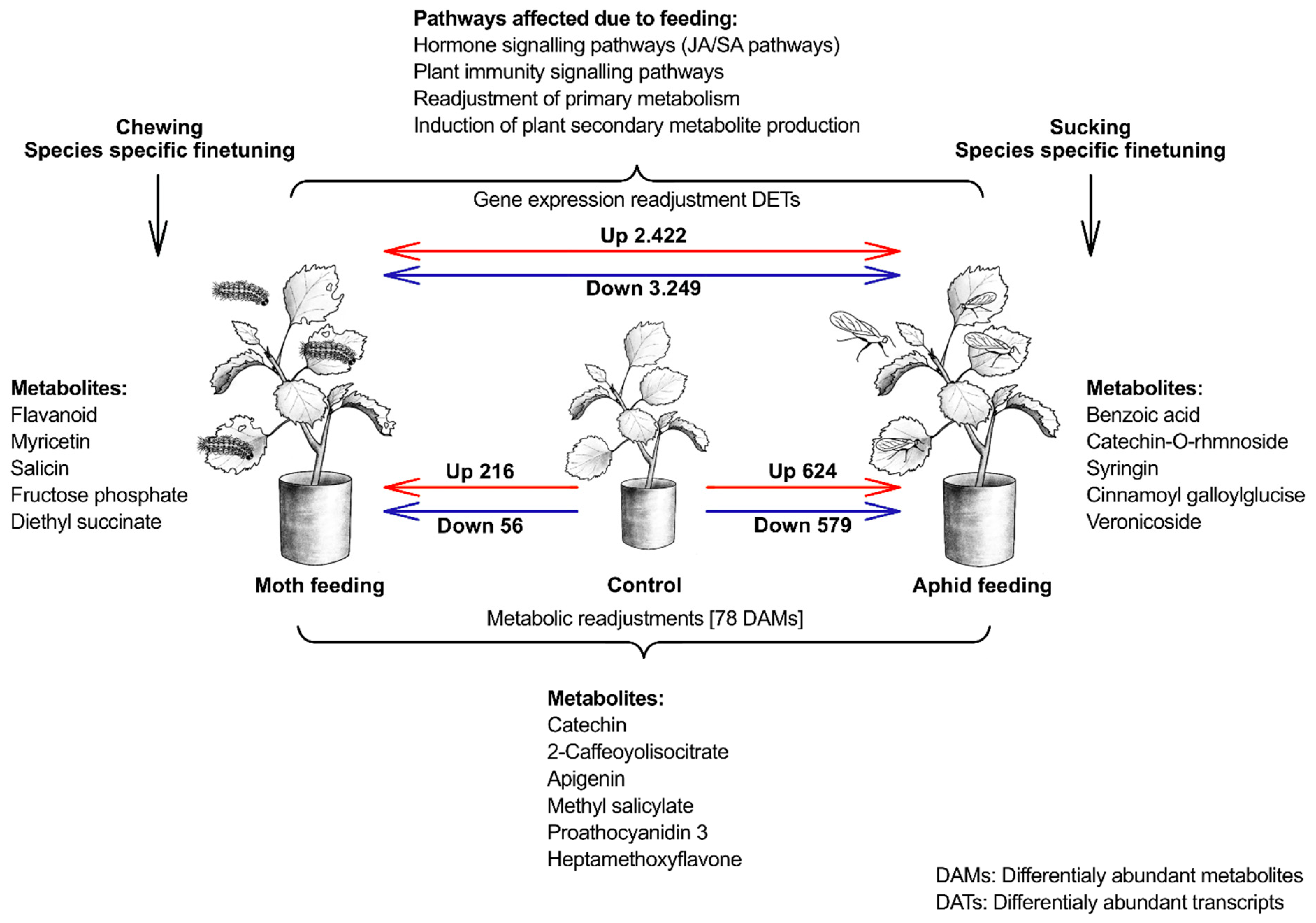
2. Results
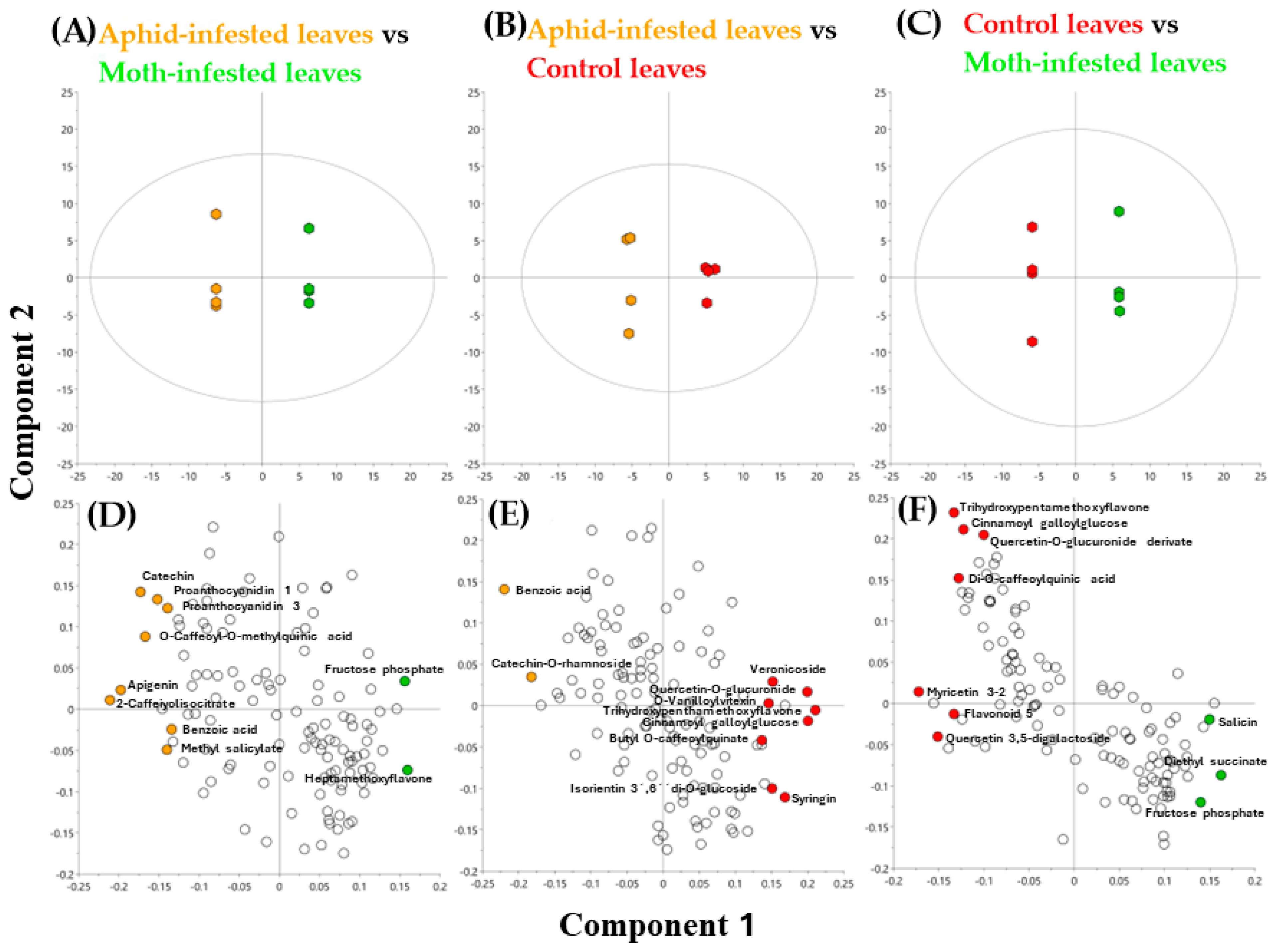
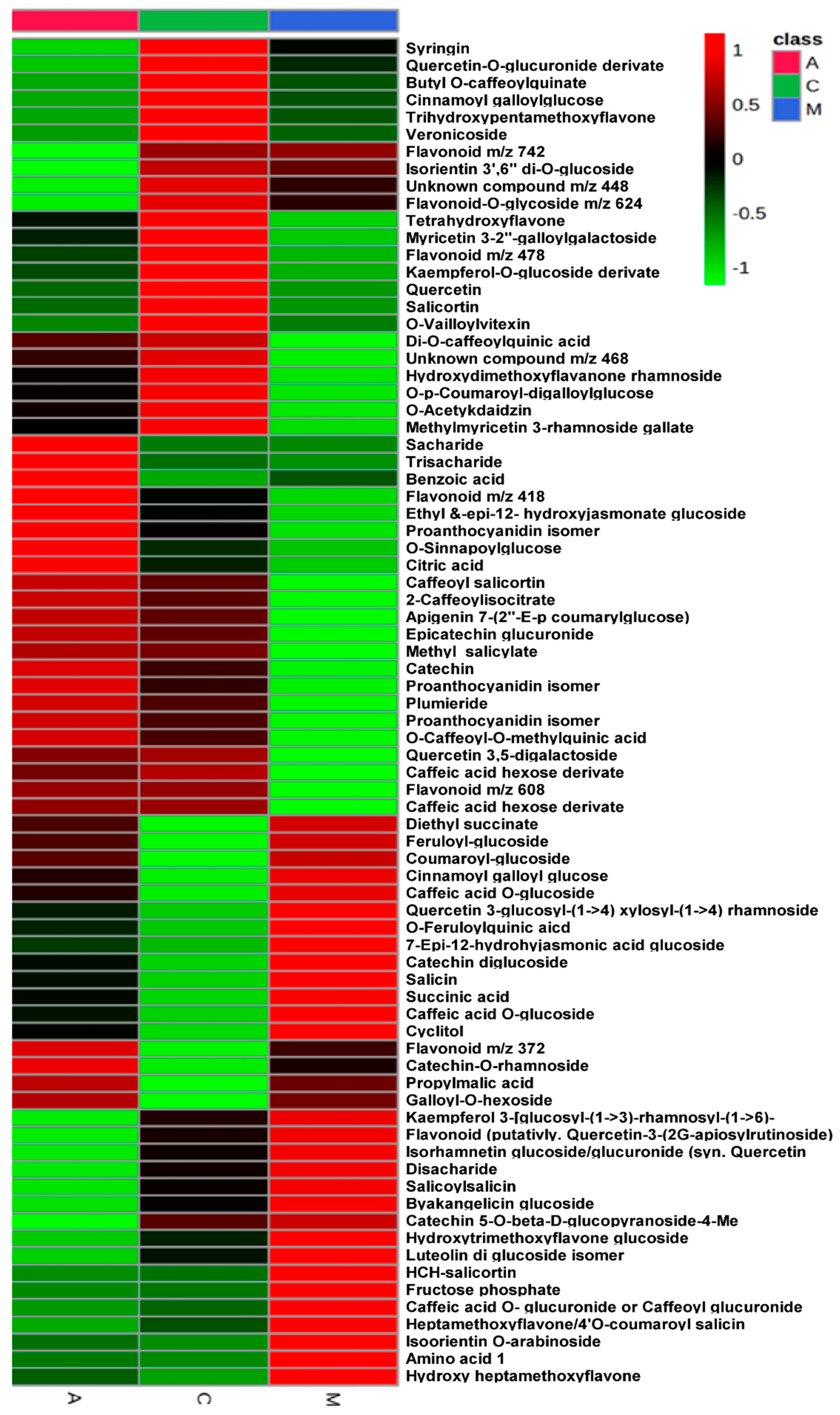
2.1. Metabolomic Analysis
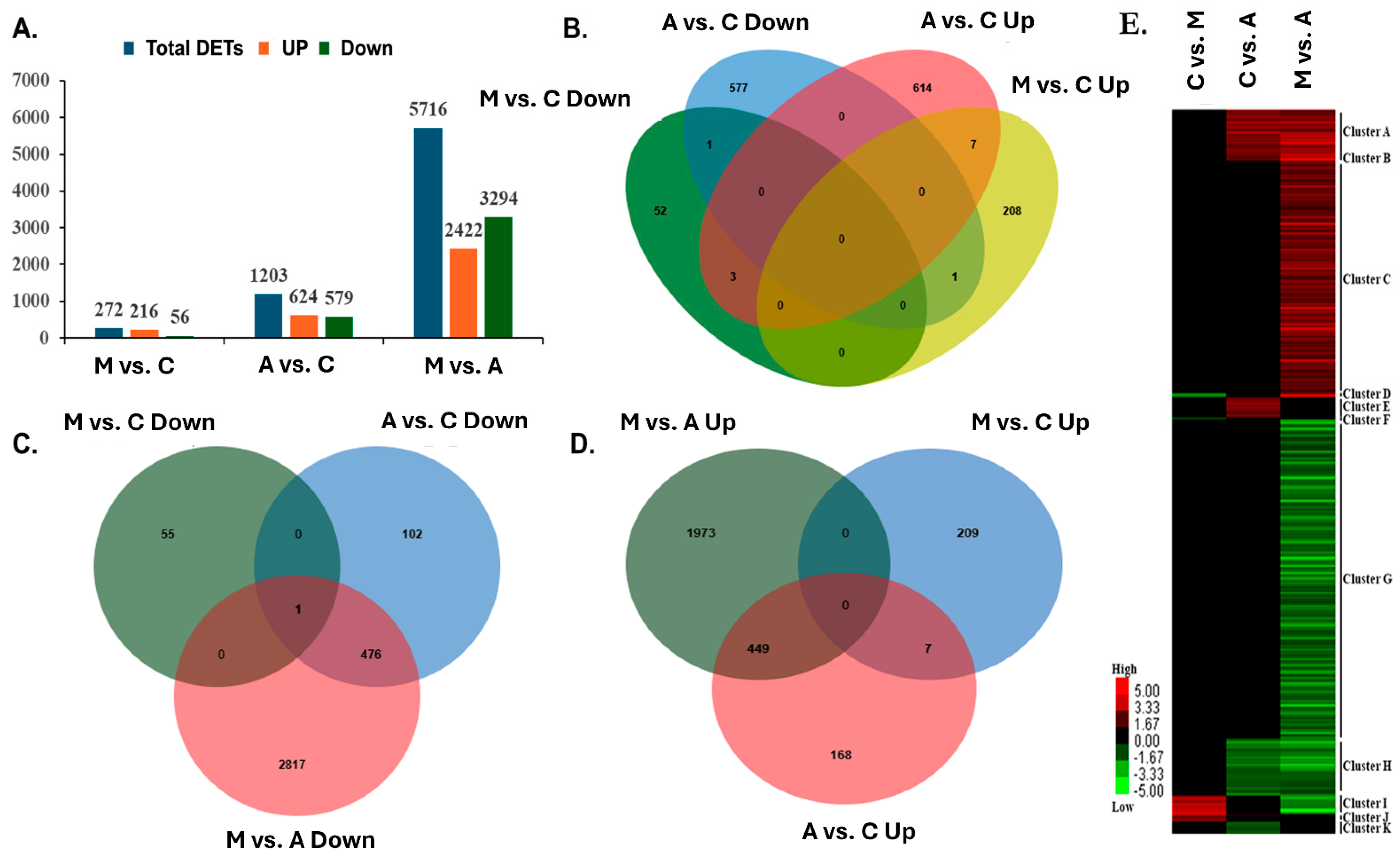
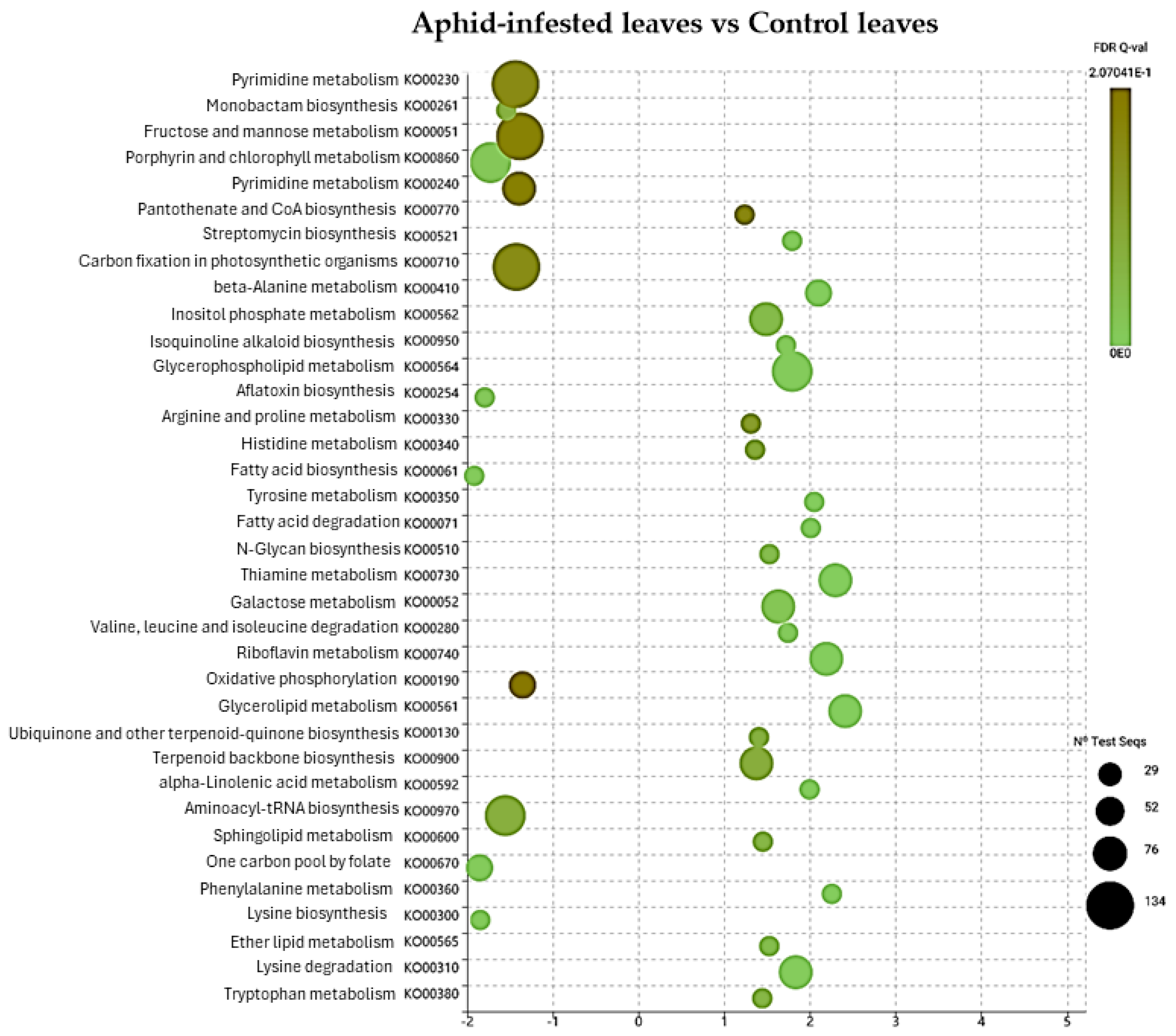
2.2. DGE Analysis on Poplar Leaves Infested by Sucking and Chewing Pests
2.3. Reference Gene Selection
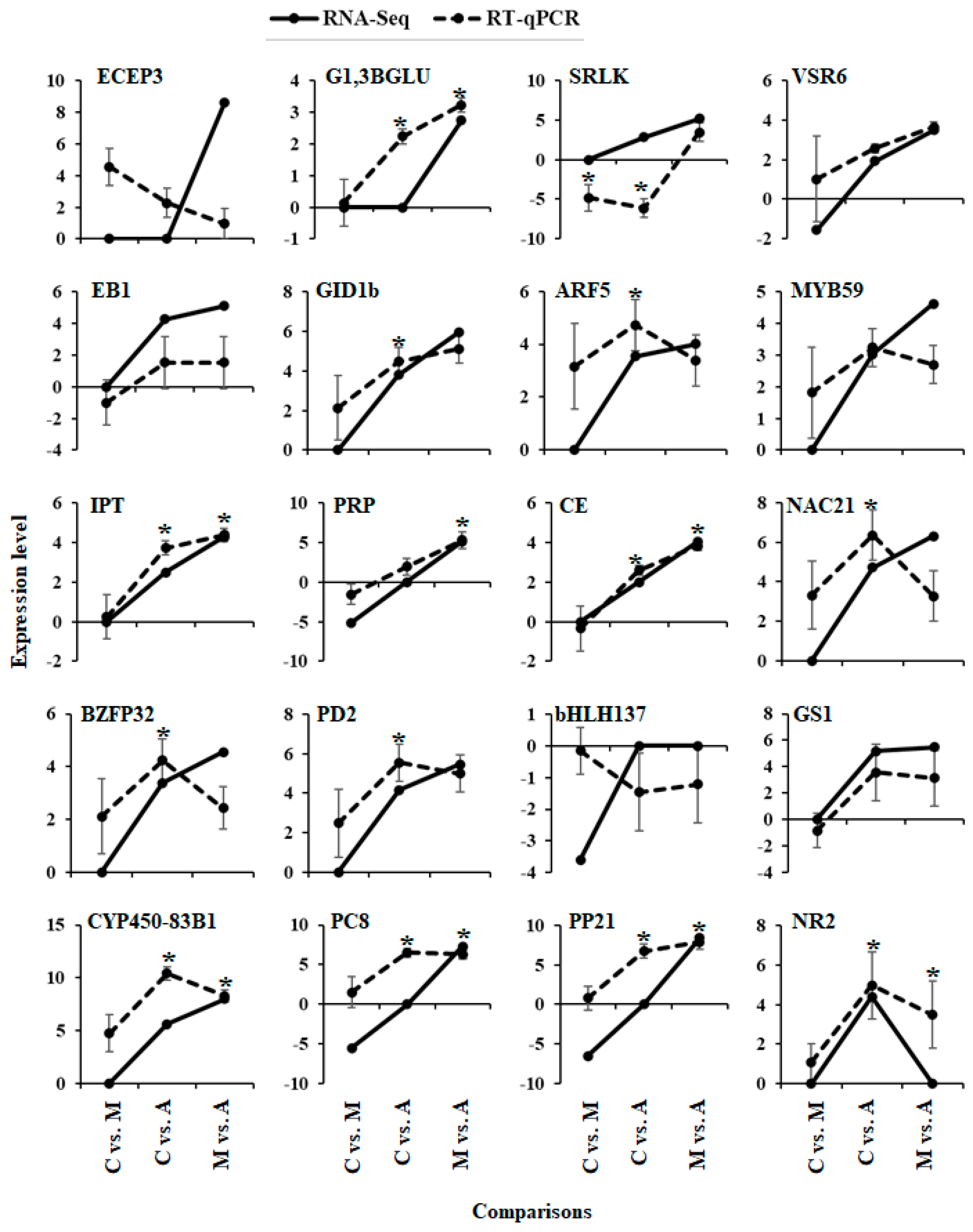
2.4. Gene Expression Validation
3. Discussion
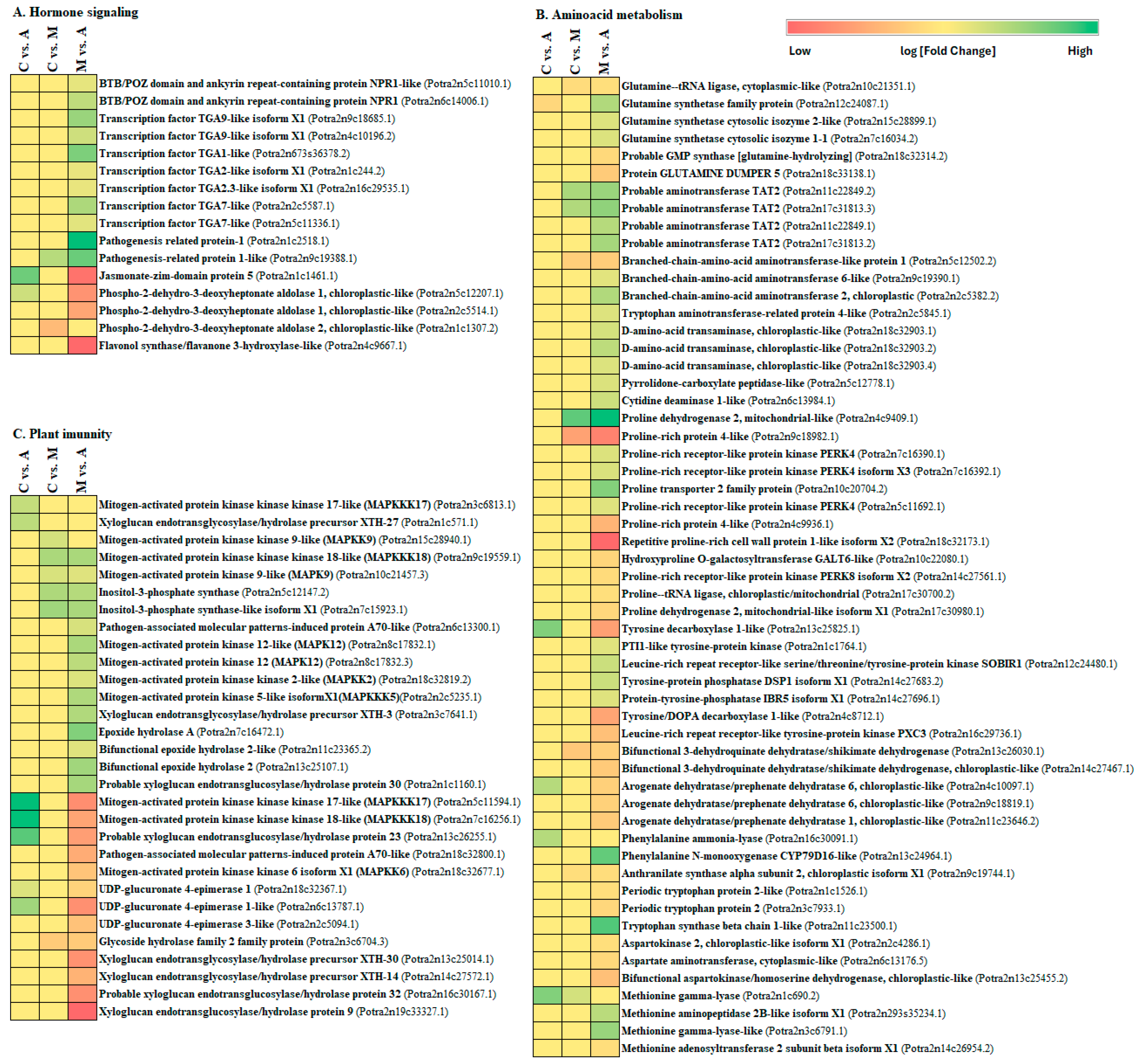
3.1. Response of Hormones Signalling Pathways after Aphid and Spongy Moth Infestation
3.2. Plant Immune Defence against Aphid and Spongy Moth Infestation
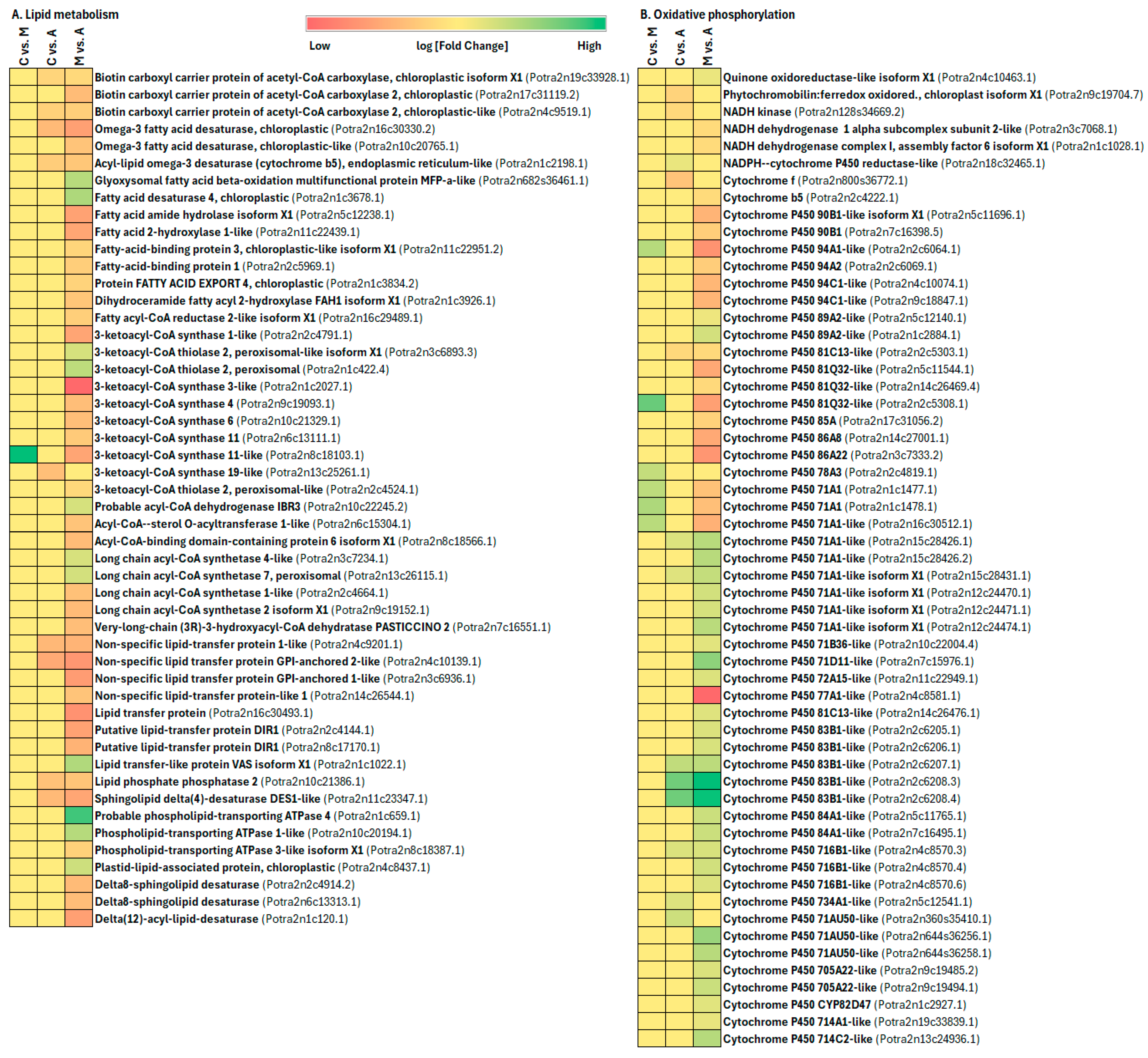
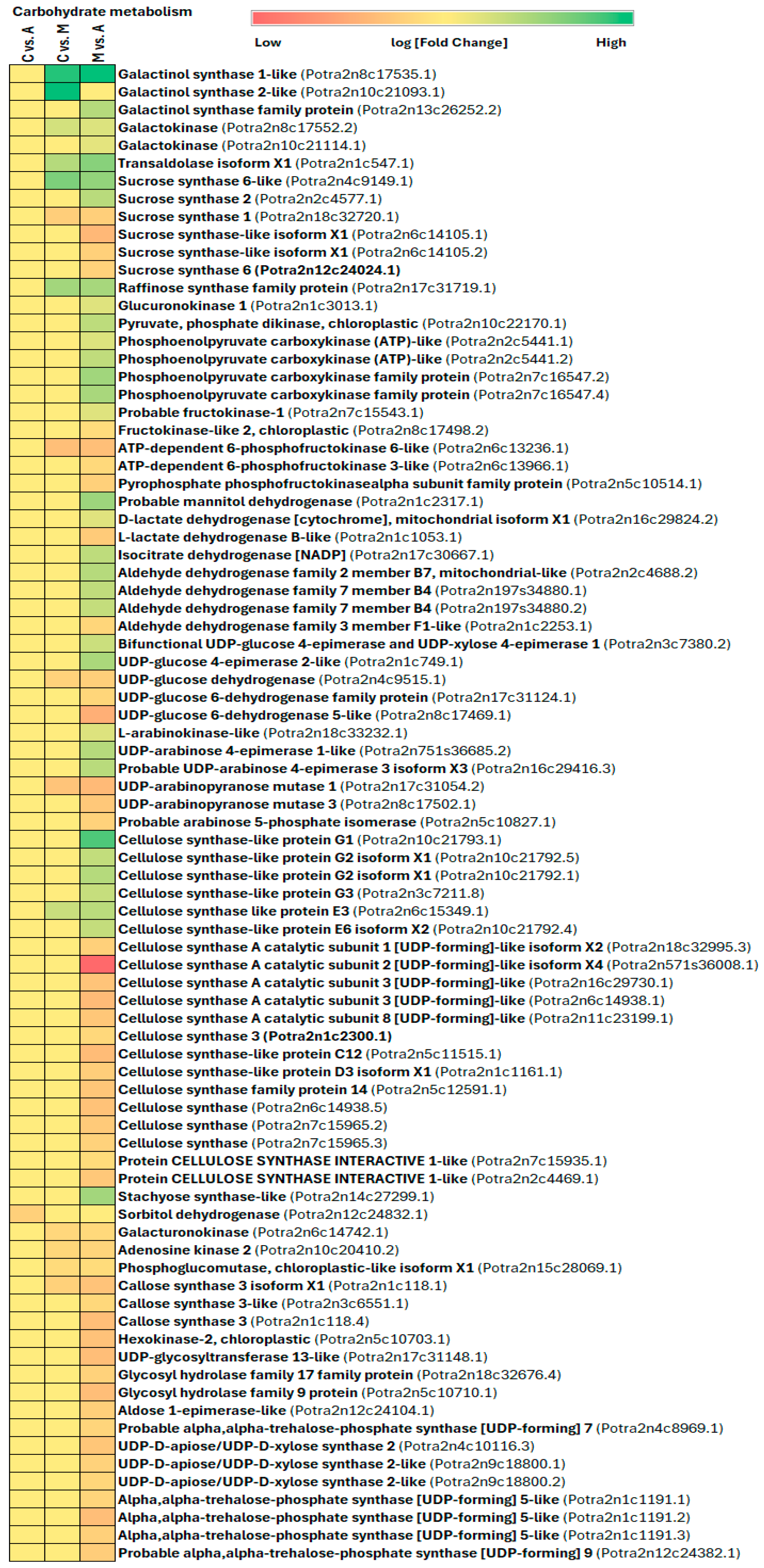
3.3. Primary Metabolism Alteration after Aphid and Spongy Moth Infestation
3.4. Plant Secondary Metabolism Alteration Due to Aphid and Spongy Moth Infestation
4. Materials and Methods
4.1. Plants and Insects
4.2. Experimental Design
4.3. Poplar Tissue Feeding, Collection and Processing
4.4. Metabolomics Non-Targeted Analysis
4.4.1. Extraction Procedure
4.4.2. LC-MS-qTOF Metabolomic Analysis
