
Rapid Determination of SARS-CoV-2 Integrity and Infectivity by Using Propidium Monoazide Coupled with Digital Droplet PCR

 , , , and
, , , and {kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
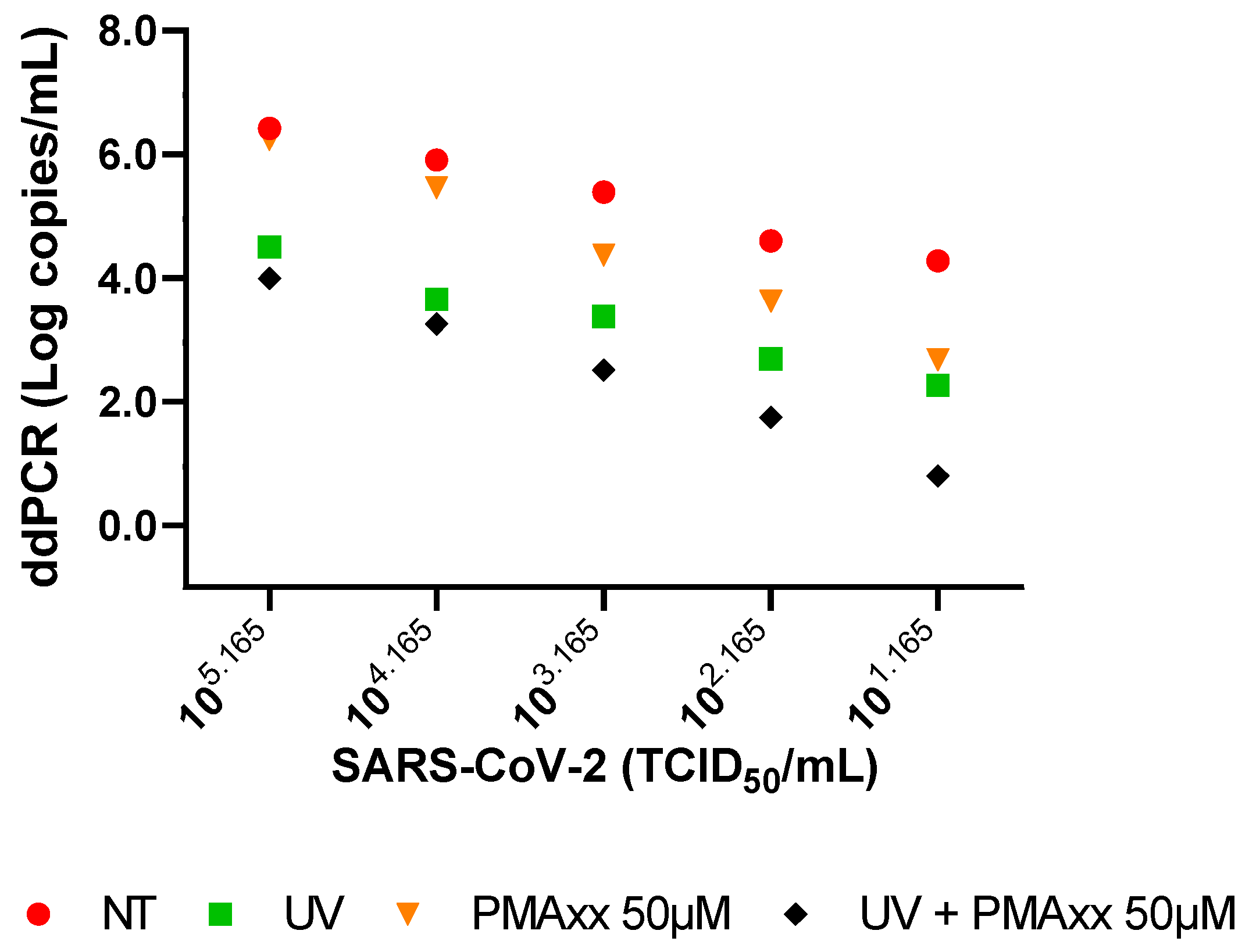
2. Results
2.1. In Vitro Settings of PMAxx Treatment
2.2. Clinical Samples Treated with PMAxx
2.3. Comparison of PMAxx-ddPCR with SARS-CoV-2 Isolation and Negative-Chain PCR
3. Discussion
4. Materials and Methods
4.1. SARS-CoV-2 Stock Preparation and UV Inactivation
4.2. Clinical Specimens
4.3. Propidium Monoazide Treatment and Viral RNA Extraction
4.4. SARS-CoV-2 Quantification with ddPCR
4.5. Negative-Chain SARS-CoV-2 RNA PCR
4.6. SARS-CoV-2 Isolation from Clinical Samples
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- World Health Organization. Available online: https://www.who.int/emergencies/diseases/novel-coronavirus-2019 (accessed on 3 May 2024).
- Chan, M.; Linn, M.M.N.; O’Hagan, T.; Guerra-Assunção, J.A.; Lackenby, A.; Workman, S.; Dacre, A.; Burns, S.O.; Breuer, J.; Hart, J.; et al. Persistent SARS-CoV-2 PCR Positivity Despite Anti-Viral Treatment in Immunodeficient Patients. J. Clin. Immunol. 2023, 43, 1083–1092. [Google Scholar] [CrossRef]
- Grimaldi, P.; Russo, A.; Pisaturo, M.; Maggi, P.; Allegorico, E.; Gentile, I.; Sangiovanni, V.; Rossomando, A.; Pacilio, R.; Calabria, G.; et al. Clinical and epidemiological factors causing longer SARS-CoV-2 viral shedding: The results from the CoviCamp cohort. Infection 2024, 52, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Danzetta, M.L.; Amato, L.; Cito, F.; Di Giuseppe, A.; Morelli, D.; Savini, G.; Mercante, M.T.; Lorusso, A.; Portanti, O.; Puglia, I.; et al. SARS-CoV-2 RNA Persistence in Naso-Pharyngeal Swabs. Microorganisms 2020, 8, 1124. [Google Scholar] [CrossRef]
- De Carvalho, J.G.; Hvozdara, K. What are the Clinical Implications of a Positive RT-PCR Test 6 Months after a Mild SARS-CoV-2 Infection? Eur. J. Case Rep. Intern. Med. 2021, 8, 002463. [Google Scholar] [CrossRef]
- Laferl, H.; Kelani, H.; Seitz, T.; Holzer, B.; Zimpernik, I.; Steinrigl, A.; Schmoll, F.; Wenisch, C.; Allerberger, F. An approach to lifting self-isolation for health care workers with prolonged shedding of SARS-CoV-2 RNA. Infection 2021, 49, 95–101. [Google Scholar] [CrossRef]
- National Institutes of Health. Available online: https://www.covid19treatmentguidelines.nih.gov/special-populations/immunocompromised/ (accessed on 7 May 2024).
- Runfeng, L.; Yunlong, H.; Jicheng, H.; Weiqi, P.; Qinhai, M.; Yongxia, S.; Chufang, L.; Jin, Z.; Zhenhua, J.; Haiming, J.; et al. Lianhuaqingwen exerts anti-viral and anti-inflammatory activity against novel coronavirus (SARS-CoV-2). Pharmacol. Res. 2020, 156, 104761. [Google Scholar] [CrossRef]
- Manenti, A.; Maggetti, M.; Casa, E.; Martinuzzi, D.; Torelli, A.; Trombetta, C.M.; Marchi, S.; Montomoli, E. Evaluation of SARS-CoV-2 neutralizing antibodies using a CPE-based colorimetric live virus micro-neutralization assay in human serum samples. J. Med. Virol. 2020, 92, 2096–2104. [Google Scholar] [CrossRef]
- Colavita, F.; Lapa, D.; Carletti, F.; Lalle, E.; Messina, F.; Rueca, M.; Matusali, G.; Meschi, S.; Bordi, L.; Marsella, P.; et al. Virological Characterization of the First 2 COVID-19 Patients Diagnosed in Italy: Phylogenetic Analysis, Virus Shedding Profile from Different Body Sites, and Antibody Response Kinetics. Open Forum Infect. Dis. 2020, 7, ofaa403. [Google Scholar] [CrossRef] [PubMed]
- Iannetta, M.; Lalle, E.; Musso, M.; Carletti, F.; Scorzolini, L.; D’Abramo, A.; Chinello, P.; Castilletti, C.; Ippolito, G.; Capobianchi, M.R.; et al. Persistent detection of dengue virus RNA in vaginal secretion of a woman returning from Sri Lanka to Italy, April 2017. Euro Surveill 2017, 22, 30600. [Google Scholar] [CrossRef]
- Biava, M.; Caglioti, C.; Castilletti, C.; Bordi, L.; Carletti, F.; Colavita, F.; Quartu, S.; Nicastri, E.; Iannetta, M.; Vairo, F.; et al. Persistence of ZIKV-RNA in the cellular fraction of semen is accompanied by a surrogate-marker of viral replication. Diagnostic implications for sexual transmission. New Microbiol. 2018, 41, 30–33. [Google Scholar]
- Biava, M.; Caglioti, C.; Bordi, L.; Castilletti, C.; Colavita, F.; Quartu, S.; Nicastri, E.; Lauria, F.N.; Petrosillo, N.; Lanini, S.; et al. Detection of Viral RNA in Tissues following Plasma Clearance from an Ebola Virus Infected Patient. PLoS Pathog. 2017, 13, e10006065. [Google Scholar] [CrossRef] [PubMed]
- Vita, S.; Bordi, L.; Sberna, G.; Caputi, P.; Lapa, D.; Corpolongo, A.; Mija, C.; D’Abramo, A.; Maggi, F.; Vairo, F.; et al. Autochthonous Dengue Fever in 2 Patients, Rome, Italy. Emerg. Infect. Dis. 2024, 30, 183–184. [Google Scholar] [CrossRef] [PubMed]
- Sberna, G.; Biagi, M.; Marafini, G.; Nardacci, R.; Biava, M.; Colavita, F.; Piselli, P.; Miraldi, E.; D’Offizi, G.; Capobianchi, M.R.; et al. In vitro Evaluation of Antiviral Efficacy of a Standardized Hydroalcoholic Extract of Poplar Type Propolis Against SARS-CoV-2. Front. Microbiol. 2022, 13, 799546. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Zhou, X.; He, X.; Wang, P.; Yue, S.; Wu, L.; Zhang, Y.; Xie, Q.; Zhang, B.; Zhao, W. Detection of infectious dengue virus by selective real-time quantitative polymerase chain reaction. Virol. Sin. 2016, 31, 342–345. [Google Scholar] [CrossRef]
- Leifels, M.; Cheng, D.; Sozzi, E.; Shoults, D.C.; Wuertz, S.; Mongkolsuk, S.; Sirikanchana, K. Capsid integrity quantitative PCR to determine virus infectivity in environmental and food applications—A systematic review. Water Res. X 2020, 11, 100080. [Google Scholar] [CrossRef] [PubMed]
- Hong, W.; Xiong, J.; Nyaruaba, R.; Li, J.; Muturi, E.; Liu, H.; Yu, J.; Yang, H.; Wei, H. Rapid determination of infectious SARS-CoV-2 in PCR-positive samples by SDS-PMA assisted RT-qPCR. Sci. Total Environ. 2021, 797, 149085. [Google Scholar] [CrossRef]
- Nocker, A.; Cheung, C.Y.; Camper, A.K. Comparison of propidium monoazide with ethidium monoazide for differentiation of live vs. dead bacteria by selective removal of DNA from dead cells. J. Microbiol. Methods 2006, 67, 310–320. [Google Scholar] [CrossRef]
- Karim, M.R.; Fout, G.S.; Johnson, C.H.; White, K.M.; Parshionikar, S.U. Propidium monoazide reverse transcriptase PCR and RT-qPCR for detecting infectious enterovirus and norovirus. J. Virol. Methods. 2015, 219, 51–61. [Google Scholar] [CrossRef]
- Li, Y.; Wang, Z.; Qing, J.; Hu, D.; Vo, H.T.; Thi, K.T.; Wang, X.; Li, X. Application of propidium monoazide quantitative PCR to discriminate of infectious African swine fever viruses. Front. Microbiol. 2024, 14, 1290302. [Google Scholar] [CrossRef]
- Kim, K.H.; Kang, G.; Woo, W.S.; Sohn, M.Y.; Son, H.J.; Park, C.I. Development of a Propidium Monoazide-Based Viability Quantitative PCR Assay for Red Sea Bream Iridovirus Detection. Int. J. Mol. Sci. 2023, 24, 3426. [Google Scholar] [CrossRef]
- Zeng, D.; Qian, B.; Li, Y.; Zong, K.; Peng, W.; Liao, K.; Yu, X.; Sun, J.; Lv, X.; Ding, L.; et al. Prospects for the application of infectious virus detection technology based on propidium monoazide in African swine fever management. Front. Microbiol. 2022, 13, 1025758. [Google Scholar] [CrossRef] [PubMed]
- Patrone, P.N.; Romsos, E.L.; Cleveland, M.H.; Vallone, P.M.; Kearsley, A.J. Affine analysis for quantitative PCR measurements. Anal. Bioanal. Chem. 2020, 412, 7977–7988. [Google Scholar] [CrossRef] [PubMed]
- DeJaco, R.F.; Roberts, M.J.; Romsos, E.L.; Vallone, P.M.; Kearsley, A.J. Reducing Bias and Quantifying Uncertainty in Fluorescence Produced by PCR. Bull. Math. Biol. 2023, 85, 83. [Google Scholar] [CrossRef]
- Yao, H.; Song, Y.; Chen, Y.; Wu, N.; Xu, J.; Sun, C.; Zhang, J.; Weng, T.; Zhang, Z.; Wu, Z.; et al. Molecular Architecture of the SARS-CoV-2 Virus. Cell 2020, 183, 730–738. [Google Scholar] [CrossRef] [PubMed]
- La Scola, B.; Le Bideau, M.; Andreani, J.; Hoang, V.T.; Grimaldier, C.; Colson, P.; Gautret, P.; Raoult, D. Viral RNA load as determined by cell culture as a management tool for discharge of SARS-CoV-2 patients from infectious disease wards. Eur. J. Clin. Microbiol. Infect. Dis. 2020, 39, 1059–1061. [Google Scholar] [CrossRef] [PubMed]
- Beeching, N.J.; Fletcher, T.E.; Beadsworth, M.B.J. Covid-19: Testing times. BMJ 2020, 369, 1403. [Google Scholar] [CrossRef] [PubMed]
- Kaul, D. An overview of coronaviruses including the SARS-2 coronavirus—Molecular biology, epidemiology and clinical implications. Curr. Med. Res. Pract. 2020, 10, 54–64. [Google Scholar] [CrossRef]
- Liu, D.; Shan, Y.; Zhu, X.; An, B.; Luo, J.; Yang, N.; Zhao, K.; Wu, T.; Qiao, Q.; Zhu, F.; et al. Establishment and Evaluation of Propidium Monoazide-Digital Pcr (Pma-Dpcr) for Rapid Discrimination of Infectious Sars-Cov-2. SSRN, 2023; in preprint. [Google Scholar] [CrossRef]
- Roingeard, P.; Eymieux, S.; Burlaud-Gaillard, J.; Hourioux, C.; Patient, R.; Blanchard, E. The double-membrane vesicle (DMV): A virus-induced organelle dedicated to the replication of SARS-CoV-2 and other positive-sense single-stranded RNA viruses. Cell Mol. Life Sci. 2022, 79, 425. [Google Scholar] [CrossRef] [PubMed]
- Gisaid. Available online: https://gisaid.org/ (accessed on 3 May 2024).
- European Virus Archive—GLOBAL. Available online: https://www.european-virus-archive.com/evag-news/sars-cov-2-collection (accessed on 3 May 2024).
- Reed, L.J.; Muench, H. A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 1938, 27, 493–497. [Google Scholar] [CrossRef]
- Biotium. Available online: https://biotium.com/ (accessed on 3 May 2024).
- Copan Italia. Available online: https://www.copangroup.com/ (accessed on 3 May 2024).
- Abbott Molecular. Available online: https://www.molecular.abbott/content/dam/add/molecular/alinity-m-sars-cov-2-assay/us/53-608191R11%20Alinity%20m%20SARS%20AMP%20Kit%20PI%20EUA_lg016.pdf (accessed on 3 May 2024).
- Qiagen. Available online: https://www.qiagen.com/us (accessed on 3 May 2024).
- Thermo-Fisher Scientific. Available online: https://www.thermofisher.com/it/en/home.html (accessed on 3 May 2024).
- Bio-Rad. Available online: https://www.bio-rad.com/ (accessed on 3 May 2024).
- Corman, V.M.; Landt, O.; Kaiser, M.; Molenkamp, R.; Meijer, A.; Chu, D.K.; Bleicker, T.; Brünink, S.; Schneider, J.; Schmidt, M.L.; et al. Detection of 2019 novel coronavirus (2019-nCoV) by real-time RT-PCR. Euro Surveill 2020, 25, 2000045. [Google Scholar] [CrossRef]
- GraphPad Prism. Available online: https://www.graphpad.com/features (accessed on 3 May 2024).


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sberna, G.; Mija, C.; Lalle, E.; Rozera, G.; Matusali, G.; Carletti, F.; Girardi, E.; Maggi, F. Rapid Determination of SARS-CoV-2 Integrity and Infectivity by Using Propidium Monoazide Coupled with Digital Droplet PCR. Int. J. Mol. Sci. 2024, 25, 6156. https://doi.org/10.3390/ijms25116156
Sberna G, Mija C, Lalle E, Rozera G, Matusali G, Carletti F, Girardi E, Maggi F. Rapid Determination of SARS-CoV-2 Integrity and Infectivity by Using Propidium Monoazide Coupled with Digital Droplet PCR. International Journal of Molecular Sciences. 2024; 25(11):6156. https://doi.org/10.3390/ijms25116156
Chicago/Turabian StyleSberna, Giuseppe, Cosmina Mija, Eleonora Lalle, Gabriella Rozera, Giulia Matusali, Fabrizio Carletti, Enrico Girardi, and Fabrizio Maggi. 2024. "Rapid Determination of SARS-CoV-2 Integrity and Infectivity by Using Propidium Monoazide Coupled with Digital Droplet PCR" International Journal of Molecular Sciences 25, no. 11: 6156. https://doi.org/10.3390/ijms25116156
APA StyleSberna, G., Mija, C., Lalle, E., Rozera, G., Matusali, G., Carletti, F., Girardi, E., & Maggi, F. (2024). Rapid Determination of SARS-CoV-2 Integrity and Infectivity by Using Propidium Monoazide Coupled with Digital Droplet PCR. International Journal of Molecular Sciences, 25(11), 6156. https://doi.org/10.3390/ijms25116156