
The Role of MicroRNAs in the Pathophysiology of Osteoarthritis

 , , ,
, , ,
Abstract
1. Introduction
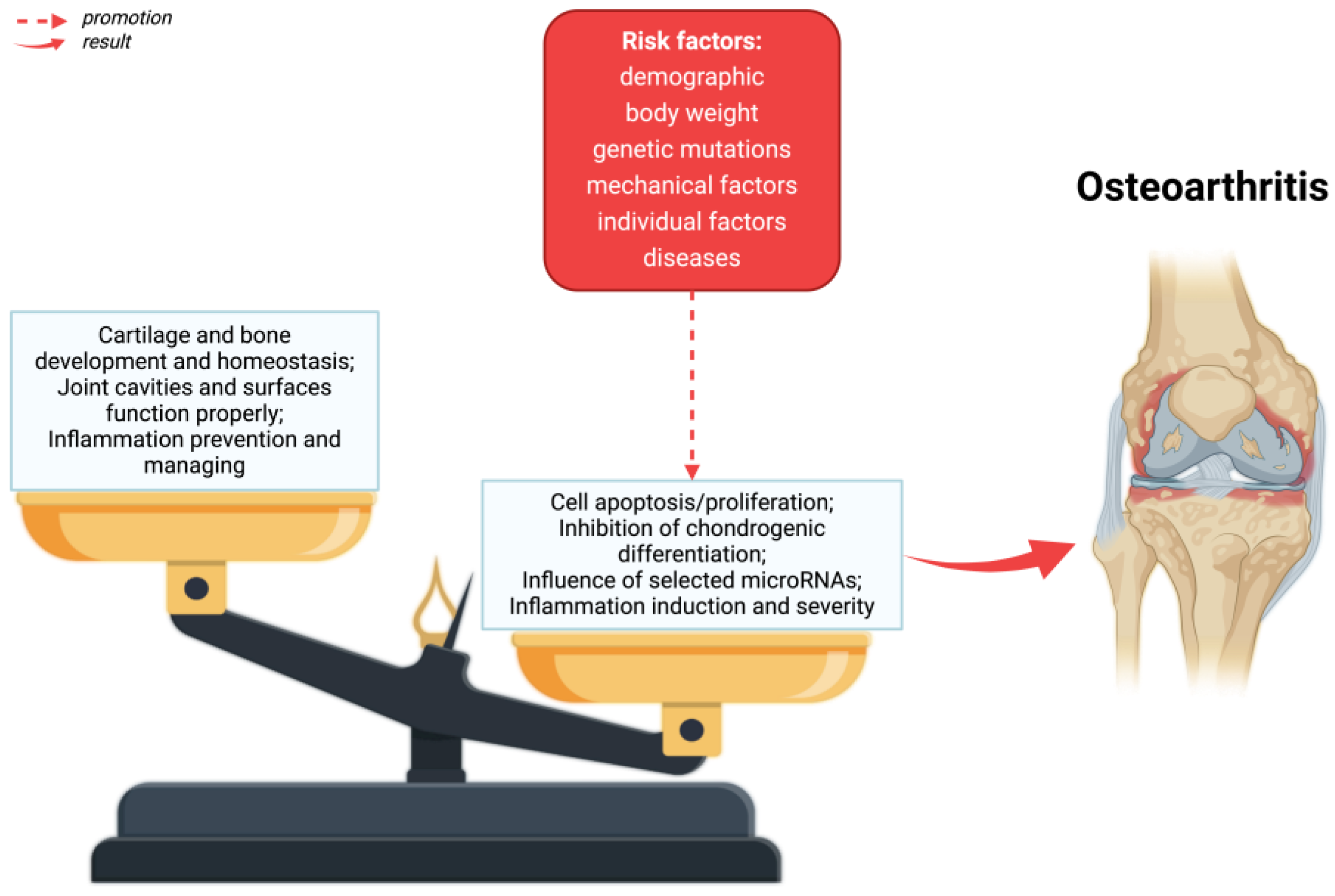
1.1. Osteoarthritis
1.2. Risk Factors
1.3. Clinical Relevance
2. Pathophysiology of Osteoarthritis
3. The Role of miRNAs in the Pathogenesis of Osteoarthritis

{kind=link}
{kind=link}
| MicroRNA | Level in OA | Potentially Action | Potential Target | Role in OA Pathogenesis | References |
|---|---|---|---|---|---|
| miR-9-5p | ↓ 1 | intensifying proliferation and suppressing chondrocyte apoptosis | MMP-13, PRTG | − 3 | [124] |
| miR-10a-5p | ↑ 2 | inhibiting chondrocyte proliferation, promoting chondrocyte apoptosis, and promoting cartilage matrix degradation | HOXA3 | + 4 | [127] |
| miR-22 | ↑ | the activation of metalloproteinases and aggrecanases and downregulation of cartilage structural proteins, cartilage degradation | PPARα, BMP-7 | + | [128] |
| miR-27b | ↑ | the fibrosis of the synovial membrane, influence on inflammatory processes, cartilage metabolism, and apoptosis of cartilage cells | MMP-13, COL1A1, α-SMA2, ADAMTS8, and CBFB | + | [144,145,146] |
| miR-34a-5p | ↑ | cell cycle arrest, promoting apoptosis, senescence, and proliferation | COL2A1, ACAN, ATG5, MMP13, ADAMTS5, IL-1β, and COL10A1 | + | [131] |
| miR-127-5p | ↓ | increasing the synthesis of cartilage extracellular matrix (ECM) | Osteoponin and MMP-13 | − | [147,148] |
| miR-128a | ↑ | impaired chondrocyte autophagy, the suppression of extracellular matrix deposition | Atg12, Bax, Bcl2, and cleaved caspase-3 | + | [137,138] |
| miR-138-5p | ↑ | the degradation of cartilage extracellular matrix (ECM) | FOXC1 and increase in IL-1β | + | [124] |
| miR-140 | ↓ | promoting chondrocyte differentiation | ADAMTS5 and AGGRECAN | − | [132] |
| miR-140-3p | ↓ | increase in the viability and migration capacity of chondrocytes | increase: SOX-9, COL2, ACAN, RUNX2, and SCX, decrease: COL1, COL6, COMP, TNC, and FMOD | − | [149] |
| miR-140-5p | ↓ | inhibits inflammation in the joint cavity, inhibits the progression of OA, promotes chondrogenesis, inhibits chondrocyte apoptosis, inhibits chondrocyte hypertrophy | IGFBP-5, IL-1β, IL-6, Syndecan-4, ADAMTS5, MMP-13, SMAD3, HMGB1, RALA, FUT1, HDAC4, and SMAD1 | − | [105,109,111,150,151,152,153,154,155,156] |
| mi-146 | ↑ | promoting the inflammatory response in the joint | TRAF6 and IRAQ1, | + | [157,158] |
| miR-146a-5p | ↑ | cartilage degradation, synovitis, neoangiogenesis, and osteoclastogenesis | TNF α, IL-1β, TRAF6 and IRAK1 genes, and MMP-13 | + | [114,124,159] |
| miR-149 | ↓ | promoting the synthesis of connective protein and proteoglycan, suppressing the inflammatory process | TNFα, IL1β, IL6, VCAM-1, and TAK1 | − | [134,135] |
| miR-210 | ↓ | promoting osteoblastic differentiation, anti-apoptotic effect, anti-inflammatory effect | AcvR1b and DR6 | − | [139,140,141,160] |
| miR-335-5p | ↑ | osteogenic and adipogenic differentiation, promoting ECM degradation | Wnt signaling pathway, IFNγ, HBP1, ACAN, MMP13, collagen X, and collagen II | + | [124,142] |
| miR-485-5p | ↑ | inhibiting the differentiation of BMSCs into chondroblasts and promoting the expression of inflammatory factors | SOX9 | + | [143] |
4. Potential Therapeutic Targets
5. Discussion
6. Conclusions
Funding
Conflicts of Interest
Abbreviations
| ACAN | Aggrecan |
| ACR | American College of Rheumatology |
| ADAMTS5 | A Disintegrin and Metalloproteinase with Thrombospondin Motifs 5 |
| ADAMTS8 | A Disintegrin and Metalloproteinase with Thrombospondin Motifs 8 |
| Atg12 | Autophagy Related 12 |
| ATG5 | Autophagy Related 5 |
| Bax | BCL2 Associated X, Apoptosis Regulator |
| Bcl2 | B-Cell Lymphoma 2 |
| BMP-7 | Bone Morphogenetic Protein 7 |
| CBFB | Core-Binding Factor Subunit Beta |
| COL10A1 | Collagen Type X Alpha 1 Chain |
| COL1A1 | Collagen Type I Alpha 1 Chain |
| COL2 | Collagen Type II |
| COL2A1 | Collagen Type II Alpha 1 Chain |
| COL6 | Collagen Type VI |
| COMP | Cartilage Oligomeric Matrix Protein |
| DR6 | Death Receptor 6 |
| ECM | Extracellular matrix |
| FMOD | Fibromodulin |
| FOXC1 | Forkhead Box C1 |
| FUT1 | Fucosyltransferase 1 |
| HDAC4 | Histone Deacetylase 4 |
| HMGB1 | High Mobility Group Box 1 |
| HOXA3 | Homeobox A3 |
| HBP1 | HMG-Box Transcription Factor 1 |
| IFNγ | Interferon Gamma |
| IGFBP-5 | Insulin-Like Growth Factor Binding Protein 5 |
| IL | interleukins |
| IRAK1 | Interleukin-1 Receptor-Associated Kinase 1 |
| MiRNA, MiR | microRNA |
| MMP-13 | Matrix Metallopeptidase 13 |
| NF-κB | Nuclear factor kappa B |
| OA | osteoarthritis |
| PARP | Poly (ADP-Ribose) Polymerase |
| PPARα/γ | Peroxisome Proliferator-Activated Receptor α/γ |
| PRTG | Protogenin |
| RALA | Ras-Like Proto-Oncogene A |
| RUNX2 | Runt-Related Transcription Factor 2 |
| SCX | Scleraxis |
| SMAD1 | SMAD Family Member 1 |
| SMAD3 | SMAD Family Member 3 |
| SOX9 | SRY-Box Transcription Factor 9 |
| TAK1 | TGF-Beta Activated Kinase 1 |
| TNC | Tenascin C |
| TNFα | Tumor Necrosis Factor Alpha |
| TRAF6 | TNF Receptor Associated Factor 6 |
| VCAM-1 | Vascular Cell Adhesion Molecule 1 |
| α-SMA2 | α Smooth Muscle Actin 2 |
References
- Yunus, M.H.M.; Nordin, A.; Kamal, H. Pathophysiological Perspective of Osteoarthritis. Medicina 2020, 56, 614. [Google Scholar] [CrossRef] [PubMed]
- Xia, B.; Di, C.; Zhang, J.; Hu, S.; Jin, H.; Tong, P. Osteoarthritis Pathogenesis: A Review of Molecular Mechanisms. Calcif. Tissue Int. 2014, 95, 495–505. [Google Scholar] [CrossRef] [PubMed]
- Hunter, D.J.; March, L.; Chew, M. Osteoarthritis in 2020 and beyond: A Lancet Commission. Lancet 2020, 396, 1711–1712. [Google Scholar] [CrossRef] [PubMed]
- Long, H.; Liu, Q.; Yin, H.; Wang, K.; Diao, N.; Zhang, Y.; Lin, J.; Guo, A. Prevalence Trends of Site-Specific Osteoarthritis from 1990 to 2019: Findings from the Global Burden of Disease Study 2019. Arthritis Rheumatol. 2022, 74, 1172–1183. [Google Scholar] [CrossRef] [PubMed]
- Mobasheri, A.; Thudium, C.S.; Bay-Jensen, A.C.; Maleitzke, T.; Geissler, S.; Duda, G.N.; Winkler, T. Biomarkers for Osteoarthritis: Current Status and Future Prospects. Best Pract. Res. Clin. Rheumatol. 2023, 37, 101852. [Google Scholar] [CrossRef] [PubMed]
- Cui, A.; Li, H.; Wang, D.; Zhong, J.; Chen, Y.; Lu, H. Global, Regional Prevalence, Incidence and Risk Factors of Knee Osteoarthritis in Population-Based Studies. eClinicalMedicine 2020, 29–30, 100587. [Google Scholar] [CrossRef] [PubMed]
- Hunter, D.J.; Felson, D.T. Osteoarthritis. Br. Med. J. 2006, 332, 639. [Google Scholar] [CrossRef] [PubMed]
- Glyn-Jones, S.; Palmer, A.J.R.; Agricola, R.; Price, A.J.; Vincent, T.L.; Weinans, H.; Carr, A.J. Osteoarthritis. Lancet 2015, 386, 376–387. [Google Scholar] [CrossRef] [PubMed]
- Moseng, T.; Vliet Vlieland, T.P.M.; Battista, S.; Beckwée, D.; Boyadzhieva, V.; Conaghan, P.G.; Costa, D.; Doherty, M.; Finney, A.G.; Georgiev, T.; et al. EULAR Recommendations for the Non-Pharmacological Core Management of Hip and Knee Osteoarthritis: 2023 Update. Ann. Rheum. Dis. 2024, 83, 730–740. [Google Scholar] [CrossRef]
- Yao, Q.; Wu, X.; Tao, C.; Gong, W.; Chen, M.; Qu, M.; Zhong, Y.; He, T.; Chen, S.; Xiao, G. Osteoarthritis: Pathogenic Signaling Pathways and Therapeutic Targets. Signal Transduct. Target. Ther. 2023, 8, 56. [Google Scholar] [CrossRef]
- Little, C.B.; Hunter, D.J. Post-Traumatic Osteoarthritis: From Mouse Models to Clinical Trials. Nat. Rev. Rheumatol. 2013, 9, 485–497. [Google Scholar] [CrossRef] [PubMed]
- Hall, M.; van der Esch, M.; Hinman, R.S.; Peat, G.; de Zwart, A.; Quicke, J.G.; Runhaar, J.; Knoop, J.; van der Leeden, M.; de Rooij, M.; et al. How Does Hip Osteoarthritis Differ from Knee Osteoarthritis? Osteoarthr. Cartil. 2022, 30, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Zhang, Z.; Chen, Y.; Zhang, Y.; Xing, D.; Zhao, L.; Lin, J.; Mei, Y.; Lin, H.-Y.; Zheng, Y.; et al. Development and Formulation of the Classification Criteria for Osteoarthritis. Ann. Transl. Med. 2020, 8, 1068. [Google Scholar] [CrossRef] [PubMed]
- Wei, G.; Lu, K.; Umar, M.; Zhu, Z.; Lu, W.W.; Speakman, J.R.; Chen, Y.; Tong, L.; Chen, D. Risk of Metabolic Abnormalities in Osteoarthritis: A New Perspective to Understand Its Pathological Mechanisms. Bone Res. 2023, 11, 63. [Google Scholar] [CrossRef] [PubMed]
- Lee, K.M.; Chung, C.Y.; Sung, K.H.; Lee, S.Y.; Won, S.H.; Kim, T.G.; Choi, Y.; Kwon, S.S.; Kim, Y.H.; Park, M.S. Risk Factors for Osteoarthritis and Contributing Factors to Current Arthritic Pain in South Korean Older Adults. Yonsei Med. J. 2015, 56, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Sharma, L. Osteoarthritis of the Knee. N. Engl. J. Med. 2021, 384, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Krakowski, P.; Karpiński, R.; Maciejewski, R.; Jonak, J.; Jurkiewicz, A. Short-Term Effects of Arthroscopic Microfracturation of Knee Chondral Defects in Osteoarthritis. Appl. Sci. 2020, 10, 8312. [Google Scholar] [CrossRef]
- Karpiński, R. Knee Joint Osteoarthritis Diagnosis Based on Selected Acoustic Signal Discriminants Using Machine Learning. Appl. Comput. Sci. 2022, 18, 71–85. [Google Scholar] [CrossRef]
- Lee, R.; Kean, W.F. Obesity and Knee Osteoarthritis. Inflammopharmacology 2012, 20, 53–58. [Google Scholar] [CrossRef]
- Michael, J.W.-P.; Schlüter-Brust, K.U.; Eysel, P. The Epidemiology, Etiology, Diagnosis, and Treatment of Osteoarthritis of the Knee. Dtsch. Arztebl. Int. 2010, 107, 152–162. [Google Scholar] [CrossRef]
- Karpiński, R.; Krakowski, P.; Jonak, J.; Machrowska, A.; Maciejewski, M. Comparison of Selected Classification Methods Based on Machine Learning as a Diagnostic Tool for Knee Joint Cartilage Damage Based on Generated Vibroacoustic Processes. Appl. Comput. Sci. 2023, 19, 136–150. [Google Scholar] [CrossRef]
- Felson, D.T. Clinical Practice. Osteoarthritis of the Knee. N. Engl. J. Med. 2006, 354, 841–848. [Google Scholar] [CrossRef] [PubMed]
- Abramoff, B.; Caldera, F.E. Osteoarthritis: Pathology, Diagnosis, and Treatment Options. Med. Clin. N. Am. 2020, 104, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann-Górska, I.; Szczepański, L. Osteoarthritis. In Interna Szczeklika 2023; Szczeklik, A., Gajewski, P., Eds.; Medycyna Praktyczna: Kraków, Poland, 2023; pp. 2162–2170. [Google Scholar]
- Palazzo, C.; Nguyen, C.; Lefevre-Colau, M.M.; Rannou, F.; Poiraudeau, S. Risk Factors and Burden of Osteoarthritis. Ann. Phys. Rehabil. Med. 2016, 59, 134–138. [Google Scholar] [CrossRef] [PubMed]
- Musumeci, G.; Aiello, F.C.; Szychlinska, M.A.; Di Rosa, M.; Castrogiovanni, P.; Mobasheri, A. Osteoarthritis in the XXIst Century: Risk Factors and Behaviours That Influence Disease Onset and Progression. Int. J. Mol. Sci. 2015, 16, 6093–6112. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, T.W.; McCabe, P.S.; McBeth, J. Update on the Epidemiology, Risk Factors and Disease Outcomes of Osteoarthritis. Best Pract. Res. Clin. Rheumatol. 2018, 32, 312–326. [Google Scholar] [CrossRef] [PubMed]
- Georgiev, T.; Angelov, A.K. Modifiable Risk Factors in Knee Osteoarthritis: Treatment Implications. Rheumatol. Int. 2019, 39, 1145–1157. [Google Scholar] [CrossRef] [PubMed]
- Chojnacki, M.; Kwapisz, A.; Synder, M.; Szemraj, J. Osteoarthritis: Etiology, Risk Factors, Molecular Mechanisms. Postep. Hig. Med. Dosw. 2014, 68, 640–652. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Moreno, M.; Rego, I.; Carreira-Garcia, V.; Blanco, F.J. Genetics in Osteoarthritis. Curr. Genom. 2008, 9, 542–547. [Google Scholar] [CrossRef]
- Hardcastle, S.A.; Dieppe, P.; Gregson, C.L.; Davey Smith, G.; Tobias, J.H. Osteoarthritis and Bone Mineral Density: Are Strong Bones Bad for Joints? Bonekey Rep. 2015, 4, 624. [Google Scholar] [CrossRef]
- Teichtahl, A.J.; Wang, Y.; Wluka, A.E.; Strauss, B.J.; Proietto, J.; Dixon, J.B.; Jones, G.; Cicuttini, F.M. Associations between Systemic Bone Mineral Density and Early Knee Cartilage Changes in Middle-Aged Adults without Clinical Knee Disease: A Prospective Cohort Study. Arthritis Res. Ther. 2017, 19, 98. [Google Scholar] [CrossRef] [PubMed]
- Nevitt, M.C.; Zhang, Y.; Javaid, M.K.; Neogi, T.; Curtis, J.R.; Niu, J.; McCulloch, C.E.; Segal, N.A.; Felson, D.T. High Systemic Bone Mineral Density Increases the Risk of Incident Knee OA and Joint Space Narrowing, but Not Radiographic Progression of Existing Knee OA: The MOST Study. Ann. Rheum. Dis. 2010, 69, 163–168. [Google Scholar] [CrossRef] [PubMed]
- Katz, J.N.; Arant, K.R.; Loeser, R.F. Diagnosis and Treatment of Hip and Knee Osteoarthritis: A Review. JAMA—J. Am. Med. Assoc. 2021, 325, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Hunter, D.J.; Bierma-Zeinstra, S. Osteoarthritis. Lancet 2019, 393, 1745–1759. [Google Scholar] [CrossRef] [PubMed]
- Altman, R.; Alarcon, G.; Appelrouth, D.; Bloch, D.; Borenstein, D.; Brandt, K.; Brown, C.; Cooke, T.D.; Daniel, W.; Gray, R.; et al. The American College of Rheumatology Criteria for the Classification and Reporting of Osteoarthritis of the Hand. Arthritis Rheum. 1990, 33, 1601–1610. [Google Scholar] [CrossRef] [PubMed]
- Altman, R.; Asch, E.; Bloch, D.; Bole, G.; Borenstein, D.; Brandt, K.; Christy, W.; Cooke, T.D.; Greenwald, R.; Hochberg, M.; et al. Development of Criteria for the Classification and Reporting of Osteoarthritis: Classification of Osteoarthritis of the Knee. Arthritis Rheum. 1986, 29, 1039–1049. [Google Scholar] [CrossRef] [PubMed]
- Ishibashi, O.; Hayashi, M.; Horikawa, A.; Owada, H.; Miyamoto, R.; Mizukami, N.; Inui, T. The Role of MiR-217-5p in the Puromycin Aminonucleoside-Induced Morphological Change of Podocytes. Noncoding RNA 2022, 8, 43. [Google Scholar] [CrossRef] [PubMed]
- Strmsek, Z.; Kunej, T. MicroRNA Silencing by DNA Methylation in Human Cancer: A Literature Analysis. Noncoding RNA 2015, 1, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Grenda, A.; Budzyński, M.; Filip, A. Biogenesis of MicroRNAs and Their Role in the Development and Course of Selected Hematologic Disorders. Postep. Hig. Med. Dosw. 2013, 67, 174–185. [Google Scholar] [CrossRef]
- Pu, M.; Chen, J.; Tao, Z.; Miao, L.; Qi, X.; Wang, Y.; Ren, J. Regulatory Network of MiRNA on Its Target: Coordination between Transcriptional and Post-Transcriptional Regulation of Gene Expression. Cell. Mol. Life Sci. 2019, 76, 441–451. [Google Scholar] [CrossRef]
- Meister, G.; Tuschl, T. Mechanisms of Gene Silencing by Double-Stranded RNA. Nature 2004, 431, 343–349. [Google Scholar] [CrossRef] [PubMed]
- Wakiyama, M.; Takimoto, K.; Ohara, O.; Yokoyama, S. Let-7 MicroRNA-Mediated MRNA Deadenylation and Translational Repression in a Mammalian Cell-Free System. Genes Dev. 2007, 21, 1857–1862. [Google Scholar] [CrossRef] [PubMed]
- Alwani, A.; Baj-Krzyworzeka, M. MiRNAs—Targets in Cancer Therapy. Postep. Biochem. 2021, 67. [Google Scholar] [CrossRef] [PubMed]
- Eastlack, S.C.; Alahari, S.K. MicroRNA and Breast Cancer: Understanding Pathogenesis, Improving Management. Noncoding RNA 2015, 1, 17–43. [Google Scholar] [CrossRef] [PubMed]
- Nedunchezhiyan, U.; Varughese, I.; Sun, A.R.J.; Wu, X.; Crawford, R.; Prasadam, I. Obesity, Inflammation, and Immune System in Osteoarthritis. Front. Immunol. 2022, 13, 907750. [Google Scholar] [CrossRef] [PubMed]
- Caldo, D.; Massarini, E.; Rucci, M.; Deaglio, S.; Ferracini, R. Epigenetics in Knee Osteoarthritis: A 2020–2023 Update Systematic Review. Life 2024, 14, 269. [Google Scholar] [CrossRef] [PubMed]
- Balaskas, P.; Goljanek-Whysall, K.; Clegg, P.D.; Fang, Y.; Cremers, A.; Smagul, A.; Welting, T.J.M.; Peffers, M.J. MicroRNA Signatures in Cartilage Ageing and Osteoarthritis. Biomedicines 2023, 11, 1189. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.C.; Chuang, S.M.; Hsu, C.J.; Tsai, C.H.; Wang, S.W.; Tang, C.H. CTGF Increases Vascular Endothelial Growth Factor-Dependent Angiogenesis in Human Synovial Fibroblasts by Increasing MiR-210 Expression. Cell Death Dis. 2014, 5, e1485. [Google Scholar] [CrossRef]
- Moran-Moguel, M.C.; Rio, S.P.D.; Mayorquin-Galvan, E.E.; Zavala-Cerna, M.G. Rheumatoid Arthritis and MiRNAs: A Critical Review through a Functional View. J. Immunol. Res. 2018, 2018, 2474529. [Google Scholar] [CrossRef]
- Robinson, W.H.; Lepus, C.M.; Wang, Q.; Raghu, H.; Mao, R.; Lindstrom, T.M.; Sokolove, J. Low-Grade Inflammation as a Key Mediator of the Pathogenesis of Osteoarthritis. Nat. Rev. Rheumatol. 2016, 12, 580–592. [Google Scholar] [CrossRef]
- Reyes, C.; Leyland, K.M.; Peat, G.; Cooper, C.; Arden, N.K.; Prieto-Alhambra, D. Association Between Overweight and Obesity and Risk of Clinically Diagnosed Knee, Hip, and Hand Osteoarthritis: A Population-Based Cohort Study. Arthritis Rheumatol. 2016, 68, 1869–1875. [Google Scholar] [CrossRef] [PubMed]
- Raud, B.; Gay, C.; Guiguet-Auclair, C.; Bonnin, A.; Gerbaud, L.; Pereira, B.; Duclos, M.; Boirie, Y.; Coudeyre, E. Level of Obesity Is Directly Associated with the Clinical and Functional Consequences of Knee Osteoarthritis. Sci. Rep. 2020, 10, 3601. [Google Scholar] [CrossRef] [PubMed]
- Galiniak, S.; Krawczyk-Marć, I.; Wawrzyniak, A.; Orkisz, S. Chondrocytes Apoptosis in Osteoarthritis. Postep. Hig. Med. Dosw. 2018, 72, 875–883. [Google Scholar] [CrossRef]
- Sandell, L.J.; Aigner, T. Articular Cartilage and Changes in Arthritis An Introduction: Cell Biology of Osteoarthritis. Arthritis Res. 2001, 3, 107. [Google Scholar] [CrossRef] [PubMed]
- Van Donkelaar, C.C.; Wilson, W. Mechanics of Chondrocyte Hypertrophy. Biomech. Model. Mechanobiol. 2012, 11, 655–664. [Google Scholar] [CrossRef] [PubMed]
- Dreier, R. Hypertrophic Differentiation of Chondrocytes in Osteoarthritis: The Developmental Aspect of Degenerative Joint Disorders. Arthritis Res. Ther. 2010, 12, 216. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Kim, W.J. The Role of Matrix Metalloproteinase in Inflammation with a Focus on Infectious Diseases. Int. J. Mol. Sci. 2022, 23, 10546. [Google Scholar] [CrossRef] [PubMed]
- Takahata, Y.; Murakami, T.; Hata, K.; Nishimura, R. Molecular Mechanisms Involved in the Progression and Protection of Osteoarthritis. Curr. Mol. Pharmacol. 2020, 14, 165–169. [Google Scholar] [CrossRef]
- Yamamoto, K.; Wilkinson, D.; Bou-Gharios, G. Targeting Dysregulation of Metalloproteinase Activity in Osteoarthritis. Calcif. Tissue Int. 2021, 109, 277–290. [Google Scholar] [CrossRef]
- Mehana, E.S.E.; Khafaga, A.F.; El-Blehi, S.S. The Role of Matrix Metalloproteinases in Osteoarthritis Pathogenesis: An Updated Review. Life Sci. 2019, 234, 116786. [Google Scholar] [CrossRef]
- Stannus, O.; Jones, G.; Cicuttini, F.; Parameswaran, V.; Quinn, S.; Burgess, J.; Ding, C. Circulating Levels of IL-6 and TNF-α Are Associated with Knee Radiographic Osteoarthritis and Knee Cartilage Loss in Older Adults. Osteoarthr. Cartil. 2010, 18, 1441–1447. [Google Scholar] [CrossRef] [PubMed]
- Attur, M.; Statnikov, A.; Samuels, J.; Li, Z.; Alekseyenko, A.V.; Greenberg, J.D.; Krasnokutsky, S.; Rybak, L.; Lu, Q.A.; Todd, J.; et al. Plasma Levels of Interleukin-1 Receptor Antagonist (IL1Ra) Predict Radiographic Progression of Symptomatic Knee Osteoarthritis. Osteoarthr. Cartil. 2015, 23, 1915–1924. [Google Scholar] [CrossRef] [PubMed]
- Chow, Y.Y.; Chin, K.Y. The Role of Inflammation in the Pathogenesis of Osteoarthritis. Mediat. Inflamm. 2020, 2020, 8293921. [Google Scholar] [CrossRef] [PubMed]
- Scanzello, C.R.; Umoh, E.; Pessler, F.; Diaz-Torne, C.; Miles, T.; DiCarlo, E.; Potter, H.G.; Mandl, L.; Marx, R.; Rodeo, S.; et al. Local Cytokine Profiles in Knee Osteoarthritis: Elevated Synovial Fluid Interleukin-15 Differentiates Early from End-Stage Disease. Osteoarthr. Cartil. 2009, 17, 1040–1048. [Google Scholar] [CrossRef] [PubMed]
- Dell’Isola, A.; Steultjens, M. Classification of Patients with Knee Osteoarthritis in Clinical Phenotypes: Data from the Osteoarthritis Initiative. PLoS ONE 2018, 13, e019104. [Google Scholar] [CrossRef] [PubMed]
- Mimpen, J.Y.; Baldwin, M.J.; Cribbs, A.P.; Philpott, M.; Carr, A.J.; Dakin, S.G.; Snelling, S.J.B. Interleukin-17A Causes Osteoarthritis-Like Transcriptional Changes in Human Osteoarthritis-Derived Chondrocytes and Synovial Fibroblasts In Vitro. Front. Immunol. 2021, 12, 676173. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xu, D.; Long, L.; Deng, X.; Tao, R.; Huang, G. Correlation between Plasma, Synovial Fluid and Articular Cartilage Interleukin-18 with Radiographic Severity in 33 Patients with Osteoarthritis of the Knee. Clin. Exp. Med. 2014, 14, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Shan, Y.; Qi, C.; Liu, Y.; Gao, H.; Zhao, D.; Jiang, Y. Increased Frequency of Peripheral Blood Follicular Helper T Cells and Elevated Serum IL-21 Levels in Patients with Knee Osteoarthritis. Mol. Med. Rep. 2017, 15, 1095–1102. [Google Scholar] [CrossRef] [PubMed]
- Deligne, C.; Casulli, S.; Pigenet, A.; Bougault, C.; Campillo-Gimenez, L.; Nourissat, G.; Berenbaum, F.; Elbim, C.; Houard, X. Differential Expression of Interleukin-17 and Interleukin-22 in Inflamed and Non-Inflamed Synovium from Osteoarthritis Patients. Osteoarthr. Cartil. 2015, 23, 1843–1852. [Google Scholar] [CrossRef]
- Yi, C.; Yi, Y.; Wei, J.; Jin, Q.; Li, J.; Sacitharan, P.K. Targeting IL-22 and IL-22R Protects against Experimental Osteoarthritis. Cell. Mol. Immunol. 2021, 18, 1329–1331. [Google Scholar] [CrossRef]
- Xiao, S.Q.; Cheng, M.; Wang, L.; Cao, J.; Fang, L.; Zhou, X.P.; He, X.J.; Hu, Y.F. The Role of Apoptosis in the Pathogenesis of Osteoarthritis. Int. Orthop. 2023, 47, 1895–1919. [Google Scholar] [CrossRef] [PubMed]
- Scanzello, C.R. Chemokines and Inflammation in Osteoarthritis: Insights from Patients and Animal Models. J. Orthop. Res. 2017, 35, 735–739. [Google Scholar] [CrossRef] [PubMed]
- Doi, H.; Nishida, K.; Yorimitsu, M.; Komiyama, T.; Kadota, Y.; Tetsunaga, T.; Yoshida, A.; Kubota, S.; Takigawa, M.; Ozaki, T. Interleukin-4 Downregulates the Cyclic Tensile Stress-Induced Matrix Metalloproteinases-13 and Cathepsin b Expression by Rat Normal Chondrocytes. Acta Med. Okayama 2008, 62, 119–126. [Google Scholar] [PubMed]
- Van Meegeren, M.E.R.; Roosendaal, G.; Jansen, N.W.D.; Wenting, M.J.G.; Van Wesel, A.C.W.; Van Roon, J.A.G.; Lafeber, F.P.J.G. IL-4 Alone and in Combination with IL-10 Protects against Blood-Induced Cartilage Damage. Osteoarthr. Cartil. 2012, 20, 764–772. [Google Scholar] [CrossRef] [PubMed]
- Schulze-Tanzil, G.; Zreiqat, H.; Sabat, R.; Kohl, B.; Halder, A.; Muller, R.; John, T. Interleukin-10 and Articular Cartilage: Experimental Therapeutical Approaches in Cartilage Disorders. Curr. Gene Ther. 2009, 9, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Barker, T.; Rogers, V.E.; Henriksen, V.T.; Trawick, R.H.; Momberger, N.G.; Lynn Rasmussen, G. Circulating IL-10 Is Compromised in Patients Predisposed to Developing and in Patients with Severe Knee Osteoarthritis. Sci. Rep. 2021, 11, 1812. [Google Scholar] [CrossRef]
- Fernandes, T.L.; Gomoll, A.H.; Lattermann, C.; Hernandez, A.J.; Bueno, D.F.; Amano, M.T. Macrophage: A Potential Target on Cartilage Regeneration. Front. Immunol. 2020, 11, 111. [Google Scholar] [CrossRef] [PubMed]
- Molnar, V.; Matišić, V.; Kodvanj, I.; Bjelica, R.; Jeleč, Ž.; Hudetz, D.; Rod, E.; Čukelj, F.; Vrdoljak, T.; Vidović, D.; et al. Cytokines and Chemokines Involved in Osteoarthritis Pathogenesis. Int. J. Mol. Sci. 2021, 22, 9208. [Google Scholar] [CrossRef] [PubMed]
- Primorac, D.; Molnar, V.; Matišić, V.; Hudetz, D.; Jeleč, Ž.; Rod, E.; Čukelj, F.; Vidović, D.; Vrdoljak, T.; Dobričić, B.; et al. Comprehensive Review of Knee Osteoarthritis Pharmacological Treatment and the Latest Professional Societies’ Guidelines. Pharmaceuticals 2021, 14, 205. [Google Scholar] [CrossRef]
- Verbruggen, G.; Wittoek, R.; Vander Cruyssen, B.; Elewaut, D. Tumour Necrosis Factor Blockade for the Treatment of Erosive Osteoarthritis of the Interphalangeal Finger Joints: A Double Blind, Randomised Trial on Structure Modification. Ann. Rheum. Dis. 2012, 71, 891–898. [Google Scholar] [CrossRef]
- Cohen, S.B.; Proudman, S.; Kivitz, A.J.; Burch, F.X.; Donohue, J.P.; Burstein, D.; Sun, Y.N.; Banfield, C.; Vincent, M.S.; Ni, L.; et al. A Randomized, Double-Blind Study of AMG 108 (a Fully Human Monoclonal Antibody to IL-1R1) in Patients with Osteoarthritis of the Knee. Arthritis Res. Ther. 2011, 13, R125. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, X.; Eymard, F.; Richette, P. Biologic Agents in Osteoarthritis: Hopes and Disappointments. Nat. Rev. Rheumatol. 2013, 9, 400–410. [Google Scholar] [CrossRef] [PubMed]
- Chevalier, X.; Goupille, P.; Beaulieu, A.D.; Burch, F.X.; Bensen, W.G.; Conrozier, T.; Loeuille, D.; Kivitz, A.J.; Silver, D.; Appleton, B.E. Intraarticular Injection of Anakinra in Osteoarthritis of the Knee: A Multicenter, Randomized, Double-Blind, Placebo-Controlled Study. Arthritis Care Res. 2009, 61, 344–352. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Yin, J.; Gao, J.; Cheng, T.S.; Pavlos, N.J.; Zhang, C.; Zheng, M.H. Subchondral Bone in Osteoarthritis: Insight into Risk Factors and Microstructural Changes. Arthritis Res. Ther. 2013, 15, 223. [Google Scholar] [CrossRef] [PubMed]
- Bobinac, D.; Spanjol, J.; Zoricic, S.; Maric, I. Changes in Articular Cartilage and Subchondral Bone Histomorphometry in Osteoarthritic Knee Joints in Humans. Bone 2003, 32, 284–290. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Cui, J.; Chu, L.; Zhang, W.; Xie, K.; Jiang, X.; He, Z.; Du, J.; Ai, S.; Sun, Q.; et al. Abnormal Subchondral Trabecular Bone Remodeling in Knee Osteoarthritis under the Influence of Knee Alignment. Osteoarthr. Cartil. 2022, 30, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Hu, W.; Chen, Y.; Dou, C.; Dong, S. Microenvironment in Subchondral Bone: Predominant Regulator for the Treatment of Osteoarthritis. Ann. Rheum. Dis. 2021, 80, 413–422. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Kirkwood, C.L.; Sohn, J.; Lau, A.; Bayers-Thering, M.; Bali, S.K.; Rachala, S.; Marzo, J.M.; Anders, M.J.; Beier, F.; et al. Expansion of Myeloid-Derived Suppressor Cells Contributes to Metabolic Osteoarthritis through Subchondral Bone Remodeling. Arthritis Res. Ther. 2021, 23, 287. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Chen, X.; Wang, S.; Jing, Y.; Su, J. Subchondral Bone Microenvironment in Osteoarthritis and Pain. Bone Res. 2021, 9, 20. [Google Scholar] [CrossRef]
- Burr, D.B.; Gallant, M.A. Bone Remodelling in Osteoarthritis. Nat. Rev. Rheumatol. 2012, 8, 665–673. [Google Scholar] [CrossRef]
- Chen, Y.; Hu, Y.; Yu, Y.E.; Zhang, X.; Watts, T.; Zhou, B.; Wang, J.; Wang, T.; Zhao, W.; Chiu, K.Y.; et al. Subchondral Trabecular Rod Loss and Plate Thickening in the Development of Osteoarthritis. J. Bone Miner. Res. 2018, 33, 316–327. [Google Scholar] [CrossRef]
- Yokota, S.; Ishizu, H.; Miyazaki, T.; Takahashi, D.; Iwasaki, N.; Shimizu, T. Osteoporosis, Osteoarthritis, and Subchondral Insufficiency Fracture: Recent Insights. Biomedicines 2024, 12, 843. [Google Scholar] [CrossRef] [PubMed]
- Goldring, S.R.; Goldring, M.B. Changes in the Osteochondral Unit during Osteoarthritis: Structure, Function and Cartilage Bone Crosstalk. Nat. Rev. Rheumatol. 2016, 12, 632–644. [Google Scholar] [CrossRef]
- Wu, X.; Liyanage, C.; Plan, M.; Stark, T.; McCubbin, T.; Barrero, R.A.; Batra, J.; Crawford, R.; Xiao, Y.; Prasadam, I. Dysregulated Energy Metabolism Impairs Chondrocyte Function in Osteoarthritis. Osteoarthr. Cartil. 2023, 31, 613–626. [Google Scholar] [CrossRef] [PubMed]
- Mobasheri, A.; Rayman, M.P.; Gualillo, O.; Sellam, J.; Van Der Kraan, P.; Fearon, U. The Role of Metabolism in the Pathogenesis of Osteoarthritis. Nat. Rev. Rheumatol. 2017, 13, 302–311. [Google Scholar] [CrossRef]
- Zheng, L.; Zhang, Z.; Sheng, P.; Mobasheri, A. The Role of Metabolism in Chondrocyte Dysfunction and the Progression of Osteoarthritis. Ageing Res. Rev. 2021, 66, 101249. [Google Scholar] [CrossRef] [PubMed]
- Maruotti, N.; Corrado, A.; Cantatore, F.P. Osteoblast Role in Osteoarthritis Pathogenesis. J. Cell. Physiol. 2017, 232, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Maglaviceanu, A.; Wu, B.; Kapoor, M. Fibroblast-like Synoviocytes: Role in Synovial Fibrosis Associated with Osteoarthritis. Wound Repair. Regen. 2021, 29, 642–649. [Google Scholar] [CrossRef]
- Li, T.; Peng, J.; Li, Q.; Shu, Y.; Zhu, P.; Hao, L. The Mechanism and Role of ADAMTS Protein Family in Osteoarthritis. Biomolecules 2022, 12, 959. [Google Scholar] [CrossRef]
- Grässel, S.; Zaucke, F.; Madry, H. Osteoarthritis: Novel Molecular Mechanisms Increase Our Understanding of the Disease Pathology. J. Clin. Med. 2021, 10, 1938. [Google Scholar] [CrossRef]
- Salman, L.A.; Ahmed, G.; Dakin, S.G.; Kendrick, B.; Price, A. Osteoarthritis: A Narrative Review of Molecular Approaches to Disease Management. Arthritis Res. Ther. 2023, 25, 27. [Google Scholar] [CrossRef] [PubMed]
- Schulze-Tanzil, G. Experimental Therapeutics for the Treatment of Osteoarthritis. J. Exp. Pharmacol. 2021, 13, 101–125. [Google Scholar] [CrossRef]
- Swingler, T.E.; Niu, L.; Smith, P.; Paddy, P.; Le, L.; Barter, M.J.; Young, D.A.; Clark, I.M. The Function of MicroRNAs in Cartilage and Osteoarthritis. Clin. Exp. Rheumatol. 2019, 37, 40–47. [Google Scholar] [PubMed]
- Tardif, G.; Hum, D.; Pelletier, J.P.; Duval, N.; Martel-Pelletier, J. Regulation of the IGFBP-5 and MMP-13 Genes by the MicroRNAs MiR-140 and MiR-27a in Human Osteoarthritic Chondrocytes. BMC Musculoskelet. Disord. 2009, 10, 148. [Google Scholar] [CrossRef] [PubMed]
- Nicolas, F.E.; Pais, H.; Schwach, F.; Lindow, M.; Kauppinen, S.; Moulton, V.; Dalmay, T. Experimental Identification of MicroRNA-140 Targets by Silencing and Overexpressing MiR-140. RNA 2008, 14, 2513–2520. [Google Scholar] [CrossRef]
- Pais, H.; Nicolas, F.E.; Soond, S.M.; Swingler, T.E.; Clark, I.M.; Chantry, A.; Moulton, V.; Dalmay, T. Analyzing MRNA Expression Identifies Smad3 as a MicroRNA-140 Target Regulated Only at Protein Level. RNA 2010, 16, 489–494. [Google Scholar] [CrossRef]
- Barter, M.J.; Tselepi, M.; Gõmez, R.; Woods, S.; Hui, W.; Smith, G.R.; Shanley, D.P.; Clark, I.M.; Young, D.A. Genome-Wide MicroRNA and Gene Analysis of Mesenchymal Stem Cell Chondrogenesis Identifies an Essential Role and Multiple Targets for MiR-140-5p. Stem Cells 2015, 33, 3266–3280. [Google Scholar] [CrossRef]
- Karlsen, T.A.; Jakobsen, R.B.; Mikkelsen, T.S.; Brinchmann, J.E. MicroRNA-140 Targets RALA and Regulates Chondrogenic Differentiation of Human Mesenchymal Stem Cells by Translational Enhancement of SOX9 and ACAN. Stem Cells Dev. 2014, 23, 290–304. [Google Scholar] [CrossRef]
- Nakamura, Y.; Inloes, J.B.; Katagiri, T.; Kobayashi, T. Chondrocyte-Specific MicroRNA-140 Regulates Endochondral Bone Development and Targets Dnpep to Modulate Bone Morphogenetic Protein Signaling. Mol. Cell. Biol. 2011, 31, 3019–3028. [Google Scholar] [CrossRef]
- Miyaki, S.; Sato, T.; Inoue, A.; Otsuki, S.; Ito, Y.; Yokoyama, S.; Kato, Y.; Takemoto, F.; Nakasa, T.; Yamashita, S.; et al. MicroRNA-140 Plays Dual Roles in Both Cartilage Development and Homeostasis. Genes Dev. 2010, 24, 1173–1185. [Google Scholar] [CrossRef]
- Papaioannou, G.; Inloes, J.B.; Nakamura, Y.; Paltrinieri, E.; Kobayashi, T. Let-7 and MiR-140 MicroRNAs Coordinately Regulate Skeletal Development. Proc. Natl. Acad. Sci. USA 2013, 110, E3291–E3300. [Google Scholar] [CrossRef] [PubMed]
- Paterson, M.R.; Kriegel, A.J. MiR-146a/b: A Family with Shared Seeds and Different Roots. Physiol. Genom. 2017, 49, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Yamasaki, K.; Nakasa, T.; Miyaki, S.; Ishikawa, M.; Deie, M.; Adachi, N.; Yasunaga, Y.; Asahara, H.; Ochi, M. Expression of MicroRNA-146a in Osteoarthritis Cartilage. Arthritis Rheum. 2009, 60, 1035–1041. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Gibson, G.; Kim, J.S.; Kroin, J.; Xu, S.; van Wijnen, A.J.; Im, H.J. MicroRNA-146a Is Linked to Pain-Related Pathophysiology of Osteoarthritis. Gene 2011, 480, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Gao, X.; Wang, J.; Yang, C.; Wang, Y.; Liu, Y.; Zou, W.; Liu, T. Hypoxia-Induced MicroRNA-146a Represses Bcl-2 through Traf6/IRAK1 but Not Smad4 to Promote Chondrocyte Autophagy. Biol. Chem. 2017, 398, 499–507. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Huang, J.; Dai, L.; Yu, D.; Chen, Q.; Zhang, X.; Dai, K. MiR-146a, an IL-1β Responsive MiRNA, Induces Vascular Endothelial Growth Factor and Chondrocyte Apoptosis by Targeting Smad4. Arthritis Res. Ther. 2012, 14, R75. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Wang, J.; Chu, J.; Yang, C.; Xiao, H.; Zhao, C.; Sun, Z.; Gao, X.; Chen, G.; Han, Z.; et al. MicroRNA-146a Induced by Hypoxia Promotes Chondrocyte Autophagy through Bcl-2. Cell. Physiol. Biochem. 2015, 37, 1442–1453. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zheng, W.; Li, D.; Zheng, J. MiR-146a-5p Promotes Chondrocyte Apoptosis and Inhibits Autophagy of Osteoarthritis by Targeting NUMB. Cartilage 2021, 13, 1467S–1477S. [Google Scholar] [CrossRef] [PubMed]
- Qin, H.; Wang, C.; He, Y.; Lu, A.; Li, T.; Zhang, B.; Shen, J. Silencing MiR-146a-5p Protects against Injury-Induced Osteoarthritis in Mice. Biomolecules 2023, 13, 123. [Google Scholar] [CrossRef]
- Guan, Y.J.; Li, J.; Yang, X.; Du, S.; Ding, J.; Gao, Y.; Zhang, Y.; Yang, K.; Chen, Q. Evidence That MiR-146a Attenuates Aging- and Trauma-Induced Osteoarthritis by Inhibiting Notch1, IL-6, and IL-1 Mediated Catabolism. Aging Cell 2018, 17, e12752. [Google Scholar] [CrossRef]
- Liu, J.N.; Lu, S.; Fu, C.M. MiR-146a Expression Profiles in Osteoarthritis in Different Tissue Sources: A Meta-Analysis of Observational Studies. J. Orthop. Surg. Res. 2022, 17, 148. [Google Scholar] [CrossRef]
- Papathanasiou, I.; Mourmoura, E.; Balis, C.; Tsezou, A. Impact of MiR-SNP Rs2910164 on MiR-146a Expression in Osteoarthritic Chondrocytes. Adv. Med. Sci. 2020, 65, 78–85. [Google Scholar] [CrossRef]
- Kopańska, M.; Szala, D.; Czech, J.; Gabło, N.; Gargasz, K.; Trzeciak, M.; Zawlik, I.; Snela, S. MiRNA Expression in the Cartilage of Patients with Osteoarthritis. J. Orthop. Surg. Res. 2017, 12, 51. [Google Scholar] [CrossRef]
- Hu, Q.; Ecker, M. Overview of MMP-13 as a Promising Target for the Treatment of Osteoarthritis. Int. J. Mol. Sci. 2021, 22, 1742. [Google Scholar] [CrossRef]
- Song, J.; Kim, D.; Chun, C.H.; Jin, E.J. MicroRNA-9 Regulates Survival of Chondroblasts and Cartilage Integrity by Targeting Protogenin. Cell Commun. Signal. 2013, 11, 66. [Google Scholar] [CrossRef]
- Li, H.Z.; Xu, X.H.; Lin, N.; Wang, D.W.; Lin, Y.M.; Su, Z.Z.; Lu, H.D. Overexpression of MiR-10a-5p Facilitates the Progression of Osteoarthritis. Aging 2020, 12, 5948–5976. [Google Scholar] [CrossRef] [PubMed]
- Iliopoulos, D.; Malizos, K.N.; Oikonomou, P.; Tsezou, A. Integrative MicroRNA and Proteomic Approaches Identify Novel Osteoarthritis Genes and Their Collaborative Metabolic and Inflammatory Networks. PLoS ONE 2008, 3, e3740. [Google Scholar] [CrossRef] [PubMed]
- Sheng, W.; Wang, Q.; Qin, H.; Cao, S.; Wei, Y.; Weng, J.; Yu, F.; Zeng, H. Osteoarthritis: Role of Peroxisome Proliferator-Activated Receptors. Int. J. Mol. Sci. 2023, 24, 13137. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.Y.; Chanalaris, A.; Troeberg, L. ADAMTS and ADAM Metalloproteinases in Osteoarthritis—Looking beyond the ‘Usual Suspects’. Osteoarthr. Cartil. 2017, 25, 1000–1009. [Google Scholar] [CrossRef]
