
Mizoribine Promotes Molecular Chaperone HSP60/HSP10 Complex Formation



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
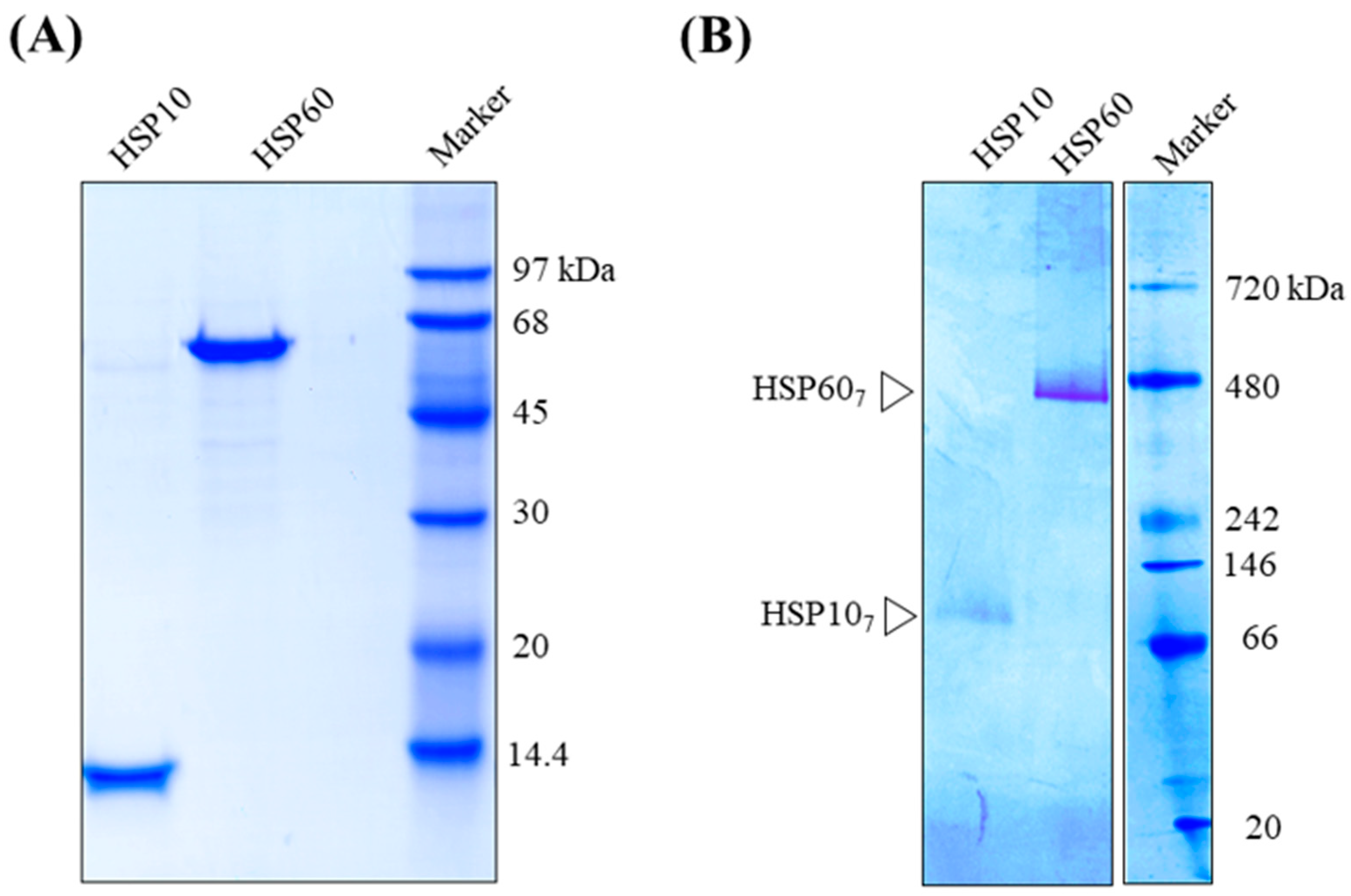
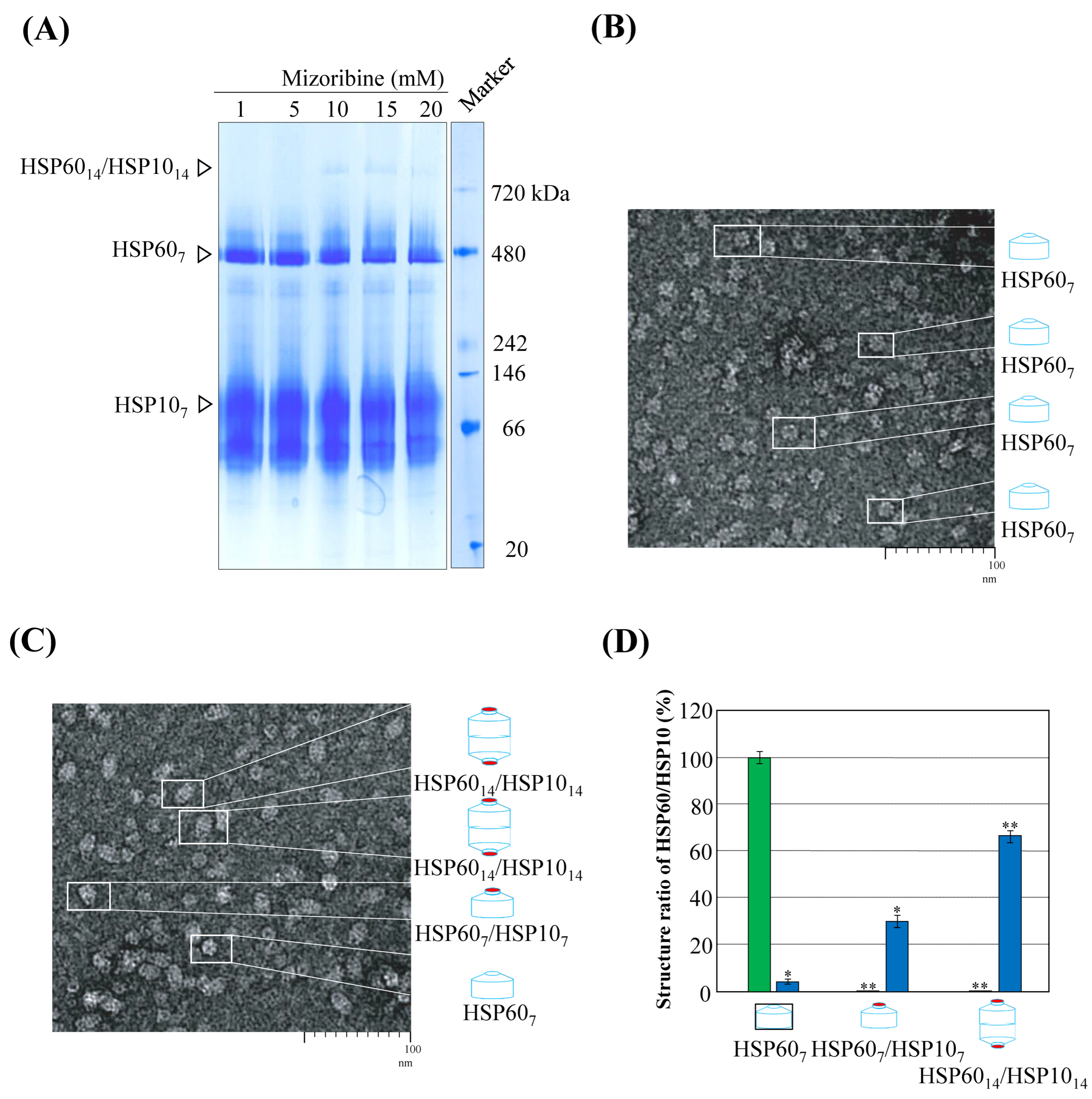
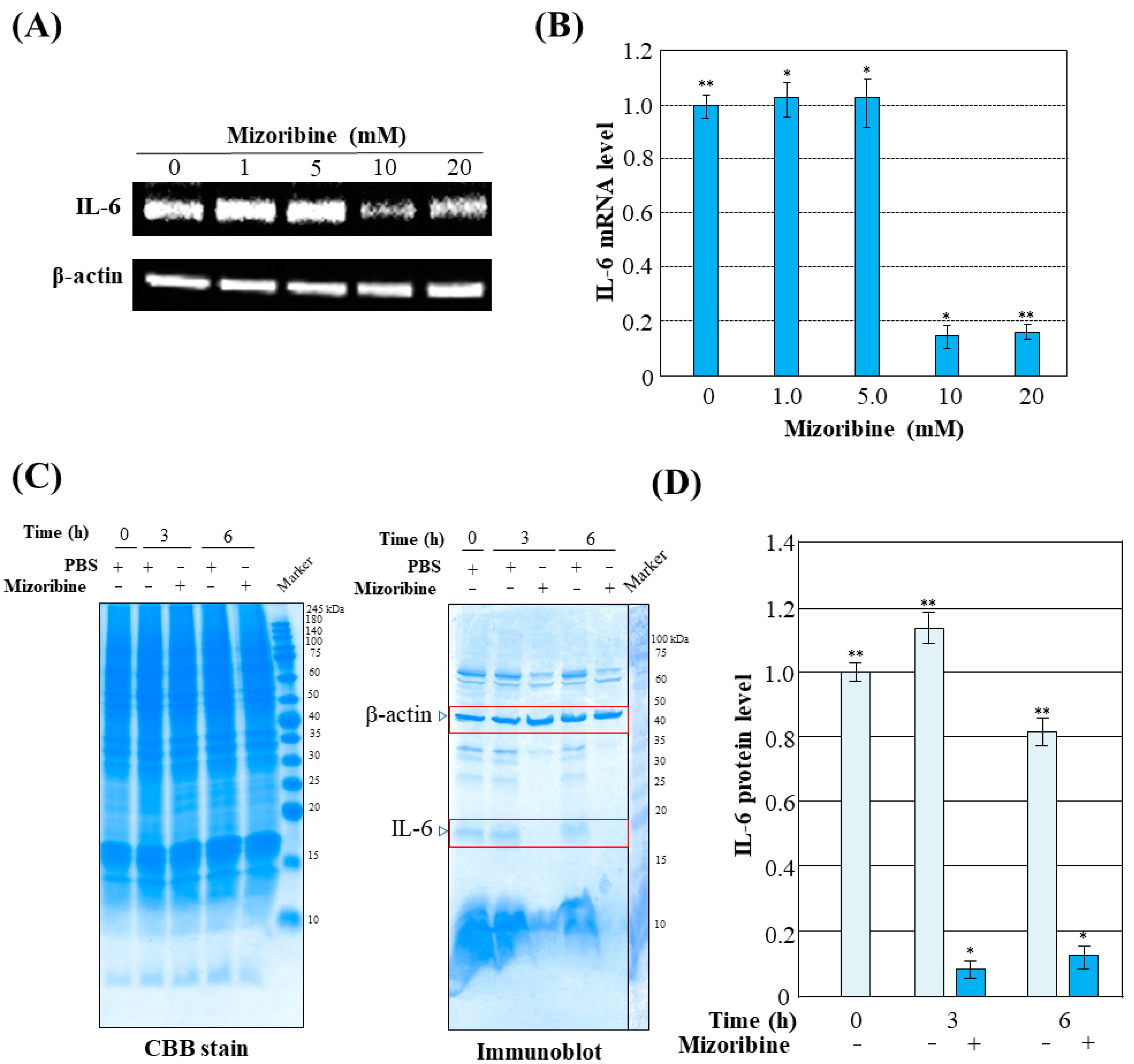
2. Results
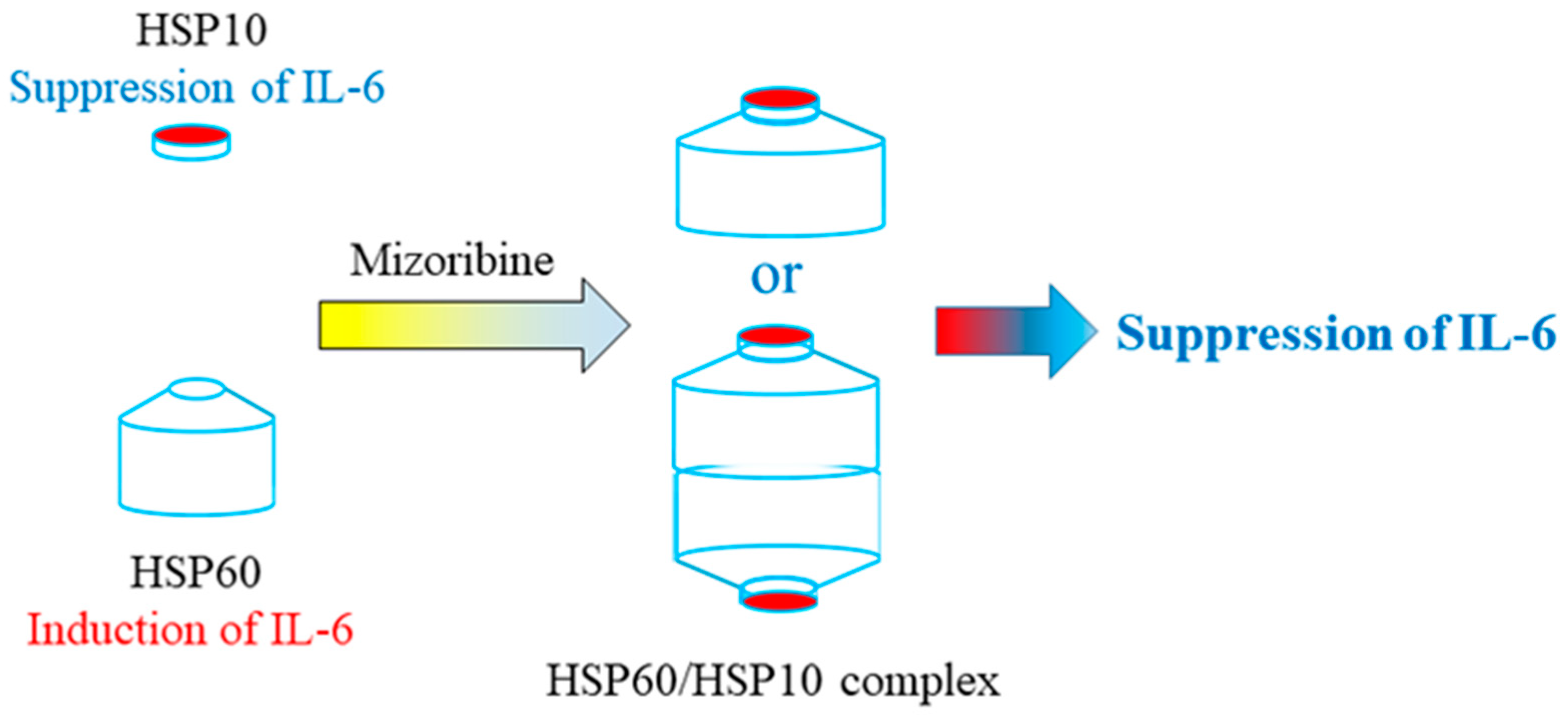
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Structural Analysis of Human HSP60 and HSP10
4.3. Cell Culture and Cell Differentiation
4.4. Analysis of mRNA Levels and Protein Levels of IL-6
4.5. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ATP | Adenosine triphosphate |
| BCIP | 5-Bromo-4-chloro-3-indolyl-phosphate |
| DMEM | Dulbecco’s converted Eagle’s |
| DMARD | Disease-modifying anti-rheumatic drug |
| ERS | Electrical resistance system |
| FBS | Fetal Bovine Serum |
| GMP | Guanosine monophosphate |
| HSP10 | 10-kDa Heat Shock Protein |
| HSP60 | 60-kDa Heat Shock Protein |
| GMP | Guanosine monophosphate |
| GTP | Guanosine triphosphate |
| ICAM | Intercellular adhesion molecule |
| IL-6 | Interleukin-6 |
| IMP | Inosine Mono Phosphate |
| KC | Mouse Chemokine CXCL1 |
| LPS | Lipopolysaccharide |
| MCP-1 | Monocyte chemoattractant protein-1 |
| NBT | Nitro Blue Tetrazolium |
| NF-κB | Nuclear factor-kappa B |
| PBS | Phosphate-Buffered Saline |
| PET | Polyethylene terephthalate |
| PMA | Phorbol12-myristate13-acetate |
| PMSF | Phenylmethylsulfonyl fluoride |
| RT-PCR | Reverse Transcription Polymerase Chain Reaction |
| TEER | Trans Epithelial Electrical Resistance |
| TEM | Transmission Electron Microscopy |
| TNF-α | Tumor Necrosis Factor-α |
| VCAM | Vascular cell adhesion molecule |
References
- Iwata, H.; Iwaki, H.; Masukawa, T.; Kasamatsu, S.; Okamoto, H. Anti-arthritic activity of bredinin, an immunosuppressive agent. Experientia 1977, 33, 502–503. [Google Scholar] [CrossRef] [PubMed]
- Koyama, H.; Tsuji, M. Genetic and biochemical studies on the activation and cytotoxic mechanism of bredinin, a potent inhibitor of purine biosynthesis in mammalian cells. Biochem. Pharmacol. 1983, 32, 3547–3553. [Google Scholar] [CrossRef] [PubMed]
- Kusumi, T.; Tsuda, M.; Katsunuma, T.; Yamamura, M. Dual inhibitory effect of bredinin. Cell Biochem. Funct. 1989, 7, 201–204. [Google Scholar] [CrossRef] [PubMed]
- Nakajo, A.; Khoshnoodi, J.; Takenaka, H.; Hagiwara, E.; Watanabe, T.; Kawakami, H.; Kurayama, R.; Sekine, Y.; Bessho, F.; Takahashi, S.; et al. Mizoribine corrects defective nephrin biogenesis by restoring intracellular energy balance. J. Am. Soc. Nephrol. 2007, 18, 2554–2564. [Google Scholar] [CrossRef] [PubMed]
- Turka, L.A.; Dayton, J.; Sinclair, G.; Thompson, C.B.; Mitchell, B.S. Guanine ribonucleotide depletion inhibits T-cell activation. Mechanism of action of the immunosuppressive drug mizoribine. J. Clin. Investig. 1991, 87, 940–948. [Google Scholar] [CrossRef] [PubMed]
- Ichinose, K.; Origuchi, T.; Kawashiri, S.Y.; Iwamoto, N.; Fujikawa, K.; Aramaki, T.; Kamachi, M.; Arima, K.; Tamai, M.; Nakamura, H.; et al. Efficacy and safety of mizoribine by one single dose administration for patients with rheumatoid arthritis. Intern. Med. 2010, 49, 2211–2218. [Google Scholar] [CrossRef] [PubMed]
- Aihara, Y.; Miyamae, T.; Ito, S.; Kobayashi, S.; Imagawa, T.; Mori, M.; Ibe, M.; Mitsuda, T.; Yokota, S. Mizoribine as an effective combined maintenance therapy with prednisolone in child-onset systemic lupus erythematosus. Pediatr. Int. 2002, 44, 199–204. [Google Scholar] [CrossRef]
- Yokota, S. Mizoribine: Mode of action and effects in clinical use. Pediatr. Int. 2002, 44, 196–198. [Google Scholar] [CrossRef]
- Kol, A.; Bourcier, T.; Lichtman, A.; Libby, P. Chlamydial and human heat shock protein 60s activate human vascular endothelium, smooth muscle cells, and macrophages. J. Clin. Investig. 1999, 103, 571–577. [Google Scholar] [CrossRef]
- Kol, A.; Lichtman, A.H.; Finberg, R.W.; Libby, P.; Kurt-Jones, E.A. Heat shock protein (HSP) 60 activates the innate immune response: CD14 is an essential receptor for HSP60 activation of mononuclear cells. J. Immunol. 2000, 164, 3–17. [Google Scholar] [CrossRef]
- Galdiero, M.; de l’Ero, G.C.; Marcatili, A. Cytokine and adhesion molecule expression in human monocytes and endothelial cells stimulated with bacterial heat shock proteins. Infect. Immun. 1997, 65, 699–707. [Google Scholar] [CrossRef] [PubMed]
- Gülden, E.; Märker, T.; Kriebel, J.; Kolb-Bachofen, V.; Burkart, V.; Habich, C. Heat shock protein 60: Evidence for receptor-mediated induction of proinflammatory mediators during adipocyte differentiation. FEBS Lett. 2009, 583, 2877–2881. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.J.; Le, T.T.; Dobbin, C.A.; Banovic, T.; Howard, C.B.; Flores, F.D.M.L.; Vanags, D.; Naylor, D.J.; Hill, G.R.; Suhrbier, A. Heat shock protein 10 inhibits lipopolysaccharide-induced inflammatory mediator production. J. Biol. Chem. 2005, 280, 4037–4047. [Google Scholar] [CrossRef] [PubMed]
- Itoh, H.; Komatsuda, A.; Wakui, H.; Miura, A.B.; Tashima, Y. Mammalian HSP60 is a major target for an immunosuppressant mizoribine. J. Biol. Chem. 1999, 274, 35147–35151. [Google Scholar] [CrossRef] [PubMed]
- Nisemblat, S.; Yaniv, O.; Parnas, A.; Frolow, F.; Azem, A. Crystal structure of the human mitochondrial chaperonin symmetrical football complex. Proc. Natl. Acad. Sci. USA 2015, 112, 6044–6049. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, T.; Ishida, R.; Yamamoto, H.; Tanabe-Ishida, M.; Haga, A.; Takahashi, H.; Takahashi, K.; Goto, D.; Grave, E.; Itoh, H. Functional structure and physiological functions of mammalian wild-type HSP60. Arch. Biochem. Biophys. 2015, 586, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, T.; Yamamoto, H.; Kudo, I.; Matsumoto, K.; Odaka, M.; Grave, E.; Itoh, H. HSP60 possesses a GTPase activity and mediates protein folding with HSP10. Sci. Rep. 2017, 7, e16931. [Google Scholar] [CrossRef]
- Ishida, R.; Okamoto, T.; Motojima, F.; Kubota, H.; Takahashi, H.; Tanabe, M.; Oka, T.; Kitamura, A.; Kinjo, M.; Yoshida, M.; et al. Physicochemical Properties of the Mammalian Molecular Chaperone HSP60. Int. J. Mol. Sci. 2018, 19, e489. [Google Scholar] [CrossRef] [PubMed]
- Kawasaki, Y. Mizoribine: A new approach in the treatment of renal disease. Clin. Dev. Immunol. 2009, 2009, e681482. [Google Scholar] [CrossRef]
- Kamata, K.; Okubo, M.; Ishigamori, E.; Masaki, Y.; Uchida, H.; Watanabe, K.; Kashiwagi, N. Immunosuppressive effect of bredinin on cell-mediated and humoral immune reactions in experimental animals. Transplantation 1983, 35, 144–149. [Google Scholar] [CrossRef]
- Mizuno, K.; Tsujino, M.; Takada, M.; Hayashi, M.; Atsumi, K. Studies on bredinin. I. Isolation, characterization, and biological properties. J. Antibiot. 1974, 27, 775–782. [Google Scholar] [CrossRef]
- Hirohata, S.; Nakanishi, K.; Yanagida, T. Inhibition of cyclin A gene expression in human B cells by an immunosuppressant mizoribine. Clin. Exp. Immunol. 2000, 120, 448–453. [Google Scholar] [PubMed]
- Takahara, S.; Takahashi, K.; Akiyama, T.; Uchida, K.; Tanabe, K.; Amada, N.; Toma, H. Randomized comparative trial of mizoribine versus mycophenolate mofetil in combination with tacrolimus for living donor renal transplantation. Clin. Exp. Nephrol. 2013, 17, 899–904. [Google Scholar] [CrossRef]
- Xing, S.; Yang, J.; Zhang, X.; Zhou, P. Comparative efficacy and safety of mizoribine with mycophenolate mofetil for Asian renal transplantation-a meta-analysis. Clin. Biochem. 2014, 7, 663–669. [Google Scholar] [CrossRef]
- Teh, C.E.; Daley, S.R.; Enders, A.; Goodnow, C.C. T-cell regulation by casitas B-lineage lymphoma (Cblb) is a critical failsafe against autoimmune disease due to autoimmune regulator (Aire) deficiency. Proc. Natl. Acad. Sci. USA 2010, 107, 14709–14714. [Google Scholar] [CrossRef]
- Pfister, G.; Stroh, C.M.; Perschinka, H.; Kind, M.; Knoflach, M.; Hinterdorfer, P.; Wick, G. Detection of HSP60 on the membrane surface of stressed human endothelial cells by atomic force and confocal microscopy. J. Cell Sci. 2005, 118, 1587–1594. [Google Scholar] [CrossRef] [PubMed]
- Pockley, A.G.; De Faire, U.; Kiessling, R.; Lemne, C.; Thulin, T.; Frostegård, J. Circulating heat shock protein and heat shock protein antibody levels in established hypertension. J. Hypertens. 2002, 20, 1815–1820. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Q.; Mandal, K.; Schett, G.; Mayr, M.; Wick, G.; Oberhollenzer, F.; Willeit, J.; Kiechl, S.; Xu, Q. Association of serum-soluble heat shock protein 60 with carotid atherosclerosis. Clinical significance determined in a follow-up study. Stroke 2005, 36, 2571–2576. [Google Scholar] [CrossRef]
- O’Brien, R.L.; Fu, Y.X.; Cranfill, R.; Dallas, A.; Ellis, C.; Reardon, C.; Lang, J.; Carding, S.R.; Kubo, R.; Born, W. Heat shock protein Hsp60-reactive gamma delta cells: A large, diversified T-lymphocyte subset with highly focused specificity. Proc. Natl. Acad. Sci. USA 1992, 89, 4348–4352. [Google Scholar] [CrossRef]
- Morton, H.; McKay, D.A.; Murphy, R.M.; Somodevilla-Torres, M.J.; Swanson, C.E.; Cassady, A.; Summers, K.; Cavanagh, A. Production of a recombinant form of early pregnancy factor that can prolong allogeneic skin graft survival time in rats. Immunol. Cell Biol. 2000, 78, 603–607. [Google Scholar]
- Zhang, B.; Walsh, M.D.; Nguyen, K.B.; Hillyard, N.C.; Cavanagh, A.C.; McCombe, P.A.; Morton, H. Early pregnancy factor treatment suppresses the inflammatory response and adhesion molecule expression in the spinal cord of SJL/J mice with experimental autoimmune encephalomyelitis and the delayed-type hypersensitivity reaction to trinitrochlorobenzene in normal BALB/c mice. J. Neurol. Sci. 2003, 212, 37–46. [Google Scholar]
- Srivastava, S.; Rasool, M. Underpinning IL-6 biology and emphasizing selective JAK blockade as the potential alternate therapeutic intervention for rheumatoid arthritis. Life Sci. 2022, 298, 120516. [Google Scholar] [CrossRef]
- Favalli, E.G. Understanding the Role of Interleukin-6 (IL-6) in the Joint and Beyond: A Comprehensive Review of IL-6 Inhibition for the Management of Rheumatoid Arthritis. Rheumatol. Ther. 2020, 7, 473–516. [Google Scholar] [CrossRef] [PubMed]
- Kaneko, K.; Nagaoka, R.; Ohtomo, Y.; Shimizu, T.; Yamashiro, Y. Effect of mizoribine on IL-6 release by peripheral blood mononuclear cells. Nephron 2002, 91, 506–508. [Google Scholar] [CrossRef] [PubMed]
- Sugiyama, E.; Ikemoto, M.; Taki, H.; Maruyama, M.; Yamashita, N.; Kobayashi, M. Mizoribine, an inhibitor of inosine monophosphate dehydrogenase, inhibits interleukin-6 production by freshly prepared rheumatoid synovial cells. Mod. Rheumatol. 2001, 11, 28–33. [Google Scholar] [CrossRef] [PubMed]
- Stypinski, D.; Obaidi, M.; Combs, M.; Weber, M.; Stewart, A.J.; Ishikawa, H. Safety, tolerability, and pharmacokinetics of higher-dose mizoribine in healthy male volunteers. Br. J. Clin. Pharmacol. 2007, 63, 459–468. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Hosaka, Y.; Itoh, K.; Matsutani, S.; Kawate, S.; Miura, A.; Mizoura, Y.; Yamada, S.; Konno, H.; Grave, E.; Nagata, K.; et al. Fermented food Tempeh induces interleukin 12 and enhances macrophage phagocytosis. J. Food Biochem. 2021, 45, e13958. [Google Scholar] [CrossRef] [PubMed]
- Towbin, H.; Staechelin, T.; Gordon, J. Electrophoretic transfer of proteins from polyacrylamide gels to nitrocellulose sheets: Procedure and some applications. Proc. Natl. Acad. Sci. USA 1979, 76, 4350–4354. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miura, A.; Narita, Y.; Sugawara, T.; Shimizu, H.; Itoh, H. Mizoribine Promotes Molecular Chaperone HSP60/HSP10 Complex Formation. Int. J. Mol. Sci. 2024, 25, 6452. https://doi.org/10.3390/ijms25126452
Miura A, Narita Y, Sugawara T, Shimizu H, Itoh H. Mizoribine Promotes Molecular Chaperone HSP60/HSP10 Complex Formation. International Journal of Molecular Sciences. 2024; 25(12):6452. https://doi.org/10.3390/ijms25126452
Chicago/Turabian StyleMiura, Atsuko, Yukihiko Narita, Taku Sugawara, Hiroaki Shimizu, and Hideaki Itoh. 2024. "Mizoribine Promotes Molecular Chaperone HSP60/HSP10 Complex Formation" International Journal of Molecular Sciences 25, no. 12: 6452. https://doi.org/10.3390/ijms25126452
APA StyleMiura, A., Narita, Y., Sugawara, T., Shimizu, H., & Itoh, H. (2024). Mizoribine Promotes Molecular Chaperone HSP60/HSP10 Complex Formation. International Journal of Molecular Sciences, 25(12), 6452. https://doi.org/10.3390/ijms25126452