

Revealing Spermatogenesis in Smooth-Hound Sharks Mustelus mustelus: Insights into the Morphological and Macromolecular Composition of Spermatogenic Cells
 ,
,  , , , , , ,
, , , , , ,  and
and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results and Discussion
2.1. First Stage: Differentiation of Germ Cells to Spermatogonia
2.2. Second Stage: Early Cyst Proliferation
2.3. Third Stage: Primary Spermatocytes
2.4. Fourth Stage: Secondary Spermatocytes
2.5. Fifth Stage: Spermatid Types
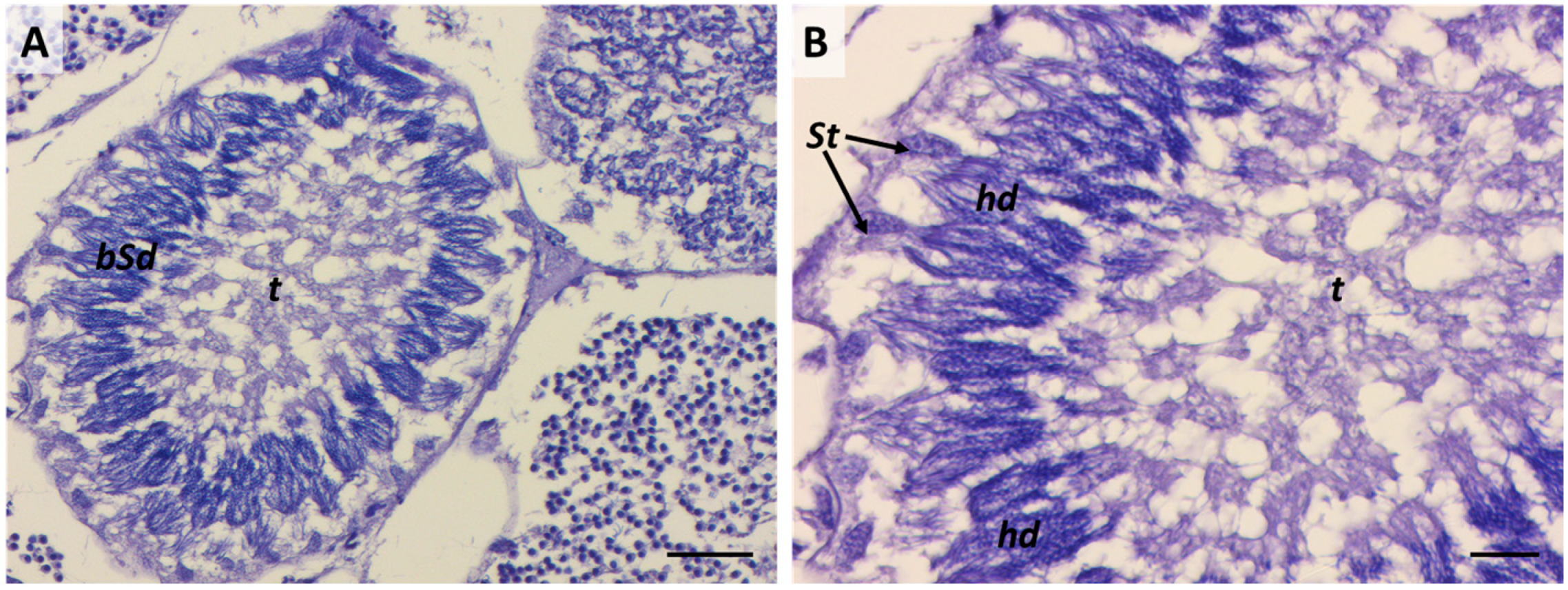
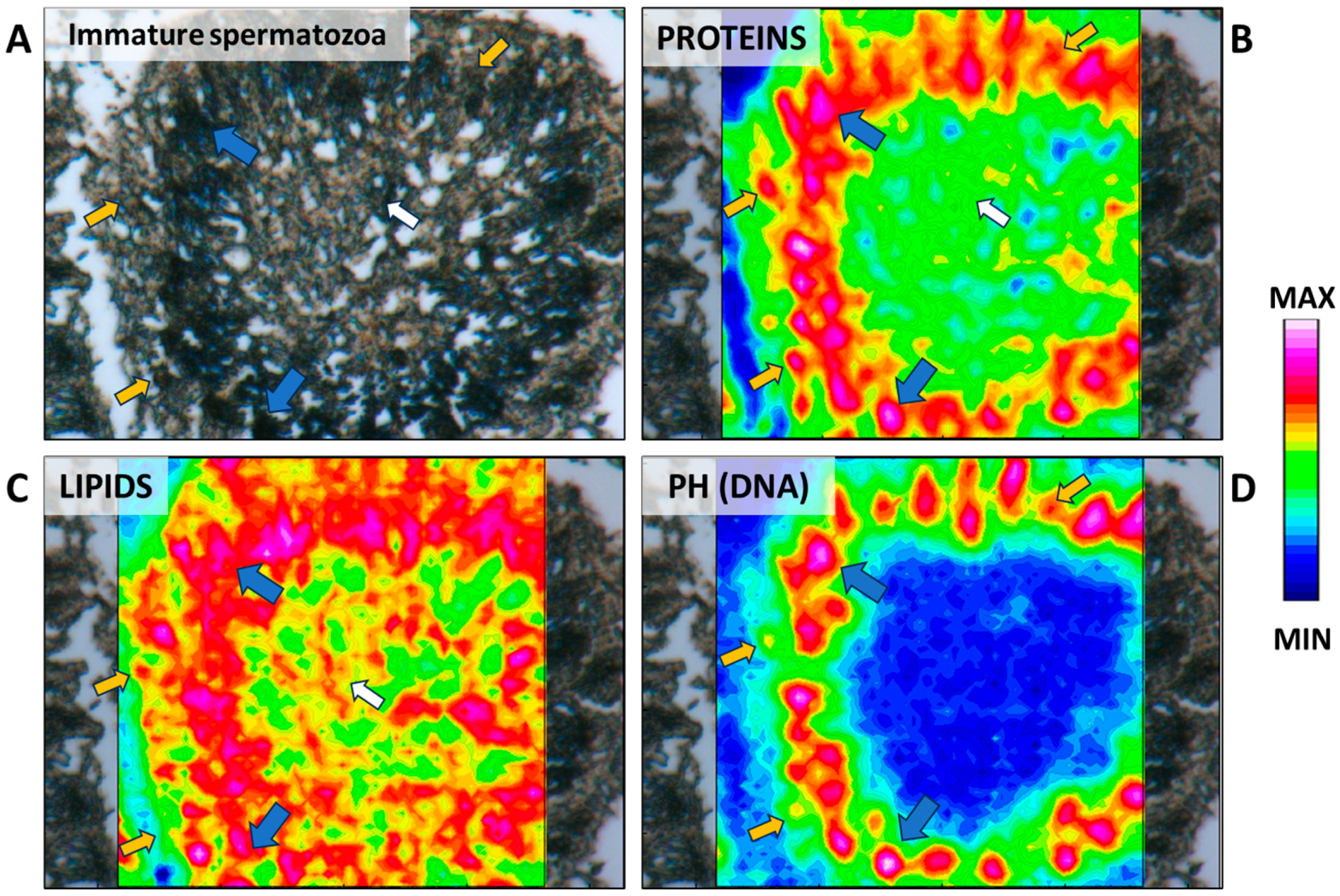
2.6. Sixth Stage: “Tree-Frond-Shaped” Immature Spermatozoa
2.7. Seventh Stage: Mature Spermatozoa
3. Materials and Methods
3.1. Sample Collection
3.2. Histological Analysis
3.3. Fourier-Transform InfraRed Imaging (FTIRI) Analysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schulz, R.W.; de França, L.R.; Lareyre, J.J.; LeGac, F.; Chiarini-Garcia, H.; Nobrega, R.H.; Miura, T. Spermatogenesis in fish. Gen. Comp. Endocrinol. 2010, 165, 390–411. [Google Scholar] [CrossRef] [PubMed]
- Carrier, J.C.; Musick, J.A.J.; Heithaus, M.R. Biology of Sharks and Their Relatives; CRC Press: Boca Raton, FL, USA, 2004; ISBN 9780203491317. [Google Scholar]
- Dulvy, N.K.; Pacoureau, N.; Rigby, C.L.; Pollom, R.A.; Jabado, R.W.; Ebert, D.A.; Finucci, B.; Pollock, C.M.; Cheok, J.; Derrick, D.H.; et al. Overfishing drives over one-third of all sharks and rays toward a global extinction crisis. Curr. Biol. 2021, 31, 4773–4787.e8. [Google Scholar] [CrossRef] [PubMed]
- Giovos, I.; Aga Spyridopoulou, R.N.; Doumpas, N.; Glaus, K.; Kleitou, P.; Kazlari, Z.; Katsada, D.; Loukovitis, D.; Mantzouni, I.; Papapetrou, M.; et al. Approaching the “real” state of elasmobranch fisheries and trade: A case study from the Mediterranean. Ocean Coast. Manag. 2021, 211, 105743. [Google Scholar] [CrossRef]
- Dulvy, N.K.; Fowler, S.L.; Musick, J.A.; Cavanagh, R.D.; Kyne, P.M.; Harrison, L.R.; Carlson, J.K.; Davidson, L.N.K.; Fordham, S.V.; Francis, M.P.; et al. Extinction risk and conservation of the world’s sharks and rays. Elife 2014, 3, e00590. [Google Scholar] [CrossRef] [PubMed]