- Endisha, H.; Datta, P.; Sharma, A.; Nakamura, S.; Rossomacha, E.; Younan, C.; Ali, S.A.; Tavallaee, G.; Lively, S.; Potla, P.; et al. MicroRNA-34a-5p Promotes Joint Destruction during Osteoarthritis. Arthritis Rheumatol. 2021, 73, 426–439. [Google Scholar] [CrossRef]
- Miyaki, S.; Nakasa, T.; Otsuki, S.; Grogan, S.P.; Higashiyama, R.; Inoue, A.; Kato, Y.; Sato, T.; Lotz, M.K.; Asahara, H. MicroRNA-140 Is Expressed in Differentiated Human Articular Chondrocytes and Modulates Interleukin-1 Responses. Arthritis Rheum. 2009, 60, 2723–2730. [Google Scholar] [CrossRef] [PubMed]
- Law, Y.Y.; Lee, W.F.; Hsu, C.J.; Lin, Y.Y.; Tsai, C.H.; Huang, C.C.; Wu, M.H.; Tang, C.H.; Liu, J.F. MiR-Let-7c-5p and MiR-149-5p Inhibit Proinflammatory Cytokine Production in Osteoarthritis and Rheumatoid Arthritis Synovial Fibroblasts. Aging 2021, 13, 17227–17236. [Google Scholar] [CrossRef] [PubMed]
- Santini, P.; Politi, L.; Vedova, P.D.; Scandurra, R.; Scotto D’Abusco, A. The Inflammatory Circuitry of MiR-149 as a Pathological Mechanism in Osteoarthritis. Rheumatol. Int. 2014, 34, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Zhang, L.; Tian, H. MicroRNA-149 Improves Osteoarthritis via Repression of VCAM-1 and Inactivation of PI3K/AKT Pathway. Exp. Gerontol. 2023, 174, 112103. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Wu, S.; Wu, Y.; Chen, L.; Pang, Q. MiR-149 Suppresses the Inflammatory Response of Chondrocytes in Osteoarthritis by down-Regulating the Activation of TAK1/NF-ΚB. Biomed. Pharmacother. 2018, 101, 763–768. [Google Scholar] [CrossRef] [PubMed]
- Lian, W.S.; Ko, J.Y.; Wu, R.W.; Sun, Y.C.; Chen, Y.S.; Wu, S.L.; Weng, L.H.; Jahr, H.; Wang, F.S. MicroRNA-128a Represses Chondrocyte Autophagy and Exacerbates Knee Osteoarthritis by Disrupting Atg12. Cell Death Dis. 2018, 9, 919. [Google Scholar] [CrossRef] [PubMed]
- Ko, J.-Y.; Wang, F.-S.; Lian, W.-S.; Fang, H.-C.; Kuo, S.-J. Cartilage-Specific Knockout of MiRNA-128a Expression Normalizes the Expression of Circadian Clock Genes (CCGs) and Mitigates the Severity of Osteoarthritis. Biomed. J. 2024, 47, 100629. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, Y.; Tokuzawa, Y.; Ninomiya, Y.; Yagi, K.; Yatsuka-Kanesaki, Y.; Suda, T.; Fukuda, T.; Katagiri, T.; Kondoh, Y.; Amemiya, T.; et al. MiR-210 Promotes Osteoblastic Differentiation through Inhibition of AcvR1b. FEBS Lett. 2009, 583, 2263–2268. [Google Scholar] [CrossRef]
- Qi, J.; Qiao, Y.; Wang, P.; Li, S.; Zhao, W.; Gao, C. MicroRNA-210 Negatively Regulates LPS-Induced Production of Proinflammatory Cytokines by Targeting NF-ΚB1 in Murine Macrophages. FEBS Lett. 2012, 586, 1201–1207. [Google Scholar] [CrossRef]
- Zhang, D.; Cao, X.; Li, J.; Zhao, G. MiR-210 Inhibits NF-ΚB Signaling Pathway by Targeting DR6 in Osteoarthritis. Sci. Rep. 2015, 5, 12775. [Google Scholar] [CrossRef]
- Lu, X.; Li, Y.; Chen, H.; Pan, Y.; Lin, R.; Chen, S. MiR-335-5P Contributes to Human Osteoarthritis by Targeting HBP1. Exp. Ther. Med. 2020, 21, 109. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.O.; Zhang, L.; Tang, Z.Y.; Gong, Z.M. MiR-485-5p Promotes the Development of Osteoarthritis by Inhibiting Cartilage Differentiation in BMSCs. Eur. Rev. Med. Pharmacol. Sci. 2021, 22, 3294–3302. [Google Scholar] [CrossRef]
- Akhtar, N.; Rasheed, Z.; Ramamurthy, S.; Anbazhagan, A.N.; Voss, F.R.; Haqqi, T.M. MicroRNA-27b Regulates the Expression of Matrix Metalloproteinase 13 in Human Osteoarthritis Chondrocytes. Arthritis Rheum. 2010, 62, 1361–1371. [Google Scholar] [CrossRef] [PubMed]
- Tavallaee, G.; Lively, S.; Rockel, J.S.; Ali, S.A.; Im, M.; Sarda, C.; Mitchell, G.M.; Rossomacha, E.; Nakamura, S.; Potla, P.; et al. Contribution of MicroRNA-27b-3p to Synovial Fibrotic Responses in Knee Osteoarthritis. Arthritis Rheumatol. 2022, 74, 1928–1942. [Google Scholar] [CrossRef] [PubMed]
- Lv, S.; Xu, J.; Chen, L.; Wu, H.; Feng, W.; Zheng, Y.; Li, P.; Zhang, H.; Zhang, L.; Chi, G.; et al. MicroRNA-27b Targets CBFB to Inhibit Differentiation of Human Bone Marrow Mesenchymal Stem Cells into Hypertrophic Chondrocytes. Stem Cell Res. Ther. 2020, 11, 392. [Google Scholar] [CrossRef] [PubMed]
- Tu, M.; Li, Y.; Zeng, C.; Deng, Z.; Gao, S.; Xiao, W.; Luo, W.; Jiang, W.; Li, L.; Lei, G. MicroRNA-127-5p Regulates Osteopontin Expression and Osteopontin-Mediated Proliferation of Human Chondrocytes. Sci. Rep. 2016, 6, 25032. [Google Scholar] [CrossRef] [PubMed]
- Park, S.J.; Cheon, E.J.; Lee, M.H.; Kim, H.A. MicroRNA-127-5p Regulates Matrix Metalloproteinase 13 Expression and Interleukin-1β-Induced Catabolic Effects in Human Chondrocytes. Arthritis Rheum. 2013, 65, 3141–3152. [Google Scholar] [CrossRef]
- Hu, Y.; Liu, H.X.; Xu, D.; Xue, X.; Xu, X. The Anti-Inflammatory Effect of MiR-140-3p in BMSCs-Exosomes on Osteoarthritis. Acta Chir. Orthop. Traumatol. Cech. 2023, 90, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Karlsen, T.A.; de Souza, G.A.; Ødegaard, B.; Engebretsen, L.; Brinchmann, J.E. MicroRNA-140 Inhibits Inflammation and Stimulates Chondrogenesis in a Model of Interleukin 1β-Induced Osteoarthritis. Mol. Ther. Nucleic Acids 2016, 5, e373. [Google Scholar] [CrossRef]
- Liang, Y.; Duan, L.; Xiong, J.; Zhu, W.; Liu, Q.; Wang, D.; Liu, W.; Li, Z.; Wang, D. E2 Regulates MMP-13 via Targeting MiR-140 in IL-1β-Induced Extracellular Matrix Degradation in Human Chondrocytes. Arthritis Res. Ther. 2016, 18, 105. [Google Scholar] [CrossRef]
- Li, W.; Zhao, S.; Yang, H.; Zhang, C.; Kang, Q.; Deng, J.; Xu, Y.; Ding, Y.; Li, S. Potential Novel Prediction of TMJ-OA: MiR-140-5p Regulates Inflammation through Smad/TGF-β Signaling. Front. Pharmacol. 2019, 9, 15. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Shen, S.; Li, Z.; Li, W.; Weng, X. MIR-140-5p Affects Chondrocyte Proliferation, Apoptosis, and Inflammation by Targeting HMGB1 in Osteoarthritis. Inflamm. Res. 2020, 69, 63–73. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Hu, J.; Pan, Y.; Shan, Y.; Jiang, L.; Qi, X.; Jia, L. MiR-140-5p/MiR-149 Affects Chondrocyte Proliferation, Apoptosis, and Autophagy by Targeting FUT1 in Osteoarthritis. Inflammation 2018, 41, 959–971. [Google Scholar] [CrossRef] [PubMed]
- Tuddenham, L.; Wheeler, G.; Ntounia-Fousara, S.; Waters, J.; Hajihosseini, M.K.; Clark, I.; Dalmay, T. The Cartilage Specific MicroRNA-140 Targets Histone Deacetylase 4 in Mouse Cells. FEBS Lett. 2006, 580, 4214–4217. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Hu, Q.; Chen, Z.; Shen, B.; Yang, J.; Kang, P.; Zhou, Z.; Pei, F. MicroRNA-140 Suppresses Human Chondrocytes Hypertrophy by Targeting SMAD1 and Controlling the Bone Morphogenetic Protein Pathway in Osteoarthritis. Am. J. Med. Sci. 2018, 355, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Taganov, K.D.; Boldin, M.P.; Chang, K.J.; Baltimore, D. NF-ΚB-Dependent Induction of MicroRNA MiR-146, an Inhibitor Targeted to Signaling Proteins of Innate Immune Responses. Proc. Natl. Acad. Sci. USA 2006, 103, 12481–12486. [Google Scholar] [CrossRef]
- Jones, S.W.; Watkins, G.; Le Good, N.; Roberts, S.; Murphy, C.L.; Brockbank, S.M.V.; Needham, M.R.C.; Read, S.J.; Newham, P. The Identification of Differentially Expressed MicroRNA in Osteoarthritic Tissue That Modulate the Production of TNF-α and MMP13. Osteoarthr. Cartil. 2009, 17, 464–472. [Google Scholar] [CrossRef] [PubMed]
- Skrzypa, M.; Szala, D.; Gablo, N.; Czech, J.; Pajak, J.; Kopanska, M.; Trzeciak, M.; Gargasz, K.; Snela, S.; Zawlik, I. MiRNA-146a-5p Is Upregulated in Serum and Cartilage Samples of Patients with Osteoarthritis. Pol. J. Surg. 2019, 91, 1–5. [Google Scholar] [CrossRef]
- Kapinas, K.; Delany, A.M. MicroRNA Biogenesis and Regulation of Bone Remodeling. Arthritis Res. Ther. 2011, 13, 220. [Google Scholar] [CrossRef]
- Liu, H.; Yan, L.; Li, X.; Li, D.; Wang, G.; Shen, N.N.; Li, J.J.; Wang, B. MicroRNA Expression in Osteoarthritis: A Meta-Analysis. Clin. Exp. Med. 2023, 23, 3737–3749. [Google Scholar] [CrossRef]
- Portal-Núñez, S.; Esbrit, P.; Alcaraz, M.J.; Largo, R. Oxidative Stress, Autophagy, Epigenetic Changes and Regulation by MiRNAs as Potential Therapeutic Targets in Osteoarthritis. Biochem. Pharmacol. 2016, 108, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Oliviero, A.; Della Porta, G.; Peretti, G.M.; Maffulli, N. MicroRNA in Osteoarthritis: Physiopathology, Diagnosis and Therapeutic Challenge. Br. Med. Bull. 2019, 130, 137–147. [Google Scholar] [CrossRef] [PubMed]
- Qin, X.B.; Wen, K.; Wu, X.X.; Yao, Z.J. MiR-183 Regulates the Differentiation of Osteoblasts in the Development of Osteoporosis by Targeting Smad4. Acta Histochem. 2021, 123, 151786. [Google Scholar] [CrossRef] [PubMed]
- Briskin, D.; Wang, P.Y.; Bartel, D.P. The Biochemical Basis for the Cooperative Action of MicroRNAs. Proc. Natl. Acad. Sci. USA 2020, 117, 17764–17774. [Google Scholar] [CrossRef]
- Zhong, L.; Huang, X.; Karperien, M.; Post, J.N. Correlation between Gene Expression and Osteoarthritis Progression in Human. Int. J. Mol. Sci. 2016, 17, 1126. [Google Scholar] [CrossRef]

| Group of Risk Factors | Risk Factors | References |
|---|---|---|
| demographic | female; older age | [13,15,16,17,18,19,20,21,22,23] |
| body weight | overweight and obesity | [13,14,15,16,17,18,19] |
| genetic mutations | e.g., mutation of the COL2A1, COL11A, COL11A2, COL1A1, and COL9A1 gene | [17,21,29,30] |
| mechanical factors | professional work requiring frequent knee bending and significant use of manual dexterity; practicing competitive sports in the past; weakness of periarticular skeletal muscles; sedentary lifestyle; intense recreational running; past injuries; previous knee surgery | [16,17,18,21,22,24,25,26,27,28,29] |
| individual factors | high bone mineral density | [27,31,32,33] |
| diseases | disturbances of deep sensation | [24] |
| Categories | Factors |
|---|---|
| developmental and congenital defects | |
| local diseases | aseptic necrosis of the femoral head in children; congenital hip dysplasia; exfoliation of the bone epiphysis |
| mechanical factors | difference in the length of the lower limbs; valgus or varus; joint hypermobility syndrome |
| dieseases | |
| metabolic | ochronosis; hereditary hemochromatosis; Wilson’s disease; Gaucher’s disease |
| endocrine | acromegaly; hyperparathyroidism; diabetes; obesity; hypothyroidism |
| from the deposition of calcium salts | chondrocalcinosis; apatite arthropathy |
| endemic diseases | Kashin/Beck disease; Mseleni disease |
| other bone and joint diseases | local: fractures; aseptic necrosis; infections; gout |
| disseminated: rheumatoid arthritis; Paget’s disease; osteopeyrosis; osteochondritis; other inflammations | |
| neurodystrophy of bones and joint | |
| other diseases | hemoglobinopathies; caisson disease |
| other factors | |
| injuries | acute; chronic |
| external factors | frostbite |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Szala, D.; Kopańska, M.; Trojniak, J.; Jabłoński, J.; Hanf-Osetek, D.; Snela, S.; Zawlik, I. The Role of MicroRNAs in the Pathophysiology of Osteoarthritis. Int. J. Mol. Sci. 2024, 25, 6352. https://doi.org/10.3390/ijms25126352
Szala D, Kopańska M, Trojniak J, Jabłoński J, Hanf-Osetek D, Snela S, Zawlik I. The Role of MicroRNAs in the Pathophysiology of Osteoarthritis. International Journal of Molecular Sciences. 2024; 25(12):6352. https://doi.org/10.3390/ijms25126352
Chicago/Turabian StyleSzala, Dariusz, Marta Kopańska, Julia Trojniak, Jarosław Jabłoński, Dorota Hanf-Osetek, Sławomir Snela, and Izabela Zawlik. 2024. "The Role of MicroRNAs in the Pathophysiology of Osteoarthritis" International Journal of Molecular Sciences 25, no. 12: 6352. https://doi.org/10.3390/ijms25126352
APA StyleSzala, D., Kopańska, M., Trojniak, J., Jabłoński, J., Hanf-Osetek, D., Snela, S., & Zawlik, I. (2024). The Role of MicroRNAs in the Pathophysiology of Osteoarthritis. International Journal of Molecular Sciences, 25(12), 6352. https://doi.org/10.3390/ijms25126352