
The Protective Role of Magnoliae Flos in Preventing Ovotoxicity and Managing Ovarian Function: An In Vitro and In Vivo Study

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
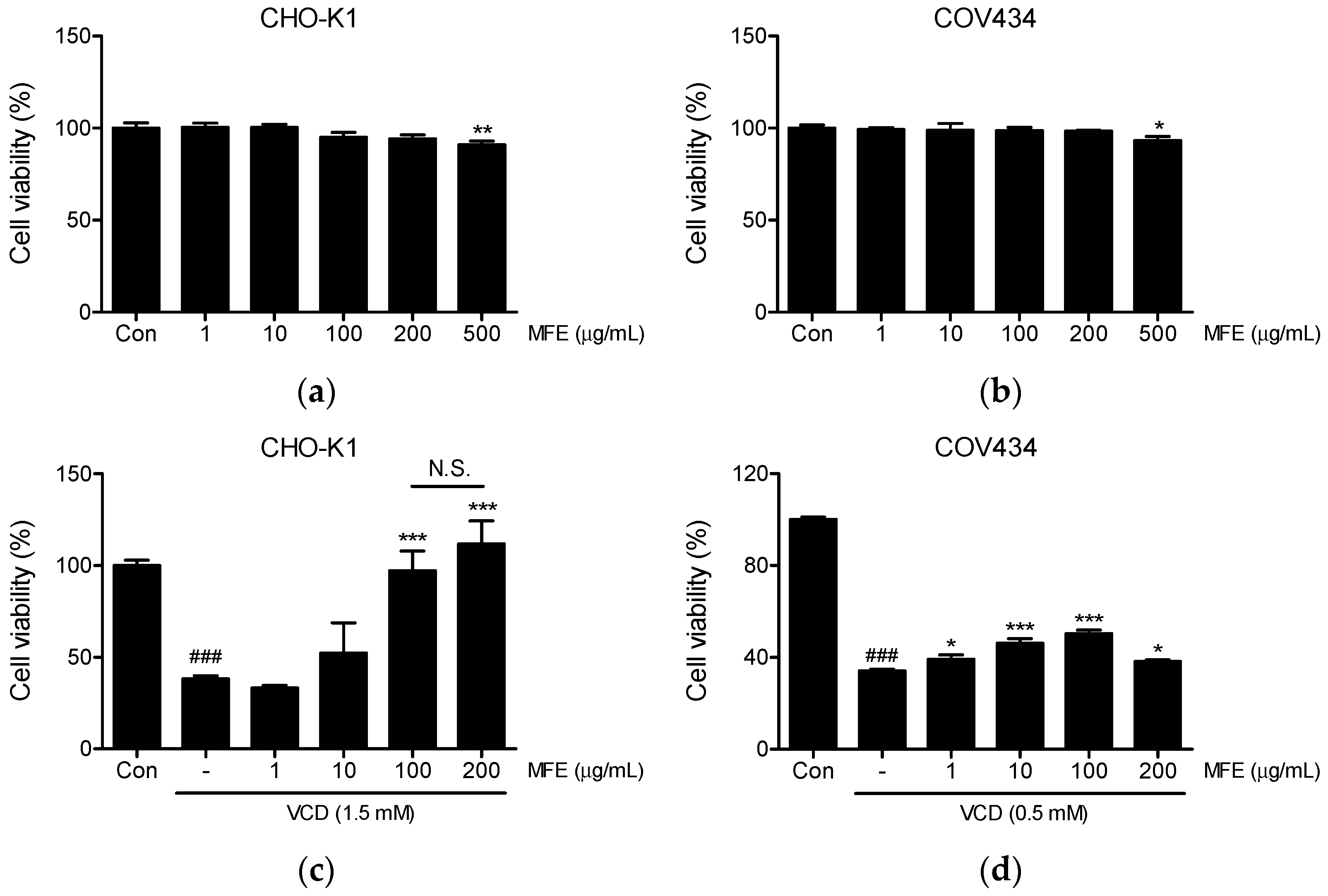
2.1. Effects of MFE on the Viability of CHO-K1 and COV434 Cells
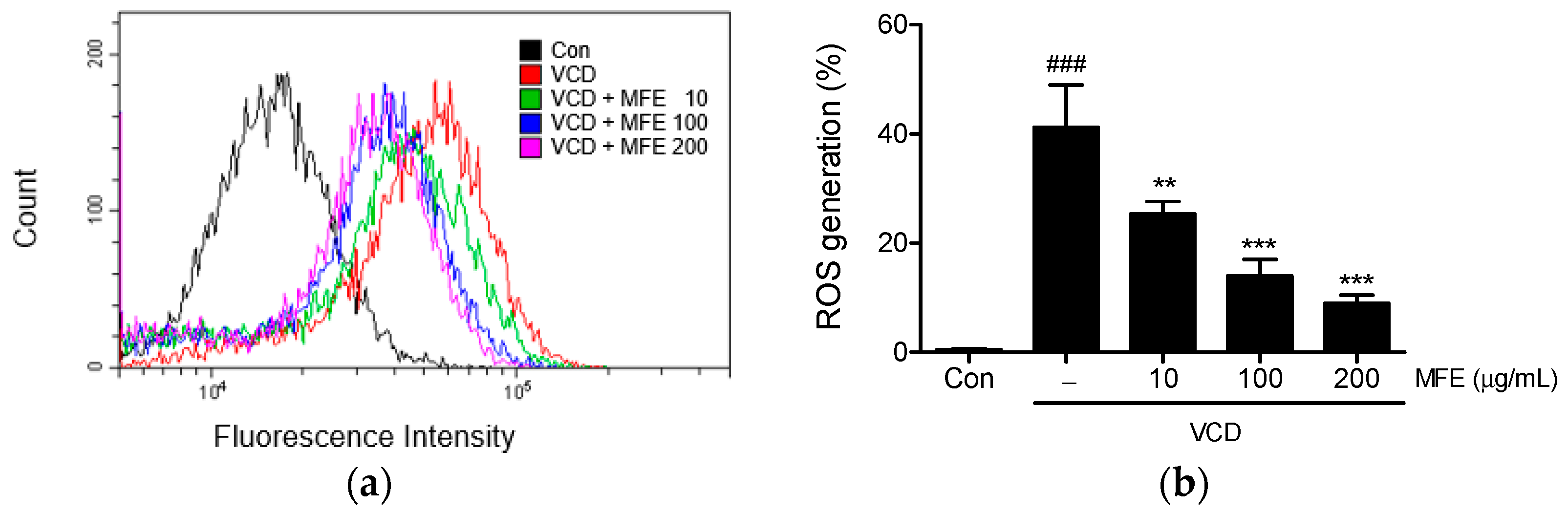
2.2. Inhibitory Effects of MFE on VCD-Induced Reactive Oxygen Species (ROS) Generation
2.3. Inhibitory Effects of MFE on VCD-Induced Apoptosis
2.4. Effects of MFE on Akt Signaling Pathway in CHO-K1 Cells
2.5. Effects of MFE Administration on VCD-Induced Ovarian Failure in Mice
2.6. Effects of MFE Administration on Serum Hormone Levels and Hepatoxicity in the VCD-Induced Ovarian Failure Model
3. Discussion
4. Materials and Methods
4.1. Preparation of Magnoliae Flos Macerated Water Extract
4.2. HPLC Analysis of MFE
4.3. Cell Culture
4.4. Cell Viability Assay
4.5. Measurement of ROS
4.6. DAPI Nuclear Staining
4.7. Western Blot Analysis
4.8. Animal Experimental Design
4.9. Histopathological Analysis
4.10. Serum Biochemistry Analysis