- Colloca, F.; Enea, M.; Ragonese, S.; Di Lorenzo, M. A century of fishery data documenting the collapse of smooth-hounds (Mustelus spp.) in the Mediterranean Sea. Aquat. Conserv. Mar. Freshw. Ecosyst. 2017, 27, 1145–1155. [Google Scholar] [CrossRef]
- Dulvy, N.K.; Allen, D.J.; Ralph, G.M.; Walls, R.H.L. The Conservation Status of Sharks, Rays and Chimaeras in the Mediterranean Sea; IUCN: Malaga, Spain, 2016. [Google Scholar]
- Giovos, I.; Arculeo, M.; Doumpas, N.; Katsada, D.; Maximiadi, M.; Mitsou, Ε.; Paravas, V.; Naasan Aga-Spyridopoulou, R.; Stoilas, V.O.; Tiralongo, F.; et al. Assessing multiple sources of data to detect illegal fishing, trade and mislabelling of elasmobranchs in Greek markets. Mar. Policy 2020, 112, 103730. [Google Scholar] [CrossRef]
- Nuez, I.; Gazo, M.; Cardona, L. A closer look at the bycatch of medium-sized and large sharks in the northern Catalan coast (north-western Mediterranean Sea): Evidence of an ongoing decline? Aquat. Conserv. Mar. Freshw. Ecosyst. 2021, 31, 2369–2380. [Google Scholar] [CrossRef]
- Vélez-Zuazo, X.; Agnarsson, I. Shark tales: A molecular species-level phylogeny of sharks (Selachimorpha, Chondrichthyes). Mol. Phylogenet. Evol. 2011, 58, 207–217. [Google Scholar] [CrossRef]
- Saïdi, B.; Bradaï, M.N.; Bouaïn, A. Reproductive biology of the smooth-hound shark Mustelus mustelus (L.) in the Gulf of Gabès (south-central Mediterranean Sea). J. Fish Biol. 2008, 72, 1343–1354. [Google Scholar] [CrossRef]
- Nieto, A.; Ralph, G.M.; Comeros-Rynal, M.T.; Heessen, H.; Rijnsdorp, A.D. European Red List of Marine Fishes; Publications Office of the European Union: Luxembourg, 2015; ISBN 9789279454127. [Google Scholar]
- Barausse, A.; Correale, V.; Curkovic, A.; Finotto, L.; Riginella, E.; Visentin, E.; Mazzoldi, C. The role of fisheries and the environment in driving the decline of elasmobranchs in the northern Adriatic Sea. ICES J. Mar. Sci. 2014, 71, 1593–1603. [Google Scholar] [CrossRef]
- Barausse, A.; Michieli, A.; Riginella, E.; Palmeri, L.; Mazzoldi, C. Long-term changes in community composition and life-history traits in a highly exploited basin (northern Adriatic Sea): The role of environment and anthropogenic pressures. J. Fish Biol. 2011, 79, 1453–1486. [Google Scholar] [CrossRef] [PubMed]
- Waltrick, D.S.; Simpfendorfer, C.A.; Awruch, C.A. A review on the morphology of ovarian follicles in elasmobranchs: A case study in Rhizoprionodon taylori. J. Morphol. 2017, 278, 486–499. [Google Scholar] [CrossRef] [PubMed]
- Davenport, I.R.; Weaver, A.L.; Wourms, J.P. A novel set of structures within the elasmobranch, ovarian follicle. J. Morphol. 2011, 272, 557–565. [Google Scholar] [CrossRef] [PubMed]
- Lance, V.; Callard, I.P. A histochemical study of ovarian function in the ovoviviparous elasmobranch, Squalus acanthias. Gen. Comp. Endocrinol. 1969, 13, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Fasano, S.; Pierantoni, R.; Chieffi, G. Reproductive biology of elasmobranchs with emphasis on endocrines. J. Exp. Zool. 1989, 252, 53–61. [Google Scholar] [CrossRef]
- Marino, I.A.M.; Riginella, E.; Gristina, M.; Rasotto, M.B.; Zane, L.; Mazzoldi, C. Multiple paternity and hybridization in two smooth-hound sharks. Sci. Rep. 2015, 5, 12919. [Google Scholar] [CrossRef]
- McClusky, L.M. Coordination of spermatogenic processes in the testis: Lessons from cystic spermatogenesis. Cell Tissue Res. 2012, 349, 703–715. [Google Scholar] [CrossRef] [PubMed]
- Bobe, J.; Labbé, C. Egg and sperm quality in fish. Gen. Comp. Endocrinol. 2010, 165, 535–548. [Google Scholar] [CrossRef] [PubMed]
- Montealegre-Quijano, S.; Cardoso, A.T.C.; Silva, R.Z.; Kinas, P.G.; Vooren, C.M. Sexual development, size at maturity, size at maternity and fecundity of the blue shark Prionace glauca (Linnaeus, 1758) in the Southwest Atlantic. Fish. Res. 2014, 160, 18–32. [Google Scholar] [CrossRef]
- Castro, J.I. The biology of the nurse shark, Ginglymostoma cirratum, off the Florida east coast and the Bahama Islands. Environ. Biol. Fishes 2000, 58, 1–22. [Google Scholar] [CrossRef]
- Francis, M.P.; Duffy, C. Length at maturity in three pelagic sharks (Lamna nasus, Isurus oxyrinchus, and Prionace glauca) from New Zealand. Fish. Bull. 2005, 103, 489–500. [Google Scholar]
- Segura, A.M.; Milessi, A.C.; Vögler, R.; Galván-Magaña, F.; Muggeo, V. The determination of maturity stages in male elasmobranchs (Chondrichthyes) using a segmented regression of clasper length on total length. Can. J. Fish. Aquat. Sci. 2013, 70, 830–833. [Google Scholar] [CrossRef]
- Jeanne, F.; Bernay, B.; Sourdaine, P. Comparative Proteome Analysis of Four Stages of Spermatogenesis in the Small-Spotted Catshark (Scyliorhinus canicula), Using High-Resolution NanoLC-ESI-MS/MS. J. Proteome Res. 2023, 22, 2477–2492. [Google Scholar] [CrossRef]
- McClusky, L.M. The caspase-dependent apoptosis gradient in the testis of the blue shark, Prionace glauca. Reproduction 2013, 145, 297–310. [Google Scholar] [CrossRef]
- Gračan, R.; Lacković, G. Histological and Morphological Aspects of Reproduction in Male Blackspotted Smooth-Hound Mustelus punctulatus in the Adriatic Sea (Eastern Mediterranean Sea). J. Mar. Biol. 2016, 2016, 3438678. [Google Scholar] [CrossRef]
- Gomes do Rêgo, M.; Fitzpatrick, J.L.; Hissa, V.; Hazin, F.; Araujo, M.L.G.; Barros, M.E.G.; Evêncio Neto, J. Comparison of the Morphology and Histomorphometry of Spermatogenic Cyst of Three Sharks Species With Diametric Testes. Anat. Rec. 2016, 299, 759–768. [Google Scholar] [CrossRef]
- Farrell, E.D.; Mariani, S.; Clarke, M.W. Reproductive biology of the starry smooth-hound shark Mustelus asterias: Geographic variation and implications for sustainable exploitation. J. Fish Biol. 2010, 77, 1505–1525. [Google Scholar] [CrossRef]
- Conrath, C.L.; Musick, J.A. Reproductive biology of the smooth dogfish, Mustelus canis, in the northwest Atlantic Ocean. Environ. Biol. Fishes 2002, 64, 367–377. [Google Scholar] [CrossRef]
- Park, J.C.; Lee, J.H.; Kodama, K.; Urushitani, H.; Ohta, Y.; Horiguchi, T. Structure of the intratesticular duct system for sperm emission in the starspotted smooth-hound Mustelus manazo. Fish. Sci. 2013, 79, 203–211. [Google Scholar] [CrossRef]
- França, L.R.; Nóbrega, R.H.; Morais, R.D.V.S.; De Castro Assis, L.H.; Schulz, R.W. Sertoli cell structure and function in anamniote vertebrates. In Sertoli Cell Biology; Academic Press: New York, NY, USA, 2015; pp. 385–407. [Google Scholar]
- Yoshida, S. From cyst to tubule: Innovations in vertebrate spermatogenesis. Wiley Interdiscip. Rev. Dev. Biol. 2016, 5, 119–131. [Google Scholar] [CrossRef] [PubMed]
- Loir, M.; Sourdaine, P.; Mendis-Handagama, S.M.L.C.; Jégou, B. Cell-cell interactions in the testis of teleosts and elasmobranchs. Microsc. Res. Tech. 1995, 32, 533–552. [Google Scholar] [CrossRef]
- Poulakis, G.R.; Grier, H.J. Ontogenetic testicular development and spermatogenesis in rays: The Cownose Ray, Rhinoptera bonasus, as a model. Environ. Biol. Fishes 2014, 97, 1013–1029. [Google Scholar] [CrossRef]
- Girard, M.; Rivalan, P.; Sinquin, G. Testis and sperm morphology in two deep-water squaloid sharks, Centroscymnus coelolepis and Centrophorus squamosus. J. Fish Biol. 2000, 57, 1575–1589. [Google Scholar]
- Rêgo, M.G.; Fitzpatrick, J.; Hazin, F.H.V.; de Araújo, M.L.G.; da Silveira, L.M.; Oliveira, P.G.V.; Evêncio-Neto, J. Characterization of testicular morphology and spermatogenesis in the nurse sharks Ginglymostoma cirratum (Bonnaterre, 1788). Zoomorphology 2015, 134, 117–123. [Google Scholar] [CrossRef]
- Marina, P.; Annamaria, L.; Barbara, D.; Loredana, R.; Piero, A.; Francesco, A. Fine structure of leydig and sertoli cells in the testis of immature and mature spotted ray Torpedo marmorata. Mol. Reprod. Dev. 2002, 63, 192–201. [Google Scholar] [CrossRef]
- Sourdaine, P.; Gautier, A.; Gribouval, L. Spermatogenesis and spermiogenesis in elasmobranchs, a short overview. In Encyclopedia of Reproduction; Elsevier: Amsterdam, The Netherlands, 2018; pp. 305–312. ISBN 9780128151457. [Google Scholar]
- McClusky, L.M. Several routes of cell death to secondary necrosis in the elasmobranch testis. Apoptosis 2022, 27, 454–464. [Google Scholar] [CrossRef]
- Jones, R.C.; Lin, M. Structure and functions of the genital ducts of the male Port Jackson shark, Heterodontus portusjacksoni. Environ. Biol. Fishes 1993, 38, 127–138. [Google Scholar] [CrossRef]
- Pudney, J.; Callard, G.V. Sertoli cell cytoplasts in the semen of the spiny dogfish Squalus acanthias. Tissue Cell 1986, 18, 375–382. [Google Scholar] [CrossRef]
- Golani, D.; Ozturk, B.; Basusta, N. Fishes of the Eastern Mediterranean. Isr. J. Aquac. Bamidgeh 2007, 59, 121. [Google Scholar] [CrossRef]
- Ozcan, E.I.; Başusta, N. Preliminary study on age, growth and reproduction of mustelus mustelus (Elasmobranchii: Carcharhiniformes: Triakidae) inhabiting the gulf of Iskenderun, north-eastern mediterranean sea. Acta Ichthyol. Piscat. 2018, 48, 27–36. [Google Scholar] [CrossRef]
- Chemello, G.; Trotta, E.; Notarstefano, V.; Papetti, L.; Di Renzo, L.; Matiddi, M.; Silvestri, C.; Carnevali, O.; Gioacchini, G. Microplastics evidence in yolk and liver of loggerhead sea turtles (Caretta caretta), a pilot study. Environ. Pollut. 2023, 337, 122589. [Google Scholar] [CrossRef] [PubMed]














Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chemello, G.; De Santis, L.J.; Trotta, E.; Zarantoniello, M.; Santoni, C.; Maradonna, F.; Olivotto, I.; Giorgini, E.; Gioacchini, G. Revealing Spermatogenesis in Smooth-Hound Sharks Mustelus mustelus: Insights into the Morphological and Macromolecular Composition of Spermatogenic Cells. Int. J. Mol. Sci. 2024, 25, 6230. https://doi.org/10.3390/ijms25116230
Chemello G, De Santis LJ, Trotta E, Zarantoniello M, Santoni C, Maradonna F, Olivotto I, Giorgini E, Gioacchini G. Revealing Spermatogenesis in Smooth-Hound Sharks Mustelus mustelus: Insights into the Morphological and Macromolecular Composition of Spermatogenic Cells. International Journal of Molecular Sciences. 2024; 25(11):6230. https://doi.org/10.3390/ijms25116230
Chicago/Turabian StyleChemello, Giulia, Lorenzo Jacopo De Santis, Erica Trotta, Matteo Zarantoniello, Chiara Santoni, Francesca Maradonna, Ike Olivotto, Elisabetta Giorgini, and Giorgia Gioacchini. 2024. "Revealing Spermatogenesis in Smooth-Hound Sharks Mustelus mustelus: Insights into the Morphological and Macromolecular Composition of Spermatogenic Cells" International Journal of Molecular Sciences 25, no. 11: 6230. https://doi.org/10.3390/ijms25116230
APA StyleChemello, G., De Santis, L. J., Trotta, E., Zarantoniello, M., Santoni, C., Maradonna, F., Olivotto, I., Giorgini, E., & Gioacchini, G. (2024). Revealing Spermatogenesis in Smooth-Hound Sharks Mustelus mustelus: Insights into the Morphological and Macromolecular Composition of Spermatogenic Cells. International Journal of Molecular Sciences, 25(11), 6230. https://doi.org/10.3390/ijms25116230