4.4.3. Statistical Evaluation of LC-MS-qTOF Data
4.5. Transcriptomics Analysis
4.5.1. Total RNA Isolation
4.5.2. NGS Sequencing and Data Analysis
4.5.3. Reference Gene Selection for RT-qPCR
4.5.4. Gene Expression Validation by RT-qPCR
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gatehouse, J.A. Plant resistance towards insect herbivores: A dynamic interaction. New Phytol. 2002, 156, 145–169. [Google Scholar] [CrossRef] [PubMed]
- Zunjarrao, S.S.; Tellis, M.B.; Joshi, S.N.; Joshi, R.S. Plant-insect interaction: The saga of molecular coevolution. In Co-Evolution of Secondary Metabolites; Springer: Berlin/Heidelberg, Germany, 2020; pp. 19–45. [Google Scholar] [CrossRef]
- Howe, G.A.; Jander, G. Plant immunity to insect herbivores. Annu. Rev. Plant Biol. 2008, 59, 41–66. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Kou, J.; Jing, W.; Han, X.; Liu, D.; Ghasemzadeh, S.; Sun, P.; Shi, W.; Zhang, Y. Transcriptomic and metabolomic reprogramming in cotton after Apolygus lucorum feeding implicated in enhancing recruitment of the parasitoid Peristenus spretus. J. Pest Sci. 2022, 95, 249–262. [Google Scholar] [CrossRef]
- Kumar, S.; Kanakachari, M.; Gurusamy, D.; Kumar, K.; Narayanasamy, P.; Kethireddy Venkata, P.; Solanke, A.; Gamanagatti, S.; Hiremath, V.; Katageri, I.S.; et al. Genome-wide transcriptomic and proteomic analyses of bollworm-infested developing cotton bolls revealed the genes and pathways involved in the insect pest defence mechanism. Plant Biotechnol. J. 2016, 14, 1438–1455. [Google Scholar] [CrossRef] [PubMed]
- Roy, A.; Chakraborty, A. Natural Insecticidal Proteins and Their Potential in Future IPM. In Plant-Pest Interactions: From Molecular Mechanisms to Chemical Ecology: Chemical Ecology; Springer: Berlin/Heidelberg, Germany, 2021; pp. 265–303. [Google Scholar]
- Singh, A.; Mehta, S.; Yadav, S.; Nagar, G.; Ghosh, R.; Roy, A.; Chakraborty, A.; Singh, I.K. How to Cope with the Challenges of Environmental Stresses in the Era of Global Climate Change: An Update on ROS Stave off in Plants. Int. J. Mol. Sci. 2022, 23, 1995. [Google Scholar] [CrossRef] [PubMed]
- Sumanti, G.; Roy, A. Plant Cell Wall: A Simple Physical Barrier or a Complex Defense Modulator–Exploring Its Dynamic Role at Plant-Fungus Interface. In Molecular Aspects of Plant-Pathogen Interaction; Springer: Berlin/Heidelberg, Germany, 2018; pp. 333–351. [Google Scholar]
- War, A.R.; Paulraj, M.G.; Ahmad, T.; Buhroo, A.A.; Hussain, B.; Ignacimuthu, S.; Sharma, H.C. Mechanisms of plant defense against insect herbivores. Plant Signal. Behav. 2012, 7, 1306–1320. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chen, Q.; Tan, Y.; Shuang, S.; Dai, R.; Jiang, X.; Temuer, B. Combined transcriptome and metabolome analysis of alfalfa response to thrips infection. Genes 2021, 12, 1967. [Google Scholar] [CrossRef] [PubMed]
- Züst, T.; Agrawal, A.A. Trade-offs between plant growth and defense against insect herbivory: An emerging mechanistic synthesis. Annu. Rev. Plant Biol. 2017, 68, 513–534. [Google Scholar] [CrossRef]
- Michel, W.; Seidling, A.K. Prescher Forest Condition in Europe: 2018 Technical Report of ICP Forests 2018, Report under the UNECE Convention on Long-Range Transboundary Air Pollution (Air Convention) (No. BFW-Dokumentation 25/2018). Available online: https://www.icp-forests.org/pdf/TR2018.pdf (accessed on 25 January 2018).
- Stange, E.E.; Ayres, M.P. Climate Change Impacts: Insects. In Encyclopedia of Life Sciences (ELS); John Wiley & Sons, Ltd.: Chichester, UK, 2010. [Google Scholar] [CrossRef]
- Clark, J.S.; Iverson, L.; Woodall, C.W.; Allen, C.D.; Bell, D.M.; Bragg, D.C.; D’Amato, A.W.; Davis, F.W.; Hersh, M.H.; Ibanez, I.; et al. Impacts of increasing drought on forest dynamics, structure, diversity, and management. In Effects of Drought on Forests and Rangelands in the United States: A Comprehensive Science Synthesis; United States Department of Agriculture: Washington, DC, USA, 2016; pp. 59–96. Available online: https://www.fs.usda.gov/nrs/pubs/jrnl/2016/nrs_2016_clark_002_gtrwo93b.pdf (accessed on 21 February 2016).
- Netherer, S.; Lehmanski, L.; Bachlehner, A.; Rosner, S.; Savi, T.; Schmidt, A.; Huang, J.; Paiva, M.R.; Mateus, E.; Hartmann, H.; et al. Drought increases Norway spruce susceptibility to the Eurasian spruce bark beetle and its associated fungi. New Phytol. 2024, 242, 1000–1017. [Google Scholar] [CrossRef]
- De Bobadilla, M.F.; Vitiello, A.; Erb, E.; Poelman, E.H. Plant defense strategies against attack by multiple herbivores. Trends Plant Sci. 2022, 27, 528–535. [Google Scholar] [CrossRef]
- Naidu, S.; Suman, S.; Roy, A.; Chakraborty, A.; Singh, I.K.; Singh, A. Impact of Genomics Tools and Techniques on Crop Improvement. In Genomics of Plant–Pathogen Interaction and the Stress Response; CRC Press: Boca Raton, FL, USA, 2023; pp. 123–156. [Google Scholar]
- Rathnasamy, S.A.; Gothandapani, S.; Chellamuthu, S.; Chakraborty, A.; Gurusamy, D.; Roy, A. Omics technologies unravelling the plant–pathogen interaction and stress response. In Genomics of Plant–Pathogen Interaction and the Stress Response; CRC Press: Boca Raton, FL, USA, 2023; pp. 74–110. [Google Scholar]
- Biselli, C.; Vietto, L.; Rosso, L.; Cattivelli, L.; Nervo, G.; Fricano, A. Advanced breeding for biotic stress resistance in poplar. Plants 2022, 11, 2032. [Google Scholar] [CrossRef] [PubMed]
- Burdon, J.J.; Zhan, J. Climate change and disease in plant communities. PLoS Biol. 2020, 18, e3000949. [Google Scholar] [CrossRef] [PubMed]
- De Tillesse, V.; Nef, L.; Charles, J.; Hopkin, A.; Augustin, S. Damaging Poplar Insects; FAO: Rome, Italy, 2007; Available online: https://foris.fao.org/static/pdf/ipc/damaging_poplar_insects_eBook.pdf (accessed on 25 April 2024).
- Escobar-Bravo, R.; Klinkhamer, P.G.; Leiss, K.A. Induction of jasmonic acid-associated defenses by thrips alters host suitability for conspecifics and correlates with increased trichome densities in tomato. Plant Cell Physiol. 2017, 58, 622–634. [Google Scholar] [CrossRef]
- Jacob, T.K.; Senthil Kumar, C.M.; Devasahayam, S.; D’Silva, S.; Kumar, R.S.; Biju, C.N.; Praveena, R.; Ankegowda, S.K. Plant morphological traits associated with field resistance to cardamom thrips (Sciothrips cardamomi) in cardamom (Elettaria cardamomum). Ann. Appl. Biol. 2020, 177, 143–151. [Google Scholar] [CrossRef]
- Kaur, B.; Kuraparthy, V.; Bacheler, J.; Fang, H.; Bowman, D.T. Screening germplasm and quantification of components contributing to thrips resistance in cotton. J. Econ. Entomol. 2018, 111, 2426–2434. [Google Scholar] [CrossRef] [PubMed]
- Steenbergen, M.; Abd-el-Haliem, A.; Bleeker, P.; Dicke, M.; Escobar-Bravo, R.; Cheng, G.; Haring, M.A.; Kant, M.R.; Kappers, I.; Klinkhamer, P.G.; et al. Thrips advisor: Exploiting thrips-induced defences to combat pests on crops. J. Exp. Bot. 2018, 69, 1837–1848. [Google Scholar] [CrossRef] [PubMed]
- Bhar, A.; Chakraborty, A.; Roy, A. Plant Responses to Biotic Stress: Old Memories Matter. Plants 2021, 11, 84. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Apuli, R.P.; Bernhardsson, C.; Schiffthaler, B.; Robinson, K.M.; Jansson, S.; Street, N.R.; Ingvarsson, P.K. Inferring the genomic landscape of recombination rate variation in European aspen (Populus tremula). G3 Genes Genomes Genet. 2020, 10, 299–309. [Google Scholar] [CrossRef] [PubMed]
- Legeai, F.; Shigenobu, S.; Gauthier, J.P.; Colbourne, J.; Rispe, C.; Collin, O.; Richards, S.; Wilson, A.C.; Murphy, T.; Tagu, D. AphidBase: A centralized bioinformatic resource for annotation of the pea aphid genome. Insect Mol. Biol. 2010, 19, 5–12. [Google Scholar] [CrossRef]
- Schiffthaler, B.; Delhomme, N.; Bernhardsson, C.; Jenkins, J.; Jansson, S.; Ingvarsson, P.; Schmutz, J.; Street, N. An improved genome assembly of the European aspen Populus tremula. bioRxiv 2019, 805614. [Google Scholar] [CrossRef]
- Sparks, M.E.; Hebert, F.O.; Johnston, J.S.; Hamelin, R.C.; Cusson, M.; Levesque, R.C.; Gundersen-Rindal, D.E. Sequencing, assembly, and annotation of the whole-insect genome of Lymantria dispar dispar, the European gypsy moth. Genes Genomes Genet. 2021, 11, jkab150. [Google Scholar] [CrossRef] [PubMed]
- Wenger, J.A.; Cassone, B.J.; Legeai, F.; Johnston, J.S.; Bansal, R.; Yates, A.D.; Coates, B.S.; Pavinato, V.A.; Michel, A. Whole genome sequence of the soybean aphid, Aphis glycines. Insect Biochem. Mol. Biol. 2020, 123, 102917. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Cong, Q.; Rex, E.A.; Hallwachs, W.; Janzen, D.H.; Grishin, N.V.; Gammon, D.B. Gypsy moth genome provides insights into flight capability and virus-host interactions. Proc. Natl. Acad. Sci. USA 2019, 116, 1669–1678. [Google Scholar] [CrossRef] [PubMed]
- Sharma, H.C.; Ortiz, R. Host plant resistance to insects: An eco-friendly approach for pest management and environment conservation. J. Environ. Biol. 2002, 23, 111–135. Available online: https://hdl.handle.net/10568/107586 (accessed on 25 April 2024). [PubMed]
- Pieterse, C.M.; Van der Does, D.; Zamioudis, C.; Leon-Reyes, A.; Van Wees, S.C. Hormonal modulation of plant immunity. Annu. Rev. Cell Dev. Biol. 2012, 28, 489–521. [Google Scholar] [CrossRef]
- Sarde, S.J.; Bouwmeester, K.; Venegas-Molina, J.; David, A.; Boland, W.; Dicke, M. Involvement of sweet pepper CaLOX2 in jasmonate-dependent induced defence against Western flower thrips. J. Integr. Plant Biol. 2019, 61, 1085–1098. [Google Scholar] [CrossRef]
- Thaler, J.S.; Humphrey, P.T.; Whiteman, N.K. Evolution of jasmonate and salicylate signal crosstalk. Trends Plant Sci. 2012, 17, 260–270. [Google Scholar] [CrossRef]
- Patrick, J.M.; Gary, A. Thompson, Molecular Responses to Aphid Feeding in Arabidopsis in Relation to Plant Defense Pathways. Plant Physiol. 2001, 125, 1074–1085. [Google Scholar] [CrossRef]
- Moran, P.J.; Cheng, Y.; Cassell, J.L.; Thompson, G.A. Gene expression profiling of Arabidopsis thaliana in compatible plant-aphid interactions. Arch. Insect Biochem. Physiol. 2002, 51, 182–203. [Google Scholar] [CrossRef]
- De Vos, M.; Van Oosten, V.R.; Van Poecke, R.M.P.; Van Pelt, J.A.; Pozo, M.J.; Mueller, M.J.; Buchala, A.J.; Metraux, J.P.; Van Loon, L.C.; Dicke, M.; et al. Signal signature and transcriptome changes of Arabidopsis during pathogen and insect attack. Mol. Plant-Microbe Interact. 2005, 18, 923–937. [Google Scholar] [CrossRef]
- Zarate, S.I.; Kempema, L.A.; Walling, L.L. Silverleaf whitefly induces salicylic acid defenses and suppresses effectual jasmonic acid defenses. Plant Physiol. 2007, 143, 866–875. [Google Scholar] [CrossRef] [PubMed]
- Puthoff, D.P.; Holzer, F.M.; Perring, T.M.; Walling, L.L. Tomato Pathogenesis-related Protein Genes are Expressed in Response to Trialeurodes vaporariorum and Bemisia tabaci Biotype B Feeding. J. Chem. Ecol. 2010, 36, 1271–1285. [Google Scholar] [CrossRef] [PubMed]
- Pegadaraju, V.; Knepper, C.; Reese, J.; Shah, J. Premature Leaf Senescence Modulated by the Arabidopsis PHYTOALEXIN DEFICIENT4 Gene Is Associated with Defense against the Phloem-Feeding Green Peach Aphid. Plant Physiol. 2005, 139, 1927–1934. [Google Scholar] [CrossRef] [PubMed]
- Thompson, G.A.; Goggin, F.L. Transcriptomics and functional genomics of plant defence induction by phloem-feeding insects. J. Exp. Bot. 2006, 57, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Feng, Z.; Swihart, R.; Bryant, J.; Huntly, N. Modeling the Impact of Plant Toxicity on Plant–Herbivore Dynamics. J. Dyn. Differ. Equ. 2006, 18, 1021–1042. [Google Scholar] [CrossRef]
- Zhou, G.; Qi, J.; Ren, N.; Cheng, J.; Erb, M.; Mao, B.; Lou, Y. Silencing OsHI-LOX makes rice more susceptible to chewing herbivores, but enhances resistance to a phloem feeder. Plant J. 2009, 60, 638–648. [Google Scholar] [CrossRef]
- Pieterse, C.M.J.; Dicke, M. Plant interactions with microbes and insects: From molecular mechanisms to ecology. Trends Plant Sci. 2007, 12, 564–569. [Google Scholar] [CrossRef]
- Walling, L.L. The myriad plant responses to herbivores. J. Plant Growth Regul. 2000, 19, 195–216. [Google Scholar] [CrossRef] [PubMed]
- Walling, L.L. Avoiding effective defenses: Strategies employed by phloem-feeding insects. Plant Physiol. 2008, 146, 859–866. [Google Scholar] [CrossRef]
- Howe, G.A.; Lightner, J.; Browse, J.; Ryan, C.A. An octadecanoid pathway mutant (JL5) of tomato is compromised in signaling for defense against insect attack. Plant Cell 1996, 8, 2067–2077. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Constabel, C.P.; Yip, L.; Patton, J.J.; Christopher, M.E. Polyphenol oxidase from hybrid poplar. Cloning and expression in response to wounding and herbivory. Plant Physiol. 2000, 124, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Green, T.R.; Ryan, C.A. Wound-induced proteinase inhibitor in plant leaves: A possible defense mechanism against insects. Science 1972, 175, 776–777. [Google Scholar] [CrossRef] [PubMed]
- Glawe, G.A.; Zavala, A.; Kessler, A.; Van Dam, N.M.; Baldwin, I.T. Ecological costs and benefits correlated with trypsin protease inhibitor production in Nicotiana attenuata. Ecology 2003, 84, 79–90. [Google Scholar] [CrossRef]
- Thaler, J.S.; Farag, M.A.; Pare, P.W.; Dicke, M. Jasmonate-deficient plants have reduced direct and indirect defences against herbivores. Ecol. Lett. 2002, 5, 764–774. [Google Scholar] [CrossRef]
- Avdiushko, S.A.; Brown, G.C.; Dahlman, D.L.; Hildebrand, D.F. Methyl jasmonate exposure induces insect resistance in cabbage and tobacco. Environ. Entomol. 1997, 26, 642–654. [Google Scholar] [CrossRef]
- Stout, M.J.; Workman, K.V.; Bostock, R.M.; Duffey, S.S. Stimulation and attenuation of induced resistance by elicitors and inhibitors of chemical induction in tomato (Lycopersicon esculentum) foliage. Entomol. Exp. Et Appl. 1998, 86, 267–279. [Google Scholar] [CrossRef]
- Cipollini, D.F.; Redman, A.M. Age-dependent effects of jasmonic acid treatment and wind exposure on foliar oxidase activity and insect resistance in tomato. J. Chem. Ecol. 1999, 25, 271–281. [Google Scholar] [CrossRef]
- Omer, A.D.; Granett, J.; Karban, R.; Villa, E.M. Chemically induced resistance against multiple pests in cotton. Int. J. Pest Manag. 2001, 47, 49–54. [Google Scholar] [CrossRef]
- Boughton, A.J.; Hoover, K.; Felton, G.W. Methyl jasmonate application induces increased densities of glandular trichomes on tomato, Lycopersicon esculentum. J. Chem. Ecol. 2005, 31, 2211–2216. [Google Scholar] [CrossRef]
- De Vos, M.; Van Zaanen, W.; Koorneef, A.; Korzelius, J.P.; Dicke, M.; Van Loon, L.C.; Pieterse, C.M.J. Herbivore-induced resistance against microbial pathogens in Arabidopsis. Plant Physiol. 2006, 142, 352–363. [Google Scholar] [CrossRef]
- Diezel, C.; von Dahl, C.; Gaquerel, E.; Baldwin, I.T. Different lepidopteran elicitors account for crosstalk in herbivory-induced phytohormone signaling. Plant Physiol. 2009, 150, 1576–1586. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Baldwin, I.T. Herbivory-induced signalling in plants: Perception and action. Plant Cell Environ. 2009, 32, 1161–1174. [Google Scholar] [CrossRef] [PubMed]
- Ellis, C.; Karafyllidis, I.; Turner, J.G. Constitutive activation of jasmonate signaling in an Arabidopsis mutant correlates with enhanced resistance to Erysiphe cichoracearum, Pseudomonas syringae, and Myzus persicae. Mol. Plant-Microbe Interact. 2002, 15, 1025–1030. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Allmann, S.; Wu, J.S.; Baldwin, I.T. Comparisons of LIPOXYGENASE3- and JASMONATE-RESISTANT4/6-silenced plants reveal that jasmonic acid and jasmonic acid-amino acid conjugates play different roles in herbivore resistance of Nicotiana attenuata. Plant Physiol. 2008, 146, 904–915. [Google Scholar] [CrossRef] [PubMed]
- Spoel, S.H.; Koornneef, A.; Claessens, S.M.C.; Korzelius, J.P.; Van Pelt, J.A.; Mueller, M.J.; Buchala, A.J.; Me’traux, J.P.; Brown, R.; Kazan, K.; et al. NPR1 modulates cross-talk between salicylate- and jasmonate-dependent defense pathways through a novel function in the cytosol. Plant Cell 2003, 15, 760–770. [Google Scholar] [CrossRef] [PubMed]
- Glazebrook, J. Contrasting mechanisms of defense against biotrophic and necrotrophic pathogens. Annu. Rev. Phytopathol. 2005, 43, 205–227. [Google Scholar] [CrossRef] [PubMed]
- Mur, L.A.J.; Kenton, P.; Atzorn, R.; Miersch, O.; Wasternack, C. The outcomes of concentration-specific interactions between salicylate and jasmonate signaling include synergy, antagonism, and oxidative stress leading to cell death. Plant Physiol. 2006, 140, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Kawazu, K.; Mochizuki, A.; Sato, Y.; Sugeno, W.; Murata, M.; Seo, S.; Mitsuhara, I. Different expression profiles of jasmonic acid and salicylic acid inducible genes in the tomato plant against herbivores with various feeding modes. Arthropod-Plant Interact. 2012, 6, 221–230. [Google Scholar] [CrossRef]
- Tetreault, H.M.; Grover, S.; Scully, E.D.; Gries, T.; Palmer, N.A.; Sarath, G.; Louis, J.; Sattler, S.E. Global responses of resistant and susceptible sorghum (Sorghum bicolor) to sugarcane aphid (Melanaphis sacchari). Front. Plant Sci. 2019, 10, 426693. [Google Scholar] [CrossRef]
- Shalileh, S.; Ogada, P.A.; Moualeu, D.P.; Poehling, H.M. Manipulation of Frankliniella occidentalis (Thysanoptera: Thripidae) by Tomato Spotted Wilt Virus (Tospovirus) via the host plant nutrients to enhance its transmission and spread. Environ. Entomol. 2016, 45, 1235–1242. [Google Scholar] [CrossRef]
- Hettenhausen, C.; Schuman, M.C.; Wu, J. MAPK signaling: A key element in plant defense response to insects. Insect Sci. 2015, 22, 157–164. [Google Scholar] [CrossRef] [PubMed]
- Manjeet, K.; Yadav, S. Role of Mapks During Plant-Insect Interaction. In Plant-Pest Interactions: From Molecular Mechanisms to Chemical Ecology: Chemical Ecology; Springer: Berlin/Heidelberg, Germany, 2021; pp. 93–106. [Google Scholar] [CrossRef]
- Yadav, M.; Pandey, J.; Chakraborty, A.; Hassan, M.I.; Kundu, J.K.; Roy, A.; Singh, I.K.; Singh, A. A Comprehensive Analysis of Calmodulin-Like Proteins of Glycine max Indicates Their Role in Calcium Signaling and Plant Defense Against Insect Attack. Front. Plant Sci. 2022, 13, 817950. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Abidallha, E.H.; Li, Y.; Li, H.; Chen, Y.; Tambel, L.I.; Hu, D.; Chen, Y.; Zhang, X.; Chen, D. Amino acid composition and level affect Bt protein concentration in Bt cotton. Plant Growth Regul. 2017, 82, 439–446. [Google Scholar] [CrossRef]
- Zhou, S.; Lou, Y.R.; Tzin, V.; Jander, G. Alteration of plant primary metabolism in response to insect herbivory. Plant Physiol. 2015, 169, 1488–1498. [Google Scholar] [CrossRef] [PubMed]
- Abebe, W. Review on plant defense mechanisms against insect pests. Int. J. Nov. Res. Interdiscip. Stud. 2021, 8, 15–39. Available online: https://www.noveltyjournals.com/upload/paper/REVIEW%20ON%20PLANT%20DEFENSE%20MECHANISMS.pdf (accessed on 25 April 2024).
- Morin, S.; Atkinson, P.W.; Walling, L.L. Whitefly–Plant Interactions: An Integrated Molecular Perspective. Annu. Rev. Entomol. 2023, 69, 503–525. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Mahecha, O.; Klink, S.; Heinen, R.; Rothballer, M.; Zytynska, S. Impaired microbial N-acyl homoserine lactone signalling increases plant resistance to aphids across variable abiotic and biotic environments. Plant Cell Environ. 2022, 45, 3052–3069. [Google Scholar] [CrossRef]
- Appel, H.M.; Fescemyer, H.; Ehlting, J.; Weston, D.; Rehrig, E.; Joshi, T.; Xu, D.; Bohlmann, J.; Schultz, J. Transcriptional responses of Arabidopsis thaliana to chewing and sucking insect herbivores. Front. Plant Sci. 2014, 5, 110818. [Google Scholar] [CrossRef]
- Steinbrenner, A.D.; Gómez, S.; Osorio, S.; Fernie, A.R.; Orians, C.M. Herbivore-induced changes in tomato (Solanum lycopersicum) primary metabolism: A whole plant perspective. J. Chem. Ecol. 2011, 37, 1294–1303. [Google Scholar] [CrossRef]
- Sandström, J.; Telang, A.; Moran, N.A. Nutritional enhancement of host plants by aphids—A comparison of three aphid species on grasses. J. Insect Physiol. 2000, 46, 33–40. [Google Scholar] [CrossRef]
- Koyama, Y.; Yao, I.; Akimoto, S.I. Aphid galls accumulate high concentrations of amino acids: A support for the nutrition hypothesis for gall formation. Entomol. Exp. Et Appl. 2004, 113, 35–44. [Google Scholar] [CrossRef]
- Marti, G.; Erb, M.; Boccard, J.; Glauser, G.; Doyen, G.R.; Villard, N.; Robert, C.A.M.; Turlings, T.C.; Rudaz, S.; Wolfender, J.L. Metabolomics reveals herbivore-induced metabolites of resistance and susceptibility in maize leaves and roots. Plant Cell Environ. 2013, 36, 621–639. [Google Scholar] [CrossRef] [PubMed]
- Karmakar, A.; Malik, U.; Barik, A. Effects of leaf epicuticular wax compounds from Solena amplexicaulis (Lam.) Gandhi on olfactory responses of a generalist insect herbivore. Allelopath. J. 2016, 37, 253–272. [Google Scholar]
- Reina-Pinto, J.J.; Yephremov, A. Surface lipids and plant defenses. Plant Physiol. Biochem. 2009, 47, 540–549. [Google Scholar] [CrossRef] [PubMed]
- Zogli, P.; Pingault, L.; Grover, S.; Louis, J. Ento(o)mics: The intersection of ‘omic’approaches to decipher plant defense against sap-sucking insect pests. Curr. Opin. Plant Biol. 2020, 56, 153–161. [Google Scholar] [CrossRef]
- Mehta, S.; Chakraborty, A.; Roy, A.; Singh, I.K.; Singh, A. Fight Hard or Die Trying: Current Status of Lipid Signaling during Plant-Pathogen Interaction. Plants 2021, 10, 1098. [Google Scholar] [CrossRef] [PubMed] [PubMed Central]
- Botha, A.M.; Lacock, L.; van Niekerk, C.; Matsioloko, M.T.; du Preez, F.B.; Loots, S.; Venter, E.; Kunert, K.J.; Cullis, C.A. Is photosynthetic transcriptional regulation in Triticum aestivum L. cv.‘TugelaDN’a contributing factor for tolerance to Diuraphis noxia (Homoptera: Aphididae)? Plant Cell Rep. 2006, 25, 41–54. [Google Scholar] [CrossRef]
- Gutsche, A.; Heng-Moss, T.; Sarath, G.; Twigg, P.A.U.L.; Xia, Y.; Lu, G.; Mornhinweg, D. Gene expression profiling of tolerant barley in response to Diuraphis noxia (Hemiptera: Aphididae) feeding. Bull. Entomol. Res. 2009, 99, 163–173. [Google Scholar] [CrossRef]
- Coppola, V.; Coppola, M.; Rocco, M.; Digilio, M.C.; D’Ambrosio, C.; Renzone, G.; Martinelli, R.; Scaloni, A.; Pennacchio, F.; Rao, R.; et al. Transcriptomic and proteomic analysis of a compatible tomato-aphid interaction reveals a predominant salicylic acid-dependent plant response. BMC Genom. 2013, 14, 515. [Google Scholar] [CrossRef]
- Lawrence, S.D.; Novak, N.G.; Ju, C.J.T.; Cooke, J.E. Potato, Solanum tuberosum, defense against Colorado potato beetle, Leptinotarsa decemlineata (Say): Microarray gene expression profiling of potato by Colorado potato beetle regurgitant treatment of wounded leaves. J. Chem. Ecol. 2008, 34, 1013–1025. [Google Scholar] [CrossRef]
- Arimura, G.I.; Ozawa, R.; Shimoda, T.; Nishioka, T.; Boland, W.; Takabayashi, J. Herbivory-induced volatiles elicit defence genes in lima bean leaves. Nature 2000, 406, 512–515. [Google Scholar] [CrossRef]
- Velikova, V.; Salerno, G.; Frati, F.; Peri, E.; Conti, E.; Colazza, S.; Loreto, F. Influence of feeding and oviposition by phytophagous pentatomids on photosynthesis of herbaceous plants. J. Chem. Ecol. 2010, 36, 629–641. [Google Scholar] [CrossRef]
- Castrillón-Arbeláez, P.A.; Martínez-Gallardo, N.; Arnaut, H.A.; Tiessen, A.; Délano-Frier, J.P. Metabolic and enzymatic changes associated with carbon mobilization, utilization and replenishment triggered in grain amaranth (Amaranthus cruentus) in response to partial defoliation by mechanical injury or insect herbivory. BMC Plant Biol. 2012, 12, 163. [Google Scholar] [CrossRef]
- Priyanka, S.L.; Navya, R.N.; Chandrasekaran, M.; Hasan, W. Role of Secondary Plant Metabolites against Insects; Biotech Books: Mumbai, India, 2022; pp. 325–340. [Google Scholar]
- Grayson, D.H. Monoterpenoids. Nat. Prod. Rep. 1998, 5, 497–521. [Google Scholar] [CrossRef]
- López-Goldar, X.; Villari, C.; Bonello, P.; Borg-Karlson, A.K.; Grivet, D.; Zas, R.; Sampedro, L. Inducibility of plant secondary metabolites in the stem predicts genetic variation in resistance against a key insect herbivore in maritime pine. Front. Plant Sci. 2018, 9, 1651. [Google Scholar] [CrossRef]
- Ren, X.; Wu, S.; Xing, Z.; Gao, Y.; Cai, W.; Lei, Z. Abundances of thrips on plants in vegetative and flowering stages are related to plant volatiles. J. Appl. Entomol. 2020, 144, 732–742. [Google Scholar] [CrossRef]
- Soltis, N.E.; Kliebenstein, D.J. Natural variation of plant metabolism: Genetic mechanisms, interpretive caveats, and evolutionary and mechanistic insights. Plant Physiol. 2015, 169, 1456–1468. [Google Scholar] [CrossRef]
- Wink, M. Plant secondary metabolites modulate insect behavior-steps toward addiction? Front. Physiol. 2018, 9, 364. [Google Scholar] [CrossRef]
- Falcone Ferreyra, M.L.; Rius, S.P.; Casati, P. Flavonoids: Biosynthesis, biological functions, and biotechnological applications. Front. Plant Sci. 2012, 3, 222. [Google Scholar] [CrossRef]
- Li, L.; Li, T.; Jiang, Y.; Yang, Y.; Zhang, L.; Jiang, Z.; Wei, C.; Wan, X.; Yang, H. Alteration of local and systemic amino acids metabolism for the inducible defense in tea plant (Camellia sinensis) in response to leaf herbivory by Ectropis oblique. Arch. Biochem. Biophys. 2020, 683, 108301. [Google Scholar] [CrossRef]
- Jin, S.; Ren, Q.; Lian, L.; Cai, X.; Bian, L.; Luo, Z.; Li, Z.; Ye, N.; Wei, R.; He, W.; et al. Comparative transcriptomic analysis of resistant and susceptible tea cultivars in response to Empoasca onukii (Matsuda) damage. Planta 2020, 252, 10. [Google Scholar] [CrossRef]
- Jing, T.; Du, W.; Qian, X.; Wang, K.; Luo, L.; Zhang, X.; Deng, Y.; Li, B.; Gao, T.; Zhang, M.; et al. UGT89AC1-mediated quercetin glucosylation is induced upon herbivore damage and enhances Camellia sinensis resistance to insect feeding. Plant Cell Environ. 2024, 47, 682–697. [Google Scholar] [CrossRef]
- Zhao, X.; Chen, S.; Wang, S.; Shan, W.; Wang, X.; Lin, Y.; Su, F.; Yang, Z.; Yu, X. Defensive responses of tea plants (Camellia sinensis) against tea green leafhopper attack: A multi-omics study. Front. Plant Sci. 2020, 10, 1705. [Google Scholar] [CrossRef]
- Sun, Y.; Xia, X.L.; Jiang, J.F.; Chen, S.M.; Chen, F.D.; Lv, G.S. Salicylic acid-induced changes in physiological parameters and genes of the flavonoid biosynthesis pathway in Artemisia vulgaris and Dendranthema nankingense during aphid feeding. Genet. Mol. Res. 2016, 15, gmr.15017546. [Google Scholar] [CrossRef]
- War, A.R.; Buhroo, A.A.; Hussain, B.; Ahmad, T.; Nair, R.M.; Sharma, H.C. Plant defense and insect adaptation with reference to secondary metabolites. In Co-Evolution of Secondary Metabolites; Springer: Berlin/Heidelberg, Germany, 2020; pp. 795–822. [Google Scholar] [CrossRef]
- Wari, D.; Aboshi, T.; Shinya, T.; Galis, I. Integrated view of plant metabolic defense with particular focus on chewing herbivores. J. Integr. Plant Biol. 2022, 64, 449–475. [Google Scholar] [CrossRef]
- Chen, H.; Su, H.; Zhang, S.; Jing, T.; Liu, Z.; Yang, Y. Transcriptomic and Metabolomic Responses in Cotton Plant to Apolygus lucorum Infestation. Insects 2022, 13, 391. [Google Scholar] [CrossRef]
- Dixit, G.; Srivastava, A.; Rai, K.M.; Dubey, R.S.; Srivastava, R.; Verma, P.C. Distinct defensive activity of phenolics and phenylpropanoid pathway genes in different cotton varieties toward chewing pests. Plant Signal. Behav. 2020, 15, 1747689. [Google Scholar] [CrossRef] [PubMed]
- Boruah, J. Effect of 0.1% HgCl2 on Surface Sterilization of Som (Persea bombycina King) Explant during Tissue Culture—A Major Host Plant of Muga Silkworm. Int. J. Curr. Microbiol. App. Sci. 2020, 9, 954–958. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A Revised Medium for Rapid Growth and Bio Assays with Tobacco Tissue Cultures. Physiol. Plant. 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Hajek, A. Larval behavior in Lymantria dispar increases risk of fungal infection. Oecologia 2001, 126, 285–291. [Google Scholar] [CrossRef]
- Mewis, I.; Appel, H.M.; Hom, A.; Raina, R.; Schultz, J.C. Major signaling pathways modulate Arabidopsis glucosinolate accumulation and response to both phloem-feeding and chewing insects. Plant Physiol. 2005, 138, 1149–1162. [Google Scholar] [CrossRef] [PubMed]
- Mewis, I.; Tokuhisa, J.G.; Schultz, J.C.; Appel, H.M.; Ulrichs, C.; Gershenzon, J. Gene expression and glucosinolate accumulation in Arabidopsis thaliana in response to generalist and specialist herbivores of different feeding guilds and the role of defense signaling pathways. Phytochemistry 2006, 67, 2450–2462. [Google Scholar] [CrossRef] [PubMed]
- Sellamuthu, G.; Naseer, A.; Hradecký, J.; Chakraborty, A.; Synek, J.; Modlinger, R.; Roy, A. Gene expression plasticity facilitates different host feeding in Ips sexdentatus (Coleoptera: Curculionidae: Scolytinae). Insect Biochem. Mol. Biol. 2024, 165, 104061. [Google Scholar] [CrossRef] [PubMed]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef]
- Eisen, M.B.; Spellman, P.T.; Brown, P.O.; Botstein, D. Cluster analysis and display of genome-wide expression patterns. Proc. Natl. Acad. Sci. USA 1998, 95, 14863–14868. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Sen, M.K.; Hamouzová, K.; Košnarová, P.; Roy, A.; Soukup, J. Identification of the most suitable reference gene for gene expression studies with development and abiotic stress response in Bromus sterilis. Sci. Rep. 2021, 11, 13393. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control Leaves vs. Moth-Infested Leaves | Control Leaves vs. Aphid-Infested Leaves | Aphid-Infested Leaves vs. Moth-Infested Leaves | ||||||
|---|---|---|---|---|---|---|---|---|
| Metabolite | VIP Value | KO Number | Metabolite | VIP Value | KO Number | Metabolite | VIP Value | KO Number |
| Myricetin 3-2 | 1.7 | 00941 | Trihydroxypentamethoxyflavone | 2.2 | 00941 | Catechin | 1.8 | 00941 |
| Trihydroxypentamethoxyflavone | 1.6 | 00941 | Benzoic acid | 2.2 | 00362 | 2-Caffeoylisocitrate | 1.7 | 00940 |
| Diethyl succinate | 1.6 | 00020 | Cinnamoyl galloylglucose | 2.1 | 00941 | Proanthocyanidin 1 | 1.6 | 00942 |
| Cinnamoyl galloylglucose | 1.6 | 00941 | Quercetin-O-glucuronide derivate | 2.1 | 00941 | Fructose phosphate | 1.6 | 00010 |
| Flavonoid 5 | 1.5 | 00941 | Catechin-O-rhmnoside | 1.8 | 00941 | Heptamethoxyflavone | 1.5 | 00941 |
| Salicin | 1.4 | 00940 | Syringin | 1.7 | 00940 | Proanthocyanidin 3 | 1.5 | 00942 |
| Quercetin 3,5-digalactoside | 1.4 | 00941 | Veronicoside | 1.6 | 00941 | Benzoic acid | 1.4 | 00362 |
| Fructose phosphate | 1.4 | 00010 | Butyl O-caffeoylquinate | 1.5 | 00940 | Methyl salicylate | 1.3 | 00940 |
| Di-O-caffeoylquinic acid | 1.4 | 00940 | O-Vanilloylvitexin | 1.5 | 00940 | Apigenin | 1.3 | 00941 |
| Quercetin-O-glucuronide derivate | 1.4 | 00941 | Isorientin 3′,6′di-O-glucoside | 1.5 | 00941 | O-Caffeoyl-O-methylquinic acid | 1.3 | 00940 |
| Gene Name | geNorm | NormFinder | BestKeeper | ΔCT | Comprehensive | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| M | R | SV | R | SD | R | SD | R | GM | R | |
| Ubiquitin | 1.32 | 3 | 1.334 | 5 | 2.95 | 6 | 2.14 | 4 | 4.68 | 5 |
| GAPDH | 0.841 | 1 | 1.002 | 3 | 2 | 2 | 2.08 | 3 | 2.06 | 2 |
| Act7 | 1.078 | 2 | 0.785 | 2 | 2.48 | 5 | 1.9 | 2 | 2.78 | 3 |
| PP2A | 0.841 | 1 | 0.42 | 1 | 1.95 | 1 | 1.82 | 1 | 1 | 1 |
| EF1B1 | 1.662 | 5 | 2.079 | 6 | 3.31 | 7 | 2.57 | 6 | 6.24 | 7 |
| EF1A | 2.468 | 6 | 4.331 | 7 | 2.44 | 4 | 4.48 | 7 | 6.09 | 6 |
| Tub4 | 1.47 | 4 | 1.29 | 4 | 2.2 | 3 | 2.28 | 5 | 4.16 | 4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pastierovič, F.; Mogilicherla, K.; Hradecký, J.; Kalyniukova, A.; Dvořák, O.; Roy, A.; Tomášková, I. Genome-Wide Transcriptomic and Metabolomic Analyses Unveiling the Defence Mechanisms of Populus tremula against Sucking and Chewing Insect Herbivores. Int. J. Mol. Sci. 2024, 25, 6124. https://doi.org/10.3390/ijms25116124
Pastierovič F, Mogilicherla K, Hradecký J, Kalyniukova A, Dvořák O, Roy A, Tomášková I. Genome-Wide Transcriptomic and Metabolomic Analyses Unveiling the Defence Mechanisms of Populus tremula against Sucking and Chewing Insect Herbivores. International Journal of Molecular Sciences. 2024; 25(11):6124. https://doi.org/10.3390/ijms25116124
Chicago/Turabian StylePastierovič, Filip, Kanakachari Mogilicherla, Jaromír Hradecký, Alina Kalyniukova, Ondřej Dvořák, Amit Roy, and Ivana Tomášková. 2024. "Genome-Wide Transcriptomic and Metabolomic Analyses Unveiling the Defence Mechanisms of Populus tremula against Sucking and Chewing Insect Herbivores" International Journal of Molecular Sciences 25, no. 11: 6124. https://doi.org/10.3390/ijms25116124