4.11. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Hart, R.J. Physiological Aspects of Female Fertility: Role of the Environment, Modern Lifestyle, and Genetics. Physiol. Rev. 2016, 96, 873–909. [Google Scholar] [CrossRef] [PubMed]
- Tesarik, J.; Galan-Lazaro, M.; Mendoza-Tesarik, R. Ovarian Aging: Molecular Mechanisms and Medical Management. Int. J. Mol. Sci. 2021, 22, 1371. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Zhao, Q.; Li, Y.; Zheng, Z.; Kong, X.; Shu, C.; Liu, Y.; Shi, Y. The role of oxidative stress in ovarian aging: A review. J. Ovarian Res. 2022, 15, 100. [Google Scholar] [CrossRef] [PubMed]
- Bahrehbar, K.; Rezazadeh Valojerdi, M.; Esfandiari, F.; Fathi, R.; Hassani, S.N.; Baharvand, H. Human embryonic stem cell-derived mesenchymal stem cells improved premature ovarian failure. World J. Stem Cells 2020, 12, 857–878. [Google Scholar] [CrossRef] [PubMed]
- Health Insurance Review & Assessment Service. HIRA Bigdata Open Portal. Available online: https://opendata.hira.or.kr/op/opc/olap4thDsInfoTab3.do (accessed on 20 November 2023).
- Ryu, J.H. Menopause Is Coming Even for Those in Their 20s and 30s Who Are Not Married. Prepare by Testing Your Ovarian Age. The JoongAng 07.22.2023, 2023. Available online: https://www.joongang.co.kr/article/25179227#home (accessed on 5 June 2024).
- Jankowska, K. Premature ovarian failure. Prz. Menopauzalny 2017, 16, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Wesevich, V.; Kellen, A.N.; Pal, L. Recent advances in understanding primary ovarian insufficiency. F1000Research 2020, 9, 1101. [Google Scholar] [CrossRef] [PubMed]
- Chon, S.J.; Umair, Z.; Yoon, M.S. Premature Ovarian Insufficiency: Past, Present, and Future. Front. Cell Dev. Biol. 2021, 9, 672890. [Google Scholar] [CrossRef] [PubMed]
- Podfigurna-Stopa, A.; Czyzyk, A.; Grymowicz, M.; Smolarczyk, R.; Katulski, K.; Czajkowski, K.; Meczekalski, B. Premature ovarian insufficiency: The context of long-term effects. J. Endocrinol. Investig. 2016, 39, 983–990. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.Y.; Chen, S.R.; Chen, J.M.; Shi, Q.Y.; Lin, S. Therapeutic options for premature ovarian insufficiency: An updated review. Reprod. Biol. Endocrinol. 2022, 20, 28. [Google Scholar] [CrossRef]
- Lambrinoudaki, I.; Paschou, S.A.; Lumsden, M.A.; Faubion, S.; Makrakis, E.; Kalantaridou, S.; Panay, N. Premature ovarian insufficiency: A toolkit for the primary care physician. Maturitas 2021, 147, 53–63. [Google Scholar] [CrossRef]
- Dragojevic-Dikic, S.; Rakic, S.; Nikolic, B.; Popovac, S. Hormone replacement therapy and successful pregnancy in a patient with premature ovarian failure. Gynecol. Endocrinol. 2009, 25, 769–772. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Kim, M.S.; Kim, P.W.; Kim, S. Korean medicine treatment for premature ovarian failure: Three case reports. Explore 2023, 19, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Nam, E.Y.; Kim, S.A.; Kim, H.; Kim, S.H.; Han, J.H.; Lee, J.H.; Kim, D.I. Akt activation by Evodiae Fructus extract protects ovary against 4-vinylcyclohexene diepoxide-induced ovotoxicity. J. Ethnopharmacol. 2016, 194, 733–739. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Wu, S.; Cao, D.; Li, M.; Liu, J.; Nie, G.; Li, Y.; Yang, H. Huyang yangkun formula protects against 4-Vinylcyclohexene diepoxide-induced premature ovarian insufficiency in rats via the Hippo-JAK2/STAT3 signaling pathway. Biomed. Pharmacother. 2019, 116, 109008. [Google Scholar] [CrossRef] [PubMed]
- Su, X.; Wang, X.; Liu, Y.; Kong, W.; Yan, F.; Han, F.; Liu, Q.; Shi, Y. Effect of Jiajian Guishen Formula on the senescence-associated heterochromatic foci in mouse ovaria after induction of premature ovarian aging by the endocrine-disrupting agent 4-vinylcyclohexene diepoxide. J. Ethnopharmacol. 2021, 269, 113720. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Song, Y.; Liu, X.; Zhang, C.; Li, F.; Hu, R.; Huang, Y.; Ma, W.; Song, K.; Zhang, M. Si-Wu-Tang facilitates ovarian function through improving ovarian microenvironment and angiogenesis in a mouse model of premature ovarian failure. J. Ethnopharmacol. 2021, 280, 114431. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Li, M.; Liu, J.; Nie, G.; Li, Y.; Yang, H. Protective effect of Huyang Yangkun Formula on ovarian function in premature ovarian insufficiency rats based on apoptotic mechanism. J. Ethnopharmacol. 2021, 280, 114477. [Google Scholar] [CrossRef]
- Dou, X.; Jin, X.; Chen, X.; Zhou, Q.; Chen, H.; Wen, M.; Chen, W. Bu-Shen-Ning-Xin decoction alleviates premature ovarian insufficiency (POI) by regulating autophagy of granule cells through activating PI3K/AKT/mTOR pathway. Gynecol. Endocrinol. 2022, 38, 754–764. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Li, C.G.; Zhou, S.F.; Pang, E.C.; Story, D.F.; Xue, C.C. Chemistry and bioactivity of Flos Magnoliae, a Chinese herb for rhinitis and sinusitis. Curr. Med. Chem. 2008, 15, 1616–1627. [Google Scholar] [CrossRef]
- Minami, M.; Konishi, T.; Jiang, Z.; Arai, T.; Makino, T. Effect of Shin’iseihaito on murine allergic reaction induced by nasal sensitization. J. Tradit. Complement. Med. 2016, 6, 252–256. [Google Scholar] [CrossRef]
- Kim, H.M.; Yi, J.M.; Lim, K.S. Magnoliae flos inhibits mast cell-dependent immediate-type allergic reactions. Pharmacol. Res. 1999, 39, 107–111. [Google Scholar] [CrossRef] [PubMed]
- Ham, J.R.; Yun, K.W.; Lee, M.K. Anti-Inflammatory and Antioxidant in Vitro Activities of Magnoliae Flos Ethanol Extract. Prev. Nutr. Food Sci. 2021, 26, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Gil, T.Y.; Jin, B.R.; Cha, Y.Y.; An, H.J. Magnoliae flos Downregulated Lipopolysaccharide-Induced Inflammatory Responses via NF-kappaB/ERK-JNK MAPK/STAT3 Pathways. Mediators Inflamm. 2022, 2022, 6281892. [Google Scholar] [CrossRef] [PubMed]
- Jung, Y.S.; Weon, J.B.; Yang, W.S.; Ryu, G.; Ma, C.J. Neuroprotective effects of Magnoliae Flos extract in mouse hippocampal neuronal cells. Sci. Rep. 2018, 8, 9693. [Google Scholar] [CrossRef] [PubMed]
- Jun, A.Y.; Kim, H.J.; Park, K.K.; Son, K.H.; Lee, D.H.; Woo, M.H.; Kim, Y.S.; Lee, S.K.; Chung, W.Y. Extract of Magnoliae Flos inhibits ovariectomy-induced osteoporosis by blocking osteoclastogenesis and reducing osteoclast-mediated bone resorption. Fitoterapia 2012, 83, 1523–1531. [Google Scholar] [CrossRef]
- Kimura, M.; Suzuki, J.; Yamada, T.; Yoshizaki, M.; Kikuchi, T.; Kadota, S.; Matsuda, S. Anti-Inflammatory Effect of Neolignans Newly Isolated from the Crude Drug “Shin-I” (Flos Magnoliae). Planta Med. 1985, 51, 291–293. [Google Scholar] [CrossRef] [PubMed]
- Ma, P.; Che, D.; Zhao, T.; Zhang, Y.; Li, C.; An, H.; Zhang, T.; He, H. Magnolin inhibits IgE/Ag-induced allergy in vivo and in vitro. Int. Immunopharmacol. 2019, 76, 105867. [Google Scholar] [CrossRef] [PubMed]
- Phan, H.T.L.; Nam, Y.R.; Kim, H.J.; Woo, J.H.; NamKung, W.; Nam, J.H.; Kim, W.K. In-vitro and in-vivo anti-allergic effects of magnolol on allergic rhinitis via inhibition of ORAI1 and ANO1 channels. J. Ethnopharmacol. 2022, 289, 115061. [Google Scholar] [CrossRef] [PubMed]
- Hong, P.T.L.; Kim, H.J.; Kim, W.K.; Nam, J.H. Flos magnoliae constituent fargesin has an anti-allergic effect via ORAI1 channel inhibition. Korean J. Physiol. Pharmacol. 2021, 25, 251–258. [Google Scholar] [CrossRef]
- Kong, C.S.; Lee, J.I.; Kim, J.A.; Seo, Y. In vitro evaluation on the antiobesity effect of lignans from the flower buds of Magnolia denudata. J. Agric. Food Chem. 2011, 59, 5665–5670. [Google Scholar] [CrossRef]
- Hu, M.; Bai, M.; Ye, W.; Wang, Y.; Wu, H. Variations in Volatile Oil Yield and Composition of “Xin-yi” (Magnolia biondii Pamp. Flower Buds) at Different Growth Stages. J. Oleo Sci. 2018, 67, 779–787. [Google Scholar] [CrossRef] [PubMed]
- Kappeler, C.J.; Hoyer, P.B. 4-vinylcyclohexene diepoxide: A model chemical for ovotoxicity. Syst. Biol. Reprod. Med. 2012, 58, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Cho, O.H.; Hwang, K.H. Comparison of quality of life and related factors according to premature menopause using the Korea National Health and Nutrition Examination Survey (2014–2017). Korean J. Women Health Nurs. 2020, 26, 131–140. [Google Scholar] [CrossRef] [PubMed]
- Desmeules, P.; Devine, P.J. Characterizing the ovotoxicity of cyclophosphamide metabolites on cultured mouse ovaries. Toxicol. Sci. 2006, 90, 500–509. [Google Scholar] [CrossRef]
- Mentese, A.; Demir, S.; Kucuk, H.; Yulug, E.; Alemdar, N.T.; Demir, E.A.; Aliyazicioglu, Y. Vanillic acid abrogates cisplatin-induced ovotoxicity through activating Nrf2 pathway. Tissue Cell 2023, 84, 102161. [Google Scholar] [CrossRef]
- Takizawa, K.; Yagi, H.; Jerina, D.M.; Mattison, D.R. Murine strain differences in ovotoxicity following intraovarian injection with benzo(a)pyrene, (+)-(7R,8S)-oxide, (−)-(7R,8R)-dihydrodiol, or (+)-(7R,8S)-diol-(9S,10R)-epoxide-2. Cancer Res. 1984, 44, 2571–2576. [Google Scholar] [PubMed]
- Igawa, Y.; Keating, A.F.; Rajapaksa, K.S.; Sipes, I.G.; Hoyer, P.B. Evaluation of ovotoxicity induced by 7, 12-dimethylbenz[a]anthracene and its 3,4-diol metabolite utilizing a rat in vitro ovarian culture system. Toxicol. Appl. Pharmacol. 2009, 234, 361–369. [Google Scholar] [CrossRef]
- Hu, X.; Roberts, J.R.; Apopa, P.L.; Kan, Y.W.; Ma, Q. Accelerated ovarian failure induced by 4-vinyl cyclohexene diepoxide in Nrf2 null mice. Mol. Cell. Biol. 2006, 26, 940–954. [Google Scholar] [CrossRef]
- Koebele, S.V.; Bimonte-Nelson, H.A. Modeling menopause: The utility of rodents in translational behavioral endocrinology research. Maturitas 2016, 87, 5–17. [Google Scholar] [CrossRef]
- Song, G.; Ouyang, G.; Bao, S. The activation of Akt/PKB signaling pathway and cell survival. J. Cell. Mol. Med. 2005, 9, 59–71. [Google Scholar] [CrossