Abstract
Although glaucoma is a leading cause of irreversible blindness worldwide, its pathogenesis is incompletely understood, and intraocular pressure (IOP) is the only modifiable risk factor to target the disease. Several associations between the gut microbiome and glaucoma, including the IOP, have been suggested. There is growing evidence that interactions between microbes on the ocular surface, termed the ocular surface microbiome (OSM), and tear proteins, collectively called the tear proteome, may also play a role in ocular diseases such as glaucoma. This study aimed to find characteristic features of the OSM and tear proteins in patients with glaucoma. The whole-metagenome shotgun sequencing of 32 conjunctival swabs identified Actinobacteria, Firmicutes, and Proteobacteria as the dominant phyla in the cohort. The species Corynebacterium mastitidis was only found in healthy controls, and their conjunctival microbiomes may be enriched in genes of the phospholipase pathway compared to glaucoma patients. Despite these minor differences in the OSM, patients showed an enrichment of many tear proteins associated with the immune system compared to controls. In contrast to the OSM, this emphasizes the role of the proteome, with a potential involvement of immunological processes in glaucoma. These findings may contribute to the design of new therapeutic approaches targeting glaucoma and other associated diseases.
1. Introduction
Glaucoma is a worldwide leading cause of irreversible vision loss. It is characterized by the gradual degeneration of retinal ganglion cells and their axons [1,2], associated with narrowing of the visual field [3]. In 2013, the global occurrence of glaucoma among individuals aged 40 to 80 years was approximately 3.5%. The total number of glaucoma patients was nearly 80 million in 2020, and it is projected to surpass 100 million by the year 2040 [4,5].
Glaucoma can be divided into several subtypes, including primary open-angle glaucoma (POAG), primary angle-closure glaucoma (PACG), and secondary glaucoma. In POAG, serious damage may be caused to the optic nerve and the trabecular meshwork, which is the main pathway for drainage of aqueous humor and whose dysfunction leads to intraocular pressure (IOP) elevation [6]. The most common form of secondary glaucoma is pseudoexfoliation glaucoma (PEXG), caused by the deposition of fibrillar protein aggregates (PEX fibrils) on the anterior and posterior eye tissues [7].
IOP is the only modifiable risk factor that can influence the progression of glaucoma [8]. Treatment options to reduce IOP includes topical and oral therapies, laser treatments targeting the trabecular meshwork or ciliary body, and incisional surgery [2,5,9]. In addition to IOP elevation, the risk factors for glaucoma include advancing age, gender, diet, obesity, depression, and anxiety [5]. Since there are associations between the gut microbiome and these risk factors [10,11,12,13,14,15], the gut microbiome has gained much attention in the pathophysiology of glaucoma. Several studies identified significant changes in the microbial composition of glaucoma patients compared to healthy controls, with a potential influence of neuroinflammation and autoimmunity [16].
However, while there are associations between gut microbiome dysbiosis [17,18,19] and the development of glaucoma [5], as well as other diseases, the role of the ocular surface microbiome (OSM) in glaucoma has not been explored so far. The OSM refers to the collective community of microorganisms and their genomes on the surface of the eye, including bacteria, viruses, and fungi [20,21,22,23,24,25,26,27]. The investigation of the OSM in this study stems from a gap in prior research, particularly the Human Microbiome Project, which primarily focused on characterizing microbiomes across various body sites but excluded the eye [28]. Unlike other areas explored, such as the skin, gastrointestinal tract, and oral cavity, the OSM has been relatively unexplored. Modern sequencing technologies are employed to gain a comprehensive understanding of the OSM, including its interactions with antimicrobial proteins in tear fluid [22] and involvement in eye diseases such as dry eye disease [29]. To gain insights into the complex interplay between the OSM, tear proteome, immune system processes, and glaucoma, the OSM and tear fluid of 16 glaucoma patients, compared to 16 healthy controls, were explored in this study. Furthermore, since the previous research has shown alterations in bacterial composition and diversity due to various types of eye drops, such as antibacterial agents and sodium hyaluronate [30,31], the influence of IOP-reducing eye drops on the OSM and its potential implications for ocular health were assessed.
Prebiotics, probiotics, fecal transplantation, and more recently postbiotics have been suggested as interventions to modulate the gut microbiome, thereby ameliorating associated conditions [32]. This study enables the identification of potential targets for glaucoma treatment by microbiome-altering approaches and aid in the development of personalized therapeutic strategies in the future [33].
2. Results
2.1. Demographic Data
A total of 32 conjunctival swabs, as well as the correspondent tear fluids, were collected from 16 glaucoma patients and 16 healthy controls. While the two groups did not show differences in the sex ratio (Table 1), there was a significant difference in age between the groups (p = 0.0046, Welch’s t test). Since previous studies have shown an age-dependence for both the OSM [34,35] and the tear proteome [36], we considered this difference in a Multivariate Association with Linear Models (MaAsLin2) analysis [37] (see Section 2.2 and Section 4.3) and screened for age-independent protein expressions (see Section 2.3 and Section 4.4). In the glaucoma group, there were eight patients with POAG and eight patients with PEXG. A subgroup analysis was performed for the OSM characterization (see Section 2.2).

Table 1.
Demographic information of study participants.
The ocular hypotensive therapy for glaucoma frequently incorporates active pharmaceutical ingredients such as alpha-agonists, beta-blockers, carbonic anhydrase inhibitors, or prostaglandin analogs to reduce IOP. In our cohort of 16 patients, the therapeutic regimens for glaucoma management revealed a distribution of ocular hypotensive agents as follows: prostaglandin analogs (14 patients), carbonic anhydrase inhibitors (13 patients), beta-blockers (15 patients) and alpha-agonists (6 patients; Table 2). Notably, the majority of patients were subjected to multiple agents concurrently, indicative of prevalent combination therapies in the management of glaucoma.
Table 2.
Topical intraocular pressure-lowering active ingredients of all 16 glaucoma patients.
2.2. Characterization of the Ocular Surface Microbiome in Glaucoma Patients
In total, 1.83 billion 150 bp paired-end reads with an average insert size of 350 bp were generated, with an average of 78.21 ± 22.46 (s.d.) million reads per sample. As expected and described in previous studies, most of these reads were of human origin due to the low abundance of the OSM [38]. After trimming and host-filtering, we kept about 331 million high-quality, non-human reads with an average of 6.94 ± 4.59 (s.d.) million per sample. After removing duplicated reads using Unique Molecular Identifiers (UMIs), we had a total of 4.27 million deduplicated reads for further analysis, with an average of 0.17 ± 0.14 (s.d.) million per sample. Since sequencing libraries from negative controls failed quality and quantity controls for sequencing, we concluded that contaminations were not present in a significant concentration.
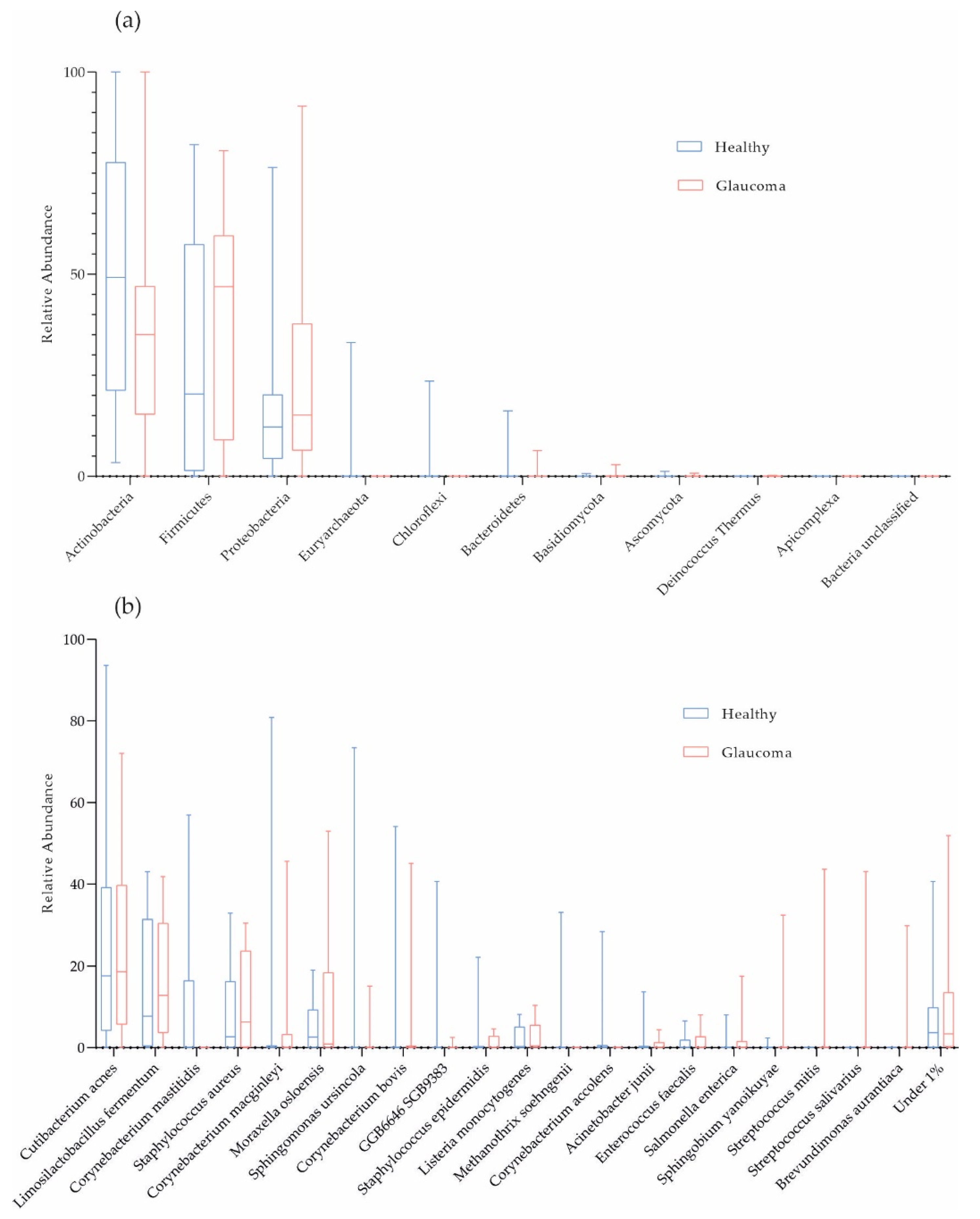
On average, 98.72% of the host-filtered reads were of bacterial origin. Consistent with previous studies, the phyla Actinobacteria (41.50%), Firmicutes (33.51%), and Proteobacteria (22.10%) dominated OSM composition in the cohort (Figure 1a). Cutibacterium acnes (24.17%), Limosilactobacillus fermentum (15.42%), and Staphylococcus aureus (9.84%; mean values) were the most abundant species in the cohort (Figure 1b). In Figure 1b, on average, we filtered 9.19% and 6.71% of the data for patients and controls, respectively, because these taxonomic units were present at less than 1% (low-abundant species termed “Under 1%”).
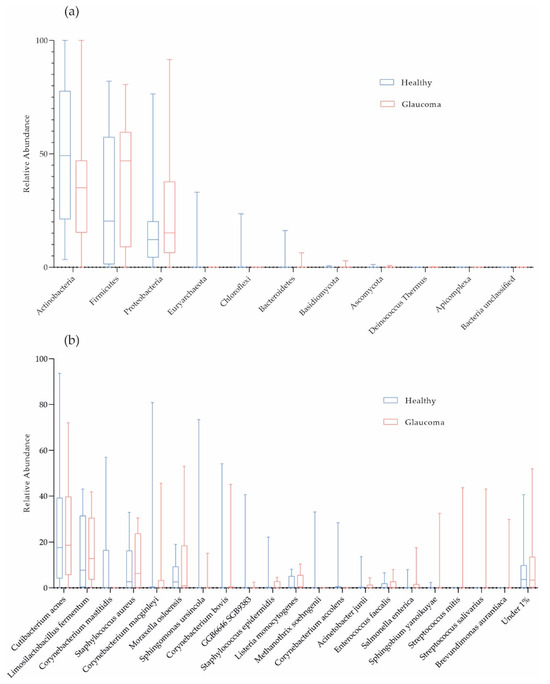
Figure 1.
Taxonomic characterization of ocular surface microbiome. Box plots for relative abundances at phylum (a) and species level (b) are shown (red, glaucoma, n = 16; blue, healthy controls, n = 16).
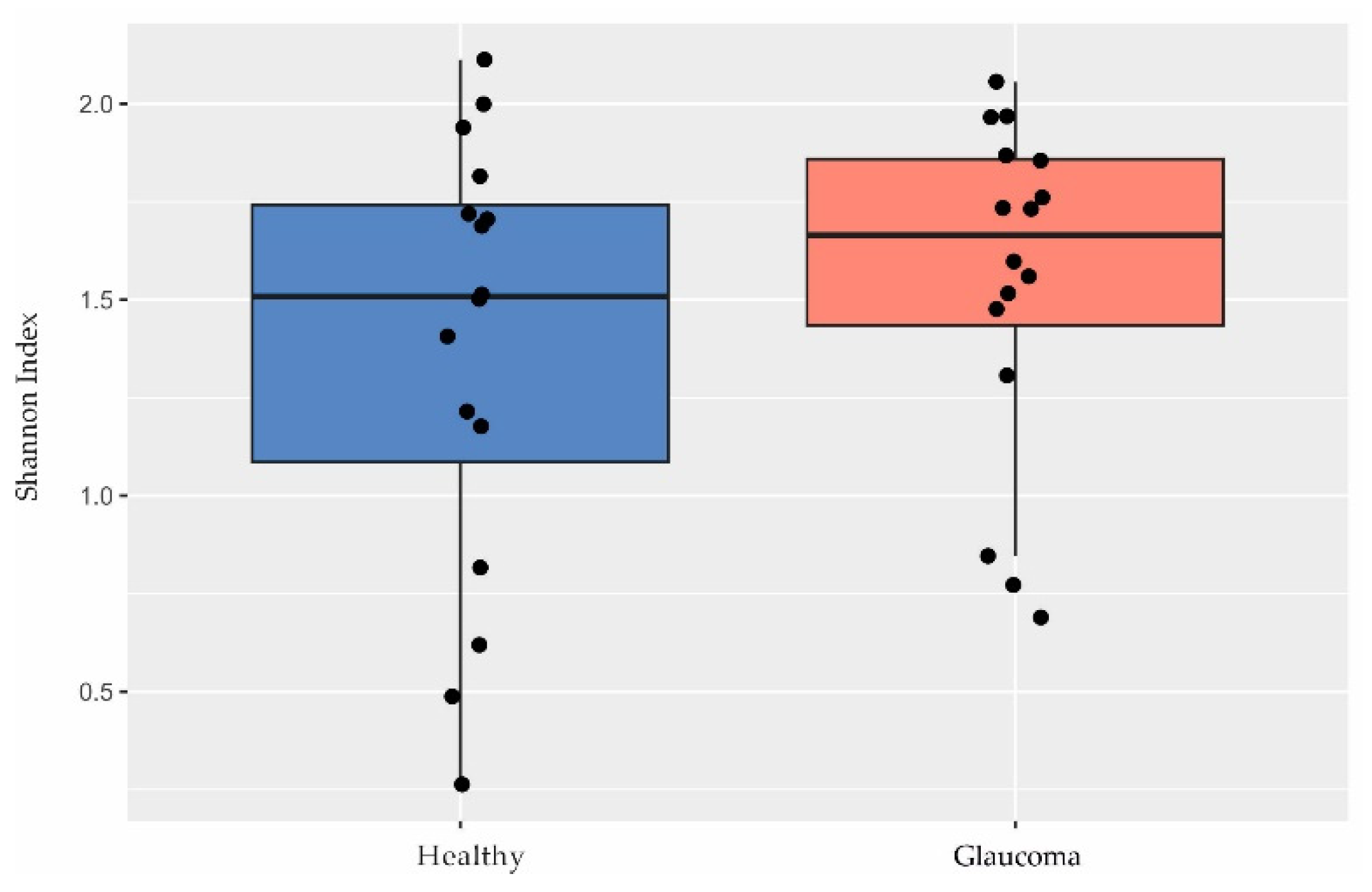
Although other studies have shown a lower microbial diversity in glaucoma [39,40,41], we found no differences in diversity between patients and controls based on the Shannon index (p = 0.35, Welch’s t test; Figure 2).
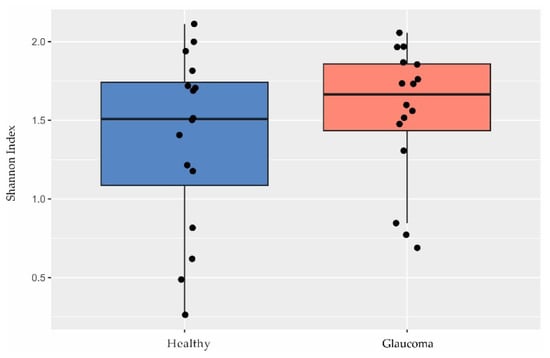
Figure 2.
Diversity in the ocular surface microbiome. There were no differences in Shannon index observed between glaucoma patients (n = 16) and controls (n = 16; p = 0.35, Welch’s t test).
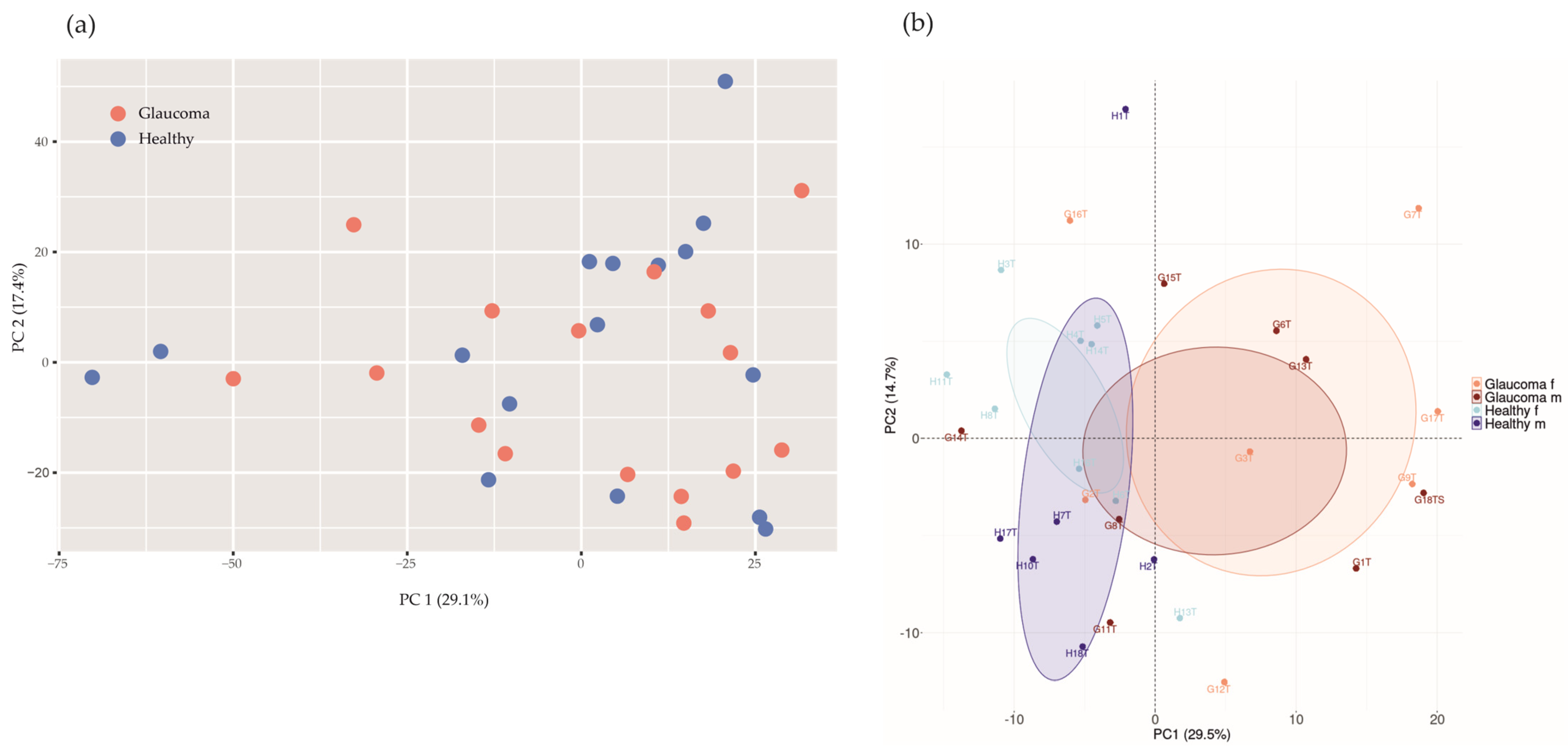
If applying a principal component analysis (PCA) with health status as the grouping variable, the glaucoma group would not separate from healthy controls based on differences in microbial abundances (p = 0.51, PERMANOVA, n repeat = 1000; Figure 3a).
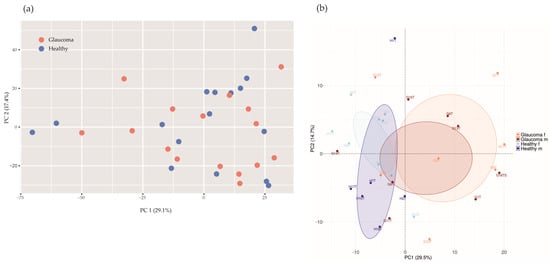
Figure 3.
Principal component analysis (PCA) of microbial abundances (a) and protein intensities (b). (a) PCA of microbial abundances did not separate glaucoma patients (red, n = 16) from healthy controls (blue, n = 16; p = 0.51, PERMANOVA, n repeat = 1000). (b) Log2−transformed protein group intensities before imputation were used for analysis. PCA separates glaucoma patients (n = 16) from controls (n = 16; p = 0.0060, PERMANOVA, n repeat = 1000). Orange, female glaucoma patients (n = 7); red, male glaucoma patients (n = 9); light blue, healthy control females (n = 10); blue, healthy control males (n = 6).
To further examine taxonomic and functional features of the OSM in glaucoma, we used MaAsLin2 to investigate whether the relative abundances of microbial taxa and pathways were associated with glaucoma and/or demographic data.
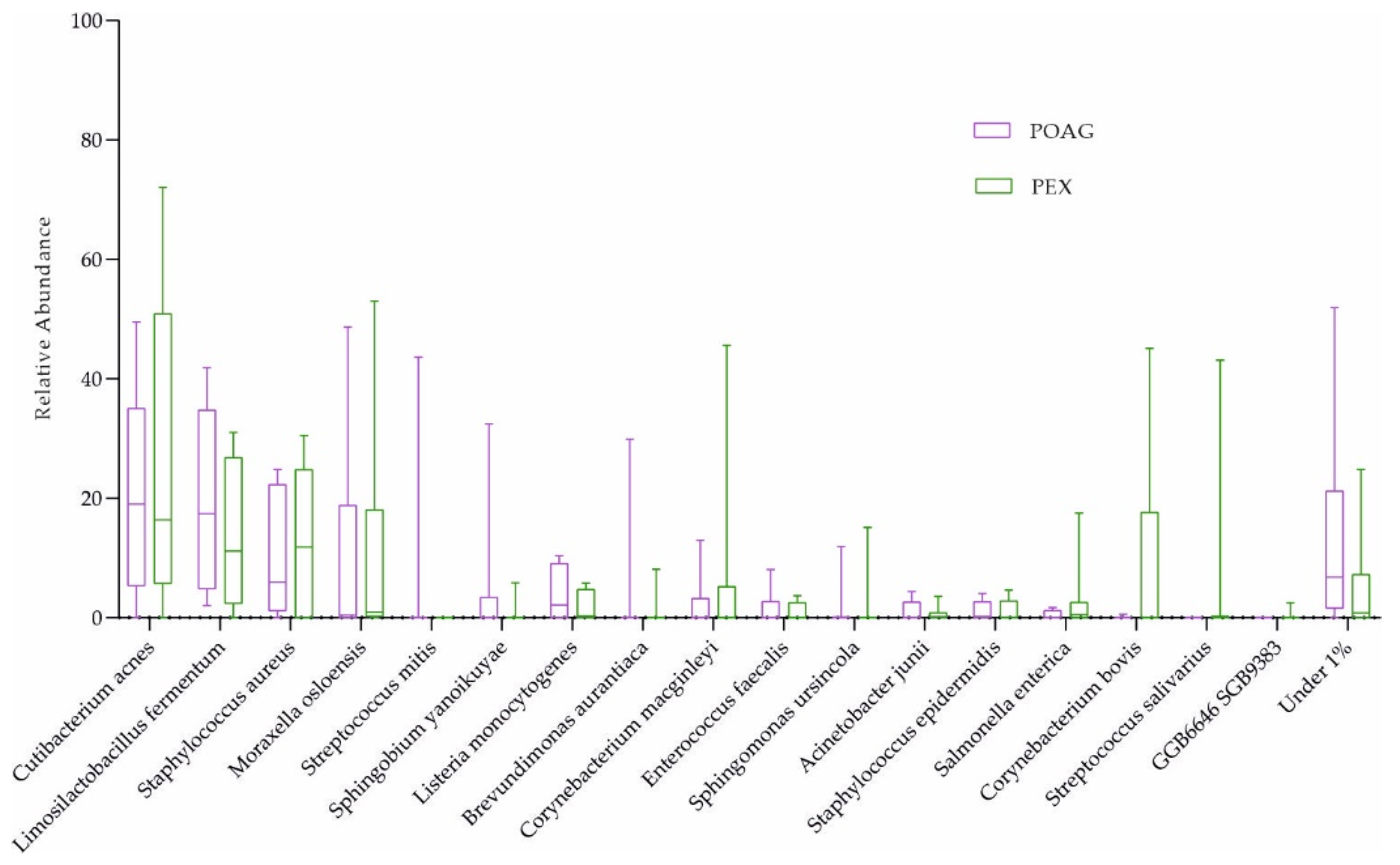
Corynebacterium mastitidis was absent in all glaucoma patients, but it was present in 7 out of 16 controls (Figure 1b and Table 3). However, there were no significant differences in microbial abundances between the subgroups (combination of four active ingredients of IOP-lowering eye drops including alpha-agonists, versus three active ingredients, versus no eye drops; Table 2; and POAG versus PEXG patients; Figure 4). For functional features, the ocular microbiomes of controls seem to be enriched in genes of the phospholipase pathway (q = 0.14).
Table 3.
Number of Corynebacterium mastitidis-positive samples. Contingency table, p = 0.0068 (two-tailed Fisher’s exact test).
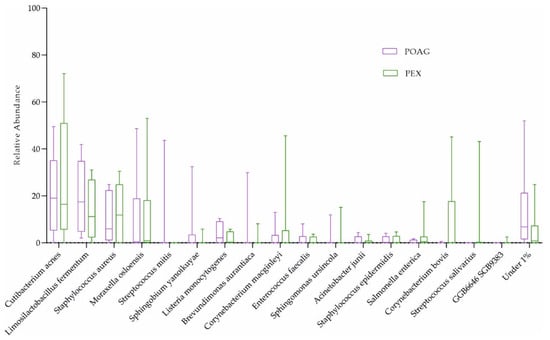
Figure 4.
The ocular surface microbiome of POAG and PEXG patients. Box plots for relative abundances of all species are shown. PEXG, pseudoexfoliation glaucoma (n = 8); POAG, primary open-angle glaucoma (n = 8).
2.3. Functional Annotation of the Tear Proteome in Glaucoma
We quantified 2250 human tear proteins. Applying a PCA based on log2-transformed protein group intensities, there are two clusters according to study groups (glaucoma versus controls; Figure 3b), suggesting a glaucoma-specific protein expression in tear fluids.
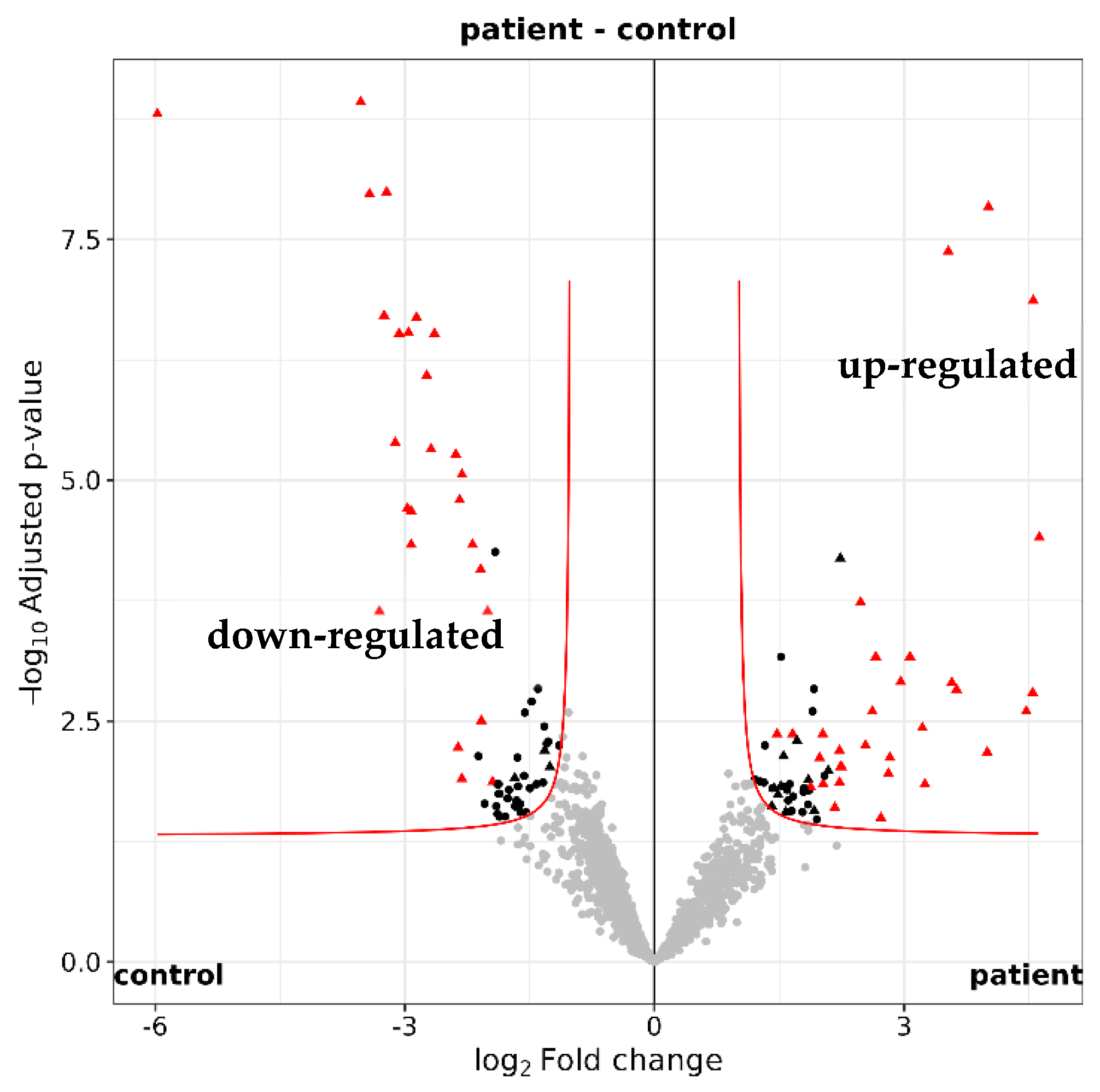
Among the 2250 identified tear proteins, 123 proteins were different in quantity between glaucoma patients and controls (according to imputed iMAX LFQ (label-free quantification) values, maximum adjusted p-value < 0.05 and fold change ≥2 or ≤−2; Figure 5 and Supplementary Table S1). Using STRING version 12.0, a functional enrichment analysis showed that many of the differentially expressed proteins are involved in biological processes associated with the immune system (Table 4).
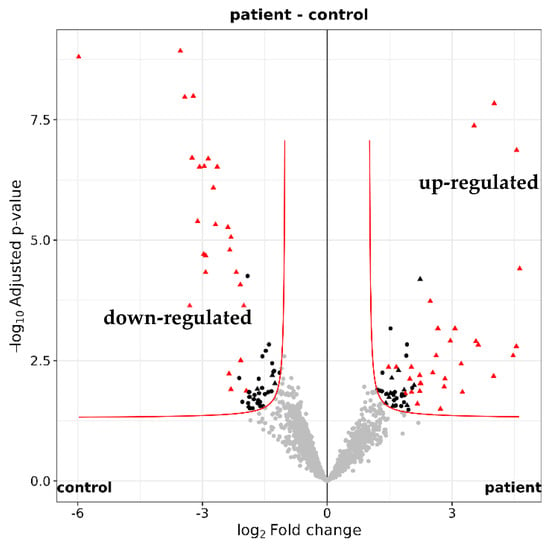
Figure 5.
Quantification of the tear proteome. Volcano plot illustrating the 2250 identifed tear proteins. A total of 123 proteins with a maximum adjusted p value of 0.05 were either up-regulated (log2 fold change ≥ 1) or down-regulated (log2 fold change ≤ −1) in glaucoma patients (n = 16) versus healthy controls (n = 16).
Table 4.
Functional annotation of tear proteins in glaucoma. Functional enrichment analysis of differentially expressed proteins between glaucoma patients and controls (based on iMAX LFQ values). The top first GO (gene ontology) terms are shown (sorted by false discovery rate).
3. Discussion
3.1. Implications of the Ocular Surface Microbiome in Glaucoma
Glaucoma, a prevalent and irreversible vision-impairing condition, is characterized by the gradual decline of retinal ganglion cells and their axons [1,2], resulting in narrowing of the visual field [3]. Over the past decade, there has been an increasing focus on exploring the connection between microbiota and glaucoma [5,6,42,43]. Microbiota imbalance (dysbiosis) triggered by various glaucoma risk factors, including aging [44], obesity [45], and depression [46], can lead to disruptions in metabolic, immune, and inflammatory processes. These disruptions are crucial mechanisms contributing to the development of the disease [5,43,45,47,48]. This study aimed to investigate the OSM and tear proteome in glaucoma patients compared to sex-matched healthy controls. The OSM, comprising microorganisms and their genomes on the eye’s surface, has been relatively unexplored compared to other body sites [20,28].
Several studies corroborate the prevalence of our top three bacterial species at phylum level, validating their prominence in ocular microbiomes [22,29,38,49,50,51,52,53]. Glaucoma patients and controls shared Actinobacteria, Proteobacteria, and Firmicutes as the predominant phyla in their OSMs, with Actinobacteria being the most abundant in our cohort. Their dominance underscores their potential significance in shaping the OSM. On species level, Corynebacterium mastitidis was only found in 7 out of 16 healthy controls but not in any of the glaucoma patients. C. mastitidis is a bacterium known for its commensal presence on mucous membranes, particularly on the ocular surface. It has been associated with promoting beneficial local immune responses [54]. The absence of C. mastitidis in glaucoma patients raises the possibility that its presence may play a stabilizing role in ocular health, suggesting a potential link between its absence and the disease. In an experimental model simulating ocular surface disease in mice, researchers found a persistent colonization of the ocular mucosa by C. mastitidis. They showed a causal relationship between the presence of C. mastitidis in the eyes and the promotion of beneficial local immune responses [55]. In another mouse model, C. mastitidis has been associated with the development of inflammation and abscesses, playing a role in protecting the eyes by releasing antimicrobials. It was demonstrated that C. mastitidis is found in both humans and mice [54]. St Leger et al. further showed that C. mastitidis triggers γδ T cells to produce interleukin 17, leading to the release of these antimicrobial molecules into tears. This protective mechanism provides a defense against invasive Candida albicans and Pseudomonas aeruginosa infections. These findings underscore the existence of true commensalism at the ocular surface, emphasizing the crucial role of local γδ T cell responses and suggesting potential broader implications for ocular surface diseases [56].
Functional analysis of the OSM identified a phospholipase pathway that may be increased in healthy controls compared to patients. The relationship between phospholipids and glaucoma has been explored, highlighting phospholipase A2 (PLA2) as a contributor to neuronal cell death, particularly in the trabecular meshwork (TM) of glaucoma patients. The suppression of lipid mediators generated by PLA2 activity presents a potential therapeutic target for treating glaucoma. However, the study emphasizes the need for caution in developing safe and isoform-specific PLA2-targeting drugs, to minimize the associated risks and side effects [57]. The potental inverse up-regulation in our study needs further investigation.
3.2. The Influence of Intraocular Pressur-Lowering Eye Drops on the Ocular Microbiome
In addition to exploring microbial composition and function in glaucoma, our study investigated the influence of IOP-lowering eye drops on the OSM. Although previous studies have demonstrated alterations in ocular microbiota following the use of various eye drops [30,31], we did not detect any influence of eye drop composition on the OSM.
A randomized trial comparing Carboxymethylcellulose to Polyethylene Glycol artificial tears found specific changes at genus level, including an increase in Bacteroidota and Actinobacteriota with Carboxymethylcellulose treatment [58]. However, overall diversity of the OSM was not significantly altered by these interventions. Limited research exists on the impact of IOP-lowering eye drops on the OSM, emphasizing the importance of further investigation. One study on the prolonged use of prostaglandin analogs in glaucoma treatment found a stable OSM among participants but lacked a healthy control group for direct comparison [59]. Similarly, a study comparing eyelid microbiomes in glaucoma patients found no association between microbiome composition and the use of preservative-containing eyedrops [60]. Furthermore, a study assessing the impact of topical antiglaucomatous eye drops on conjunctival flora in glaucoma patients and healthy controls revealed a significant increase in bacterial isolates in glaucoma patients, particularly Staphylococcus hominis and Candida spp. [61]. However, comparing conjunctival bacterial cultures in patients undergoing glaucoma and cataract surgery showed no significant difference in positive bacterial growth between the two groups, suggesting that glaucoma medications or their preservatives may not significantly alter the OSM [62]. In a research study that examined individuals with asymmetrical glaucoma and treated only one eye with preserved topical glaucoma therapy containing benzalkonium chloride, it was discovered that the OSM in the treated eye displayed a wide variety of Gram-negative bacteria. This differed significantly from the predominantly Gram-positive microbes found in healthy controls, suggesting a possible connection between microbial alterations and inflammation of the ocular surface, as well as changes in tear film and protein synthesis [63]. Overall, while various eye drops, including those used for glaucoma treatment, can impact the OSM, the extent of these changes and their clinical implications warrant further investigation. Therefore, continued research is essential to elucidate the potential systemic effects of ocular treatments and to optimize care for patients with ocular conditions.
3.3. Implications of the Tear Proteome and the Immune System in Glaucoma
Apart from the exclusive presence of C. mastitidis in controls and its potental association with ocular homeostasis, the OSMs from patients and controls seem to be comparable. This suggests a minor role of the OSM in glaucoma and is in contrast to many tear proteins with different abundances in glaucoma. The functional analysis of these tear proteins revealed an up-regulation of immune-related pathways in glaucoma patients compared to controls. In contrast to the OSM, this outcome suggests an important role of the tear proteome associated with the immune system in glaucoma.
While the innate immune system has long been linked to glaucoma through glial cells and oxidative stress, the findings support the role of the adaptive immune system [64]. Studies have shown that CD4+ T cells can facilitate RGC death [65], and an imbalance in Treg cells/T17 cells has been observed in experimental autoimmune optic neuritis, a condition sharing similarities with glaucoma [66]. Studies indicate a correlation between immune potency and resistance to RGC death, suggesting immune dysfunction as a potential condition for glaucoma development [67].
Despite being primarily associated with elevated IOP [68], the molecular mechanisms behind RGC death in glaucoma are complex and unclear [69]. Various factors contribute to RGC degeneration [70], including caspase activation [71,72], apoptosis [73], oxidative stress [74], ischemia [75], hypoxia, epigenetic changes [76], inflammatory cytokines [77], and neurotrophic factor deprivation [78]. Emerging evidence suggests the involvement of T cells in glaucoma pathology, with studies showing T cell infiltration in response to an elevated IOP and continued RGC degeneration [79].
In summary, subsequent investigations have identified autoantibodies targeting retinal and optic nerve proteins [70] in the serum [80], retina [81], and aqueous humor [82] of individuals with glaucoma. Additionally, research has indicated the existence of autoantibodies targeting heat shock proteins in individuals with glaucoma [83], which are molecular chaperones that can be up-regulated during stress [84] and potentially induce molecular mimicry, leading to the development of autoantibodies against host proteins [85]. Increased levels of specific heat shock proteins have been observed in glaucoma, with some potentially contributing to retinal ganglion cell death [86]. Administering heat shock proteins to rats through immunization has been demonstrated to cause glaucomatous damage in some studies [87]. Conversely, alternative research has indicated that the induction of heat shock proteins can confer neuroprotection for retinal ganglion cells [88,89,90]. In general, the involvement of autoantibodies and heat shock proteins in glaucoma is still not clearly understood.
3.4. Conclusions
In conclusion, the exploration of the OSM and tear proteome in glaucoma patients has provided insights into potential drivers of the disease and opened up promising avenues for targeted therapies. Previous studies proposed the use of animal models to examine changes in the microbiome pre- and post-glaucoma induction, aiming to determine whether these changes are a direct result of glaucoma or secondary to treatments involving preservative-laden eye drops. While existing studies have used mouse models to induce glaucoma [91,92,93,94], research specifically focusing on the OSM in these models has been limited.
Contrary to our initial expectations, our findings suggest that the OSM plays a minor role compared to the tear proteome in influencing the immunological processes associated with glaucoma. The significant enrichment of immune-related pathways observed in the tear proteome of glaucoma patients highlights the complexity of ocular surface homeostasis and its implications for disease pathogenesis. These results underscore the potential of the tear proteome as a rich source for innovative diagnostic and therapeutic strategies in glaucoma management.
By focusing on the tear proteome and its interaction with immune responses, personalized interventions tailored to individual patient profiles may offer new opportunities for improved clinical outcomes. Future research should continue to explore these interactions in greater depth, paving the way for the development of new treatment modalities that are more effective and less invasive than current options. This comprehensive approach is crucial for enhancing our understanding of glaucoma mechanisms and ultimately improving patient care.
4. Materials and Methods
4.1. Study Design and Recruitment
This study was approved by the Ethics Committee of the Canton of Bern (ClinicalTrials.gov: NCT04656197). The procedures followed the tenets of the Declaration of Helsinki and the International Ethical Guidelines for Biomedical Research involving Human Subjects (Council for International Organizations of Medical Sciences). After receiving oral and written information, all participants gave written informed consent before study enrollment. Participants (n = 32) were consecutively recruited from the Department of Ophthalmology of the University Hospital Bern (Inselspital), Switzerland. Patients with a history of recent (last 3 months) ocular surgery and systemic or topical antibiotics within the last 3 months were excluded. Participants fulfilling all of the following inclusion criteria were eligible for the study. The participants were willing to sign informed consent, were 50 years of age or older, and underwent glaucoma surgery within 24 h (glaucoma group). They were not eligible for the study if they were not willing or able to sign informed consent, were younger than 50 years, had a recent (3 months) history of use of systemic and/or topical antibiotics or ocular surgery, or a history of glaucoma (healthy control group).
4.2. Sample Collection
After verifying that no exclusion criteria were met and written informed consent was given, the tear fluid was collected by Schirmer’s type 1 tear test. A standard filter strip (Haag-Streit AG, Köniz, Bern, Switzerland) was inserted into the lower conjunctival bag of both eyes. After 5 min, the strip was removed and immediately processed for tear fluid extraction as described in [22]. The extracted tear fluid was stored at −80 °C until further analysis by nano-liquid chromatography–tandem mass spectrometry (nLC-MS/MS; Section 4.4).
A total of 32 conjunctival swabs using flocked nylon swabs (FLOQSwabs #518CS01, Copan, Brescia, Italy) from 16 patients and 16 controls were collected [38]. A local anesthetic (Tetracaine 1%, Théa, Schaffhausen, Switzerland) was applied. In controls, we performed a pooled swab from both eyes, while in the glaucoma group, we performed the conjunctival swab on the eye with glaucoma and a planned surgery within the next day. For negative controls, flocked nylon swabs with one drop of Tetracaine 1% (n = 2) were processed in parallel to conjunctival swabs.
4.3. Metagenomic DNA Sequencing and Analysis
Conjunctival swabs, as well as negative controls, were processed, and DNA was isolated on the same day using the QIAamp DNA Microbiome Kit (51704) from QIAGEN (Hilden, Germany). DNA was stored at −20 °C until further analysis. The DNA was then used to prepare sequencing libraries with the NEBNext Ultra II Preparation kit (SKU E7645L). The dual indexed libraries were sequenced on an Illumina NovaSeq 6000 sequencer for 150 paired-end cycles. Sequencing was performed at the Next-Generation Sequencing Platform of the University of Bern, Switzerland.
The resulting DNA sequences contain a Unique Molecular Identifier (UMI; [95]). The UMIs were first extracted using UMI-tools (v. 1.1.4). The reads were then quality-filtered using fastp (v. 0.20.1; [96]. Then, the reads were mapped to the human genome reference GRCh38 in order to remove the host DNA. Unmapped reads were extracted using SAMtools (v.1.10). The host-filtered reads were then mapped to the ChocoPhlAn reference database version mpa_vOct22_CHOCOPhlAnSGB_202212 using bowtie2 (v. 2.3.4.1; [97]). The resulting SAM alignment file was converted to the BAM format and then sorted and indexed using SAMtools (v. 1.10; [98]). The resulting file was then used to deduplicate the reads in the alignment based on the UMI and mapping location, using UMI-tools (v.1.1.4). The deduplicated BAM file was then used as input for the MetaPhlAn4 (v. 4.0.4; [99,100]) pipeline and the extracted deduplicated fastq files for the HUMAnN3 (v. 3.8) [101] pipeline.
In order to provide a global analysis of microbial abundances by PCA, the function adonis2 of the vegan R package and ggplot2 version 3.5.0 were applied. A PCA was performed using scaled values on the relative abundances of microbial species. A permutation multivariate analysis of variance (PERMANOVA) applying the R package vegan 2.6–4 was assessed with 1000 permutations to provide a p value for separation.
The Shannon index was calculated using vegan 2.6–4 and plotted by ggplot2 version 3.5.0. Associations of microbial and functional features of the microbiome with glaucoma or IOP-lowering eye drops, respectively, were identified by the MaAsLin2 R package, applying the default settings and using “groups” (glaucoma versus healthy controls, POAG versus PEXG patients, combination of four active ingredients of IOP-lowering eye drops including alpha-agonists, versus three active ingredients, versus no eye drops) as fixed and “age” as a random effect.
4.4. Tear Fluid Processing and Analysis [102,103,104,105,106,107,108,109,110,111]
For tear fluid collection, the tear fluid-soaked Schirmer strip was put into a 0.5 mL Protein LoBind tube (Eppendorf AG, Hamburg, Germany, 0030108094) punctured at the bottom with a cannula. This tube was placed into a larger 2 mL Protein LoBind tube (Eppendorf AG, Hamburg, Germany, 0030108132) and centrifuged at maximum speed (10,000 rpm) for 1 min. This procedure was initially described by Posa et al. [112] and allows the extraction of tear fluid. Tear fluid was stored at −80 °C until further analysis by nLC-MS/MS. A total of 2 µL human tear fluid was diluted with 8 µL 8M urea/100 mM Tris, reduced for 30 min at 37 °C with 0.1M DTT/100 mM Tris, alkylated for 30 min in darkness with 0.5M IAA/100 mM Tris at 37 °C, and quenched with DTT. The sample was filled up to 20 µL with 6× Laemmli buffer and heated to 95 °C for 5 min prior to loading on a 12.5% SDS-PAGE gel. The gel was run to about 1.5 cm length, stained with Coomassie blue, and each lane was cut into five bands equal in size. Gel bands were cut into small cubes, which were transferred to 1.5 mL polypropylene reaction vials and wetted with 100 µL ethanol for storage at 4 °C until digestion.
For nLC-MS/MS, the 36 samples were randomized in order to minimize the memory and matrix effect of the injections. Proteins were in-gel digested as described by Gunasekera et al. [113]. An aliquot of 5 µL from each digest was analyzed by an nLC-MS/MS instrument consisting of an EASY-nLC 1000 chromatograph coupled to a QExactive HF mass spectrometer (ThermoFisher Scientific). Peptides were trapped on an Acclaim C18 PepMap100 pre-column (5 μm, 100 Å, 300 μm × 5 mm, ThermoFisher Scientific, Reinach, Switzerland) and separated by backflush on a C18 column (3 μm, 100 Å, 75 μm × 15 cm, Nikkyo Technos, Tokyo, Japan) by applying a 40 min gradient of 5% acetonitrile to 40% in water, 0.1% formic acid, at a flow rate of 350 nL/min. Peptides of m/z 400–1400 were detected at a resolution of 60,000 (at m/z 250) with an automatic gain control (AGC) target of 1E06 and maximum ion injection time of 50 ms. A top-fifteen data-dependent method for precursor ion fragmentation was applied with the following settings: precursor ion isolation width of 1.6 m/z, resolution 15,000, AGC of 1 × 105, maximum ion time of 110 ms, charge inclusion of 2+ to 7+ ions, peptide match on, and dynamic exclusion for 20 s, respectively.
The mass spectrometry data were searched and quantified with FragPipe [fragpipe] version 20.0, using MSFragger [msfragger] version 3.8. The database used was the Human swissprot [uniprot] database (release April 2023) with isoforms and concatenated to common contaminants. The precursor and fragment tolerance were set, respectively, to 15 and 20 ppm. The search enzyme was set to trypsin, with a maximum number of allowed missed cleavages of 3. Carbamidomethylation on cysteine was set as a fixed modification; methionine oxidation, glutamine and asparagine deamidation, and protein N-terminal acetylation were given as variable modifications. The minimum of matched fragments was set to 5. Validation (Philosopher [philosopher] version 5.0.0) was performed with the Peptide Prophet option, and a protein false discovery rate of 0.01. Quantification was conducted with IonQuant [ionquant] version 1.9.8 (outcome named FragI) and Label Free Quantification (outcome named MaxLFQ). A match between runs was enabled, for a maximum of 10 top runs.
Proteins with less than 2 identified peptides were first removed from the list. After filtering off contaminants, proteins with less than 8 identifications in either control or patients, and less than 9 or 8 identifications in, respectively, the female or male groups, were discarded. MaxLFQ values were re-normalized by vsn [vsn] on this set of proteins. Missing values were imputed at protein level (iMaxLFQ) by drawing random values from a Gaussian distribution of width 0.3 × sample standard deviation and centered at the sample distribution mean minus 2.5 × sample standard deviation (protein level). Differential expression tests were performed with the moderated t test of the R limma package [ebayes], and the Benjamini and Hochberg [BH] method was further applied to correct for multiple testing. The criterion for statistically significant differential expression was that the maximum adjusted p-value for large fold changes was 0.05, and that this maximum decreased asymptotically to 0 as the log2 fold change of 1 was approached (with a curve parameter of 0.1 times the overall standard deviation; Figure 5). Proteins consistently significantly differentially expressed through 20 protein imputation cycles were subsequently flagged [transport].
Following the submission of a list containing all proteins identified in our samples with the fold change (iMaxLFQ log2FC patient–control, biological process, and iMaxLFQ EB patient–control, biological process) to STRING version 12.0 for analysis, we obtained a compilation of all biological processes analyzable in our samples. Subsequently, these processes were ranked based on the false discovery rate, which is a measure for the accuracy of the results. In order to provide a global analysis of protein abundances, the PCA was performed as described above.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/ijms25116257/s1.
Author Contributions
Conceptualization, D.C.Z.-B., J.D.U., and M.S.Z.; methodology, D.C.Z.-B., A.-C.U., M.K., and E.L.H.; formal analysis, L.S., A.-C.U., M.K., and E.L.H.; investigation, L.S.; writing—original draft preparation, L.S.; writing—review and editing, D.C.Z.-B., J.D.U., and M.S.Z.; visualization, L.S. and D.C.Z.-B.; supervision, D.C.Z.-B.; project administration, D.C.Z.-B.; funding acquisition, D.C.Z.-B. and M.S.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Foundation Bertarelli Catalyst Fund, EPFL (Ecole Polytechnique Fédérale de Lausanne), Lausanne, Switzerland (CF10000044—EPFL SCR0237812).
Institutional Review Board Statement
This study was reviewed and approved by the Ethics Committee of the Canton of Bern (ClinicalTrials.gov: NCT04656197). The procedures followed the tenets of the Declaration of Helsinki and the International Ethical Guidelines for Biomedical Research involving Human Subjects (Council for International Organizations of Medical Sciences).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The datasets supporting the conclusions of this article are available in the European Nucleotide Archive under the accession number PRJEB74248.
Acknowledgments
Mass spectrometry samples were processed and analyzed at the Proteomics and Mass Spectrometry Core Facility of the University of Bern.
Conflicts of Interest
The authors declare no conflicts of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results. Throughout the entirety of the work, ChatGPT 3.5 was utilized for structural assistance in writing the paper, as well as for linguistic and formal revisions.
References
- Weinreb, R.N.; Aung, T.; Medeiros, F.A. The pathophysiology and treatment of glaucoma: A review. JAMA 2014, 311, 1901–1911. [Google Scholar] [CrossRef]
- Agarwal, R.; Gupta, S.K.; Agarwal, P.; Saxena, R.; Agrawal, S.S. Current concepts in the pathophysiology of glaucoma. Indian J. Ophthalmol. 2009, 57, 257–266. [Google Scholar]
- Shon, K.; Wollstein, G.; Schuman, J.S.; Sung, K.R. Prediction of glaucomatous visual field progression: Pointwise analysis. Curr. Eye Res. 2014, 39, 705–710. [Google Scholar] [CrossRef] [PubMed]
- Tham, Y.C.; Li, X.; Wong, T.Y.; Quigley, H.A.; Aung, T.; Cheng, C.Y. Global prevalence of glaucoma and projections of glaucoma burden through 2040: A systematic review and meta-analysis. Ophthalmology 2014, 121, 2081–2090. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Hong, Y.; Fu, X.; Tan, H.; Chen, Y.; Wang, Y.; Chen, D. The role of the microbiota in glaucoma. Mol. Asp. Med. 2023, 94, 101221. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Wang, N.; Xiong, S.; Xia, X. The correlation between primary open-angle glaucoma (POAG) and gut microbiota: A pilot study towards predictive, preventive, and personalized medicine. EPMA J 2023, 14, 539–552. [Google Scholar] [CrossRef] [PubMed]
- Padhy, B.; Alone, D. Is pseudoexfoliation glaucoma a neurodegenerative disorder? J. Biosci. 2021, 46. [Google Scholar] [CrossRef]
- Sihota, R.; Angmo, D.; Ramaswamy, D.; Dada, T. Simplifying “target” intraocular pressure for different stages of primary open-angle glaucoma and primary angle-closure glaucoma. Indian J. Ophthalmol. 2018, 66, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Sambhara, D.; Aref, A.A. Glaucoma management: Relative value and place in therapy of available drug treatments. Ther. Adv. Chronic Dis. 2014, 5, 30–43. [Google Scholar] [CrossRef]
- Peirce, J.M.; Alviña, K.A.-O. The role of inflammation and the gut microbiome in depression and anxiety. J. Neurosci. Res. 2019, 97, 1223–1241. [Google Scholar] [CrossRef]
- Cresci, G.A.; Bawden, E. Gut Microbiome: What We Do and Don’t Know. Nutr. Clin. Pract. 2015, 30, 734–746. [Google Scholar] [CrossRef]
- Gancz, N.N.; Levinson, J.A.; Callaghan, B.L. Sex and gender as critical and distinct contributors to the human brain-gut-microbiome axis. Brain Res. Bull. 2023, 199, 110665. [Google Scholar] [CrossRef] [PubMed]
- Asadi, A.; Shadab Mehr, N.; Mohamadi, M.H.; Shokri, F.; Heidary, M.A.-O.; Sadeghifard, N.; Khoshnood, S. Obesity and gut-microbiota-brain axis: A narrative review. J. Clin. Lab. Anal. 2022, 36, e24420. [Google Scholar] [CrossRef] [PubMed]
- Trakman, G.A.-O.; Fehily, S.A.-O.; Basnayake, C.A.-O.X.; Hamilton, A.A.-O.; Russell, E.; Wilson-O’Brien, A.; Kamm, M.A.-O. Diet and gut microbiome in gastrointestinal disease. J. Gastroenterol. Hepatol. 2022, 37, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Simpson, C.A.; Diaz-Arteche, C.; Eliby, D.; Schwartz, O.S.; Simmons, J.G.; Cowan, C.S.M. The gut microbiota in anxiety and depression—A systematic review. Clin. Psychol. Rev. 2021, 83, 101943. [Google Scholar] [CrossRef] [PubMed]
- Krilis, M.; Fry, L.; Ngo, P.; Goldberg, I. The gut microbiome and primary open angle glaucoma: Evidence for a ‘gut-glaucoma’ axis? Eur. J. Ophthalmol. 2023, 11206721231219147. [Google Scholar] [CrossRef] [PubMed]
- Abraham, P.; Pratap, N. Dysbiosis in Irritable Bowel Syndrome. J. Assoc. Physicians India 2023, 79, 75–81. [Google Scholar]
- Li, J.; Wei, J.; Wang, J.; Xu, T.; Wu, B.; Yang, S.; Jing, S.; Wu, H.; Hao, H. Association between gut microbiota and spinal stenosis: A two-sample mendelian randomization study. Front. Immunol. 2024, 15, 1360132. [Google Scholar] [CrossRef] [PubMed]
- Pheiffer, C.A.-O.; Riedel, S.A.-O.; Dias, S.A.-O.; Adam, S.A.-O. Gestational Diabetes and the Gut Microbiota: Fibre and Polyphenol Supplementation as a Therapeutic Strategy. Microorganisms 2024, 12, 633. [Google Scholar] [CrossRef]
- Omar, W.E.W.; Singh, G.; McBain, A.J.; Cruickshank, F.; Radhakrishnan, H. Gut Microbiota Profiles in Myopes and Nonmyopes. Investig. Ophthalmol. Vis. Sci. 2024, 65, 2. [Google Scholar] [CrossRef]
- Yao, S.Q.; Yang, X.; Cen, L.P.; Tan, S. The Role of Gut Microbiota in Neuromyelitis Optica Spectrum Disorder. Int. J. Mol. Sci. 2024, 25, 3179. [Google Scholar] [CrossRef] [PubMed]
- Bron, A.J.; de Paiva, C.S.; Chauhan, S.K.; Bonini, S.; Gabison, E.E.; Jain, S.; Knop, E.; Markoulli, M.; Ogawa, Y.; Perez, V.; et al. TFOS DEWS II pathophysiology report. Ocul. Surf. 2017, 15, 438–510. [Google Scholar] [PubMed]
- Kalyana Chakravarthy, S.; Jayasudha, R.; Ranjith, K.; Dutta, A.; Pinna, N.K.; Mande, S.S.; Sharma, S.; Garg, P.; Murthy, S.I.; Shivaji, S. Alterations in the gut bacterial microbiome in fungal Keratitis patients. PLoS ONE 2018, 13, e0199640. [Google Scholar] [CrossRef] [PubMed]
- Petrillo, F.; Pignataro, D.; Lavano, M.A.; Santella, B.; Folliero, V.; Zannella, C.; Astarita, C.; Gagliano, C.; Franci, G.; Avitabile, T.; et al. Current Evidence on the Ocular Surface Microbiota and Related Diseases. Microorganisms 2020, 8, 1033. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Yang, B.; Li, W. Defining the normal core microbiome of conjunctival microbial communities. Clin. Microbiol. Infect. 2016, 22, 643.e7–643.e12. [Google Scholar] [CrossRef] [PubMed]
- Peter, V.G.; Morandi, S.C.; Herzog, E.L.; Zinkernagel, M.S.; Zysset-Burri, D.C. Investigating the Ocular Surface Microbiome: What Can It Tell Us? Clin. Ophthalmol. 2023, 17, 259–271. [Google Scholar] [CrossRef] [PubMed]
- Zysset-Burri, D.C.; Schlegel, I.; Lincke, J.B.; Jaggi, D.; Keller, I.; Heller, M.; Lagache, S.B.; Wolf, S.; Zinkernagel, M.S. Understanding the Interactions Between the Ocular Surface Microbiome and the Tear Proteome. Investig. Ophthalmol. Vis. Sci. 2021, 62, 8. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Hamady, M.; Fraser-Liggett, C.M.; Knight, R.; Gordon, J.I. The human microbiome project. Hum. Microbiome Project. Nat. 2007, 449, 804–810. [Google Scholar] [CrossRef] [PubMed]
- Schlegel, I.; De Gouyon Matignon de Pontourade, C.M.F.; Lincke, J.B.; Keller, I.; Zinkernagel, M.S.; Zysset-Burri, D.C. The Human Ocular Surface Microbiome and Its Associations with the Tear Proteome in Dry Eye Disease. Int. J. Mol. Sci. 2023, 24, 14091. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Fang, X.; Wang, X.; Lin, Y.A.; Wu, H.; Li, C. Effects of Sodium Hyaluronate Eye Drops with or without Preservatives on Ocular Surface Bacterial Microbiota. Front. Med. 2022, 9, 793565. [Google Scholar] [CrossRef]
- Hotta, F.; Eguchi, H.; Kuwahara, T.; Nakayama-Imaohji, H.; Shimomura, Y.; Kusaka, S. Disturbances in the ocular surface microbiome by perioperative antimicrobial eye drops. Front. Cell. Infect. Microbiol. 2023, 13, 1172345. [Google Scholar] [CrossRef] [PubMed]
- Labetoulle, M.; Baudouin, C.; Benitez Del Castillo, J.M.; Rolando, M.; Rescigno, M.; Messmer, E.M.; Aragona, P. How gut microbiota may impact ocular surface homeostasis and related disorders. Prog. Retin. Eye Res. 2024, 100, 101250. [Google Scholar] [CrossRef] [PubMed]
- Tezel, G. A proteomics view of the molecular mechanisms and biomarkers of glaucomatous neurodegeneration. Prog. Retin. Eye Res. 2013, 35, 18–43. [Google Scholar] [CrossRef] [PubMed]
- Wen, X.; Miao, L.; Deng, Y.; Bible, P.W.; Hu, X.; Zou, Y.; Liu, Y.; Guo, S.; Liang, J.; Chen, T.; et al. The Influence of Age and Sex on Ocular Surface Microbiota in Healthy Adults. Investig. Ophthalmol. Vis. Sci. 2017, 58, 6030–6037. [Google Scholar] [CrossRef] [PubMed]
- Katzka, W.; Dong, T.S.; Luu, K.; Lagishetty, V.; Sedighian, F.; Arias-Jayo, N.; Jacobs, J.P.; Hsu, H.Y. The Ocular Microbiome Is Altered by Sampling Modality and Age. Transl. Vis. Sci. Technol. 2021, 10, 24. [Google Scholar] [CrossRef] [PubMed]
- Nattinen, J.; Jylha, A.; Aapola, U.; Makinen, P.; Beuerman, R.; Pietila, J.; Vaajanen, A.; Uusitalo, H. Age-associated changes in human tear proteome. Clin. Proteom. 2019, 16, 11. [Google Scholar] [CrossRef] [PubMed]
- Mallick, H.; Rahnavard, A.; McIver, L.J.; Ma, S.; Zhang, Y.; Nguyen, L.H.; Tickle, T.L.; Weingart, G.; Ren, B.; Schwager, E.H.; et al. Multivariable association discovery in population-scale meta-omics studies. PLoS Comput. Biol. 2021, 17, e1009442. [Google Scholar] [CrossRef] [PubMed]
- Herzog, E.L.; Kreuzer, M.; Zinkernagel, M.S.; Zysset-Burri, D.C. Challenges and insights in the exploration of the low abundance human ocular surface microbiome. Front. Cell. Infect. Microbiol. 2023, 13, 1232147. [Google Scholar] [CrossRef] [PubMed]
- Gomes, J.A.P.; Frizon, L.; Demeda, V.F. Ocular Surface Microbiome in Health and Disease. Asia-Pac. J. Ophthalmol. 2020, 9, 505–511. [Google Scholar] [CrossRef] [PubMed]
- Grzybowski, A.; Brona, P.; Kim, S.J. Microbial flora and resistance in ophthalmology: A review. Graefes Arch. Clin. Exp. Ophthalmol. 2017, 255, 851–862. [Google Scholar] [CrossRef]
- Honda, R.; Toshida, H.; Suto, C.; Fujimaki, T.; Kimura, T.; Ohta, T.; Murakami, A. Effect of long-term treatment with eyedrops for glaucoma on conjunctival bacterial flora. Infect. Drug Resist. 2011, 4, 191–196. [Google Scholar] [CrossRef] [PubMed]
- Xue, W.; Li, J.J.; Zou, Y.; Zou, B.; Wei, L. Microbiota and Ocular Diseases. Front. Cell. Infect. Microbiol. 2021, 11, 759333. [Google Scholar] [CrossRef] [PubMed]
- Nayyar, A.; Gindina, S.; Barron, A.; Hu, Y.; Danias, J. Do epigenetic changes caused by commensal microbiota contribute to development of ocular disease? A review of evidence. Hum. Genom. 2020, 14, 11. [Google Scholar] [CrossRef] [PubMed]
- Guedes, G.; CTsai, J.; ALoewen, N. Glaucoma and aging. Glaucoma and aging. Curr. Aging Sci. 2011, 4, 110–117. [Google Scholar] [CrossRef] [PubMed]
- Pezzino, S.A.-O.; Sofia, M.; Greco, L.P.; Litrico, G.; Filippello, G.; Sarvà, I.; La Greca, G.; Latteri, S. Microbiome Dysbiosis: A Pathological Mechanism at the Intersection of Obesity and Glaucoma. Int. J. Mol. Sci. 2023, 24, 1166. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.Y.; Jung, K.I.; Park, H.Y.L.; Park, C.K. The effect of anxiety and depression on progression of glaucoma. Sci. Rep. 2021, 11, 1769. [Google Scholar] [CrossRef] [PubMed]
- Goyal, D.; Ali, S.A.; Singh, R.K. Emerging role of gut microbiota in modulation of neuroinflammation and neurodegeneration with emphasis on Alzheimer’s disease. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 2021, 106, 110112. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.J.; Gerasimidis, K.; Edwards, C.A.; Shaikh, M.G. Role of Gut Microbiota in the Aetiology of Obesity: Proposed Mechanisms and Review of the Literature. J. Obes. 2016, 2016, 7353642. [Google Scholar] [CrossRef] [PubMed]
- Fu, Y.; Wu, J.; Wang, D.; Li, T.; Shi, X.; Li, L.; Zhu, M.; Zhang, Z.; Yu, X.; Dai, Q. Metagenomic profiling of ocular surface microbiome changes in Demodex blepharitis patients. Front. Cell. Infect. Microbiol. 2022, 12, 922753. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zou, X.; Xue, W.; Zhang, P.; Wang, S.; Zou, H. Ocular Surface Microbiota in Diabetic Patients with Dry Eye Disease. Investig. Ophthalmol. Vis. Sci. 2021, 62, 13. [Google Scholar] [CrossRef]
- Liang, X.; Li, Y.; Xiong, K.; Chen, S.; Li, Z.; Zhang, Z.; Xia, Z.; Yi, G.; Fu, M. Demodex Infection Changes Ocular Surface Microbial Communities, in Which Meibomian Gland Dysfunction May Play a Role. Ophthalmol. Ther. 2021, 10, 601–617. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Ge, X.; Li, Y.; Zou, B.; Wen, X.; Chen, W.; Lu, L.; Zhang, M.; Zhang, X.; Li, C.; et al. Identification of an intraocular microbiota. Cell Discov. 2021, 7, 13. [Google Scholar] [CrossRef] [PubMed]
- Kittipibul, T.; Puangsricharern, V.; Chatsuwan, T. Comparison of the ocular microbiome between chronic Stevens-Johnson syndrome patients and healthy subjects. Sci. Rep. 2020, 10, 4353. [Google Scholar] [CrossRef] [PubMed]
- Cheleuitte-Nieves, C.; Gulvik, C.A.; Humrighouse, B.W.; Bell, M.E.; Villarma, A.; Westblade, L.F.; Lipman, N.S.; Fischetti, V.A.; McQuiston, J.R. Draft Reference Genome Sequence of Corynebacterium mastitidis 16-1433, Isolated from a Mouse. Genome Announc. 2018, 6. [Google Scholar] [CrossRef] [PubMed]
- St Leger, A.J.; Caspi, R.R. Visions of Eye Commensals: The Known and the Unknown about How the Microbiome Affects Eye Disease. BioEssays News Rev. Mol. Cell. Dev. Biol. 2018, 40, e1800046. [Google Scholar] [CrossRef] [PubMed]
- St Leger, A.J.; Desai, J.V.; Drummond, R.A.; Kugadas, A.; Almaghrabi, F.; Silver, P.; Raychaudhuri, K.; Gadjeva, M.; Iwakura, Y.; Lionakis, M.S.; et al. An Ocular Commensal Protects against Corneal Infection by Driving an Interleukin-17 Response from Mucosal gammadelta T Cells. Immunity 2017, 47, 148–158.e5. [Google Scholar] [CrossRef] [PubMed]
- Mandal, A.; Varghese, M.V.; James, J. Chapter 22—Clinical significance of phospholipase A2 in glaucoma. In Phospholipases in Physiology and Pathology; Chakraborti, S., Ed.; Academic Press: Cambridge, MA, USA, 2023; pp. 331–341. [Google Scholar] [CrossRef]
- Zhou, Y.; Sidhu, G.S.; Whitlock, J.A.; Abdelmalik, B.; Mayer, Z.; Li, Y.; Wang, G.P.; Steigleman, W.A. Effects of Carboxymethylcellulose Artificial Tears on Ocular Surface Microbiome Diversity and Composition, A Randomized Controlled Trial. Transl. Vis. Sci. Technol. 2023, 12, 5. [Google Scholar] [CrossRef] [PubMed]
- Ohtani, S.; Shimizu, K.; Nejima, R.; Kagaya, F.; Aihara, M.; Iwasaki, T.; Shoji, N.; Miyata, K. Conjunctival Bacteria Flora of Glaucoma Patients During Long-Term Administration of Prostaglandin Analog Drops. Investig. Ophthalmol. Vis. Sci. 2017, 58, 3991–3996. [Google Scholar] [CrossRef] [PubMed]
- Priluck, A.; Ramulu, P.; Dosto, N.; Quigley, H.; Abraham, A. Validation of 16S rRNA Gene Sequencing of the Periocular Microbiome and Lack of Alteration by Topical Eyedrops. Transl. Vis. Sci. Technol. 2023, 12, 32. [Google Scholar] [CrossRef]
- Ekici Gok, Z.; Gunduz, A.; Bozgül, P.G. Evaluation of the effects of mono or combined use of topical antiglaucomatous drops on conjunctival flora and antibiotic susceptibility. Eur. J. Ophthalmol. 2023, 11206721231219275. [Google Scholar] [CrossRef]
- de Kaspar, H.M.; Kreidl, K.O.; Singh, K.; Ta, C.N. Comparison of preoperative conjunctival bacterial flora in patients undergoing glaucoma or cataract surgery. J. Glaucoma 2004, 13, 507–509. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.J.; Somohano, K.; Zemsky, C.; Uhlemann, A.C.; Liebmann, J.; Cioffi, G.A.; Al-Aswad, L.A.; Lynch, S.V.; Winn, B.J. Topical Glaucoma Therapy Is Associated with Alterations of the Ocular Surface Microbiome. Investig. Ophthalmol. Vis. Sci. 2022, 63, 32. [Google Scholar] [CrossRef]
- Jiang, S.; Kametani, M.; Chen, D.F. Adaptive Immunity: New Aspects of Pathogenesis Underlying Neurodegeneration in Glaucoma and Optic Neuropathy. Front. Immunol. 2020, 11, 65. [Google Scholar] [CrossRef] [PubMed]
- He, C.; Xiu, W.; Chen, Q.; Peng, K.; Zhu, X.; Wang, Z.; Xu, X.; Chen, Y.; Zhang, G.; Fu, J.; et al. Gut-licensed β7+ CD4+ T cells contribute to progressive retinal ganglion cell damage in glaucoma. Sci. Transl. Med. 2023, 15, eadg1656. [Google Scholar] [CrossRef] [PubMed]
- Cong, H.; Jiang, H.; Peng, J.; Cui, S.; Liu, L.; Wang, J.; Zhang, X. Change of Th17 Lymphocytes and Treg/Th17 in Typical and Atypical Optic Neuritis. PLoS ONE 2016, 11, e0146270. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Li, Z.; van Rooijen, N.; Wang, N.; Pang, C.P.; Cui, Q. Different responses of macrophages in retinal ganglion cell survival after acute ocular hypertension in rats with different autoimmune backgrounds. Exp. Eye Res. 2007, 85, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Mick, A.B. A Few Steps Toward Improving Glaucoma Diagnostic Accuracy and Understanding Intraocular Pressure. Optom. Vis. Sci. 2018, 95, 86–87. [Google Scholar] [CrossRef] [PubMed]
- Zukerman, R.; Harris, A.; Oddone, F.; Siesky, B.; Verticchio Vercellin, A.; Ciulla, T.A. Glaucoma Heritability: Molecular Mechanisms of Disease. Genes 2021, 12, 1135. [Google Scholar] [CrossRef] [PubMed]
- DeMaio, A.; Mehrotra, S.; Sambamurti, K.; Husain, S. The role of the adaptive immune system and T cell dysfunction in neurodegenerative diseases. J. Neuroinflammation 2022, 19, 251. [Google Scholar] [CrossRef]
- Kermer, P.; Klocker, N.; Labes, M.; Thomsen, S.; Srinivasan, A.; Bahr, M. Activation of caspase-3 in axotomized rat retinal ganglion cells in vivo. FEBS Lett. 1999, 453, 361–364. [Google Scholar] [CrossRef]
- Husain, S.; Abdul, Y.; Crosson, C.E. Preservation of retina ganglion cell function by morphine in a chronic ocular-hypertensive rat model. Investig. Ophthalmol. Vis. Sci. 2012, 53, 4289–4298. [Google Scholar] [CrossRef] [PubMed]
- Almasieh, M.; Wilson, A.M.; Morquette, B.; Vargas, J.L.C.; Di Polo, A. The molecular basis of retinal ganglion cell death in glaucoma. Prog. Retin. Eye Res. 2012, 31, 152–181. [Google Scholar] [CrossRef] [PubMed]
- Himori, N.; Yamamoto, K.; Maruyama, K.; Ryu, M.; Taguchi, K.; Yamamoto, M.; Nakazawa, T. Critical role of Nrf2 in oxidative stress-induced retinal ganglion cell death. J. Neurochem. 2013, 127, 669–680. [Google Scholar] [CrossRef] [PubMed]
- Shen, W.C.; Huang, B.Q.; Yang, J. Regulatory mechanisms of retinal ganglion cell death in normal tension glaucoma and potential therapies. Neural Regen. Res. 2023, 18, 87–93. [Google Scholar] [PubMed]
- Coleman-Belin, J.A.-O.; Harris, A.; Chen, B.; Zhou, J.; Ciulla, T.A.-O.; Verticchio, A.; Antman, G.; Chang, M.; Siesky, B. Aging Effects on Optic Nerve Neurodegeneration. Int. J. Mol. Sci. 2023, 24, 2573. [Google Scholar] [CrossRef] [PubMed]
- Sivakumar, V.; Foulds, W.S.; Luu, C.D.; Ling, E.A.; Kaur, C. Retinal ganglion cell death is induced by microglia derived pro-inflammatory cytokines in the hypoxic neonatal retina. J. Pathol. 2011, 224, 245–260. [Google Scholar] [CrossRef] [PubMed]
- Claes, M.; De Groef, L.A.-O.; Moons, L.A.-O. Target-Derived Neurotrophic Factor Deprivation Puts Retinal Ganglion Cells on Death Row: Cold Hard Evidence and Caveats. Int. J. Mol. Sci. 2019, 20, 4314. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Hou, X.; Ni, Y.; Du, W.; Han, H.; Yu, Y.; Shi, G. The Imbalance of FOXP3/GATA3 in Regulatory T Cells from the Peripheral Blood of Asthmatic Patients. J. Immunol. Res. 2018, 2018, 3096183. [Google Scholar] [CrossRef] [PubMed]
- Joachim, S.C.; Grus, F.H.; Pfeiffer, N. Analysis of autoantibody repertoires in sera of patients with glaucoma. Eur. J. Ophthalmol. 2003, 13, 752–758. [Google Scholar] [CrossRef]
- Dutta Majumder, P.; Marchese, A.; Pichi, F.; Garg, I.; Agarwal, A. An update on autoimmune retinopathy. Indian J. Ophthalmol. 2020, 68, 1829–1837. [Google Scholar] [CrossRef]
- Joachim, S.C.; Bruns, K.; Lackner, K.J.; Pfeiffer, N.; Grus, F.H. Antibodies to α B-Crystallin, Vimentin, and Heat Shock Protein 70 in Aqueous Humor of Patients with Normal Tension Glaucoma and IgG Antibody Patterns against Retinal Antigen in Aqueous Humor. Curr. Eye Res. 2007, 32, 501–509. [Google Scholar] [CrossRef] [PubMed]
- Tezel, G.; Seigel, G.M.; Wax, M.B. Autoantibodies to small heat shock proteins in glaucoma. Investig. Ophthalmol. Vis. Sci. 1998, 39, 2277–2287. [Google Scholar]
- Loones, M.T.; Chang, Y.; Morange, M. The distribution of heat shock proteins in the nervous system of the unstressed mouse embryo suggests a role in neuronal and non-neuronal differentiation. Cell Stress Chaperones 2000, 5, 291. [Google Scholar] [CrossRef] [PubMed]
- Grus, F.H.; Joachim, S.C.; Wuenschig, D.; Rieck, J.; Pfeiffer, N. Autoimmunity and glaucoma. J. Glaucoma 2008, 17, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Wakefield, D.; Wildner, G. Is glaucoma an autoimmune disease? Clin. Transl. Immunol. 2020, 9, e1180. [Google Scholar] [CrossRef] [PubMed]
- Laspas, P.; Gramlich, O.W.; Muller, H.D.; Cuny, C.S.; Gottschling, P.F.; Pfeiffer, N.; Dick, H.B.; Joachim, S.C.; Grus, F.H. Autoreactive antibodies and loss of retinal ganglion cells in rats induced by immunization with ocular antigens. Investig. Ophthalmol. Vis. Sci. 2011, 52, 8835–8848. [Google Scholar] [CrossRef] [PubMed]
- Qing, G.; Duan, X.; Jiang, Y. Heat shock protein 72 protects retinal ganglion cells in rat model of acute glaucoma. Yan Ke Xue Bao 2005, 21, 163–168. [Google Scholar] [PubMed]
- Ishii, Y.; Kwong, J.M.; Caprioli, J. Retinal ganglion cell protection with geranylgeranylacetone, a heat shock protein inducer, in a rat glaucoma model. Investig. Ophthalmol. Vis. Sci. 2003, 44, 1982–1992. [Google Scholar] [CrossRef]
- Park, K.H.; Cozier, F.; Ong, O.C.; Caprioli, J. Induction of heat shock protein 72 protects retinal ganglion cells in a rat glaucoma model. Investig. Ophthalmol. Vis. Sci. 2001, 42, 1522–1530. [Google Scholar]
- Harada, C.; Kimura, A.; Guo, X.; Namekata, K.; Harada, T. Recent advances in genetically modified animal models of glaucoma and their roles in drug repositioning. Br. J. Ophthalmol. 2019, 103, 161–166. [Google Scholar] [CrossRef]
- Ishikawa, M.; Yoshitomi, T.; Zorumski, C.F.; Izumi, Y. Experimentally Induced Mammalian Models of Glaucoma. Biomed. Res. Int. 2015, 2015, 281214. [Google Scholar] [CrossRef]
- Yang, Q.; Cho, K.S.; Chen, H.; Yu, D.; Wang, W.H.; Luo, G.; Pang, I.H.; Guo, W.; Chen, D.F. Microbead-induced ocular hypertensive mouse model for screening and testing of aqueous production suppressants for glaucoma. Investig. Ophthalmol. Vis. Sci. 2012, 53, 3733–3741. [Google Scholar] [CrossRef]
- Hao, H.J.; Li, Y.H.; Yu, B.; Liu, X.; Zhang, Y.; Xing, X.L. Neuroprotective effects of acteoside in a glaucoma mouse model by targeting Serta domain-containing protein 4. Int. J. Ophthalmol. 2024, 17, 625–637. [Google Scholar] [CrossRef] [PubMed]
- Smith, T.; Heger, A.; Sudbery, I. UMI-tools: Modeling sequencing errors in Unique Molecular Identifiers to improve quantification accuracy. Genome Res. 2017, 27, 491–499. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef] [PubMed]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Genome Project Data Processing, S. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef] [PubMed]
- Blanco-Miguez, A.; Beghini, F.; Cumbo, F.; McIver, L.J.; Thompson, K.N.; Zolfo, M.; Manghi, P.; Dubois, L.; Huang, K.D.; Thomas, A.M.; et al. Extending and improving metagenomic taxonomic profiling with uncharacterized species with MetaPhlAn 4. Nat. Biotechnol. 2023, 41, 1633–1644. [Google Scholar] [CrossRef]
- Truong, D.T.; Tett, A.; Pasolli, E.; Huttenhower, C.; Segata, N. Microbial strain-level population structure and genetic diversity from metagenomes. Genome Res. 2017, 27, 626–638. [Google Scholar] [CrossRef]
- Beghini, F.; McIver, L.J.; Blanco-Míguez, A.; Dubois, L.; Asnicar, F.; Maharjan, S.; Mailyan, A.; Manghi, P.; Scholz, M.; Thomas, A.M.; et al. Integrating taxonomic, functional, and strain-level profiling of diverse microbial communities with bioBakery 3. eLife 2021, 10, e65088. [Google Scholar] [CrossRef]
- UniProt Consortium. UniProt: A worldwide hub of protein knowledge. Nucleic Acids Res. 2019, 47, D506–D515. [Google Scholar] [CrossRef]
- Yu, F.; Haynes, S.E.; Teo, G.C.; Avtonomov, D.M.; Polasky, D.A.; Nesvizhskii, A.I. Fast Quantitative Analysis of timsTOF PASEF Data with MSFragger and IonQuant. Mol. Cell. Proteom. 2020, 19, 1575–1585. [Google Scholar] [CrossRef] [PubMed]
- Kong, A.T.; Leprevost, F.A.-O.; Avtonomov, D.M.; Mellacheruvu, D.A.-O.; Nesvizhskii, A.A.-O. MSFragger: Ultrafast and comprehensive peptide identification in mass spectrometry-based proteomics. Nat. Methods 2017, 14, 513–520. [Google Scholar] [CrossRef] [PubMed]
- da Veiga Leprevost, F.; Haynes, S.E.; Avtonomov, D.M.; Chang, H.Y.; Shanmugam, A.K.; Mellacheruvu, D.; Kong, A.T.; Nesvizhskii, A.I. Philosopher: A versatile toolkit for shotgun proteomics data analysis. Nat. Methods 2020, 17, 869–870. [Google Scholar] [CrossRef]
- Yu, F.; Haynes, S.E.; Nesvizhskii, A.I. IonQuant Enables Accurate and Sensitive Label-Free Quantification with FDR-Controlled Match-Between-Runs. Mol. Cell. Proteom. 2021, 20, 100077. [Google Scholar] [CrossRef]
- Silva, J.C.; Gorenstein, M.V.; Li, G.Z.; Vissers, J.P.; Geromanos, S.J. Absolute quantification of proteins by LCMSE: A virtue of parallel MS acquisition. Mol. Cell. Proteom. 2006, 5, 144–156. [Google Scholar] [CrossRef]
- Huber, W.; Von Heydebreck, A.; Sültmann, H.; Poustka, A.; Vingron, M. Variance stabilization applied to microarray data calibration and to the quantification of differential expression. Bioinformatics 2002, 18, S96–S104. [Google Scholar] [CrossRef]
- Kammers, K.; Cole, R.N.; Tiengwe, C.; Ruczinski, I. Detecting Significant Changes in Protein Abundance. EuPA Open Proteom. 2015, 7, 11–19. [Google Scholar] [CrossRef] [PubMed]
- Uldry, A.A.-O.; Maciel-Dominguez, A.; Jornod, M.; Buchs, N.; Braga-Lagache, S.; Brodard, J.A.-O.; Jankovic, J.; Bonadies, N.A.-O.; Heller, M.A.-O. Effect of Sample Transportation on the Proteome of Human Circulating Blood Extracellular Vesicles. Int. J. Mol. Sci. 2022, 23, 4515. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B (Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Posa, A.; Bräuer, L.; Schicht, M.; Garreis, F.; Beileke, S.; Paulsen, F. Schirmer strip vs. capillary tube method: Non-invasive methods of obtaining proteins from tear fluid. Ann. Anat.-Anat. Anz. 2013, 195, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Gunasekera, K.; Wüthrich, D.; Braga-Lagache, S.; Heller, M.; Ochsenreiter, T. Proteome remodelling during development from blood to insect-form Trypanosoma brucei quantified by SILAC and mass spectrometry. BMC Genom. 2012, 13, 556. [Google Scholar] [CrossRef] [PubMed]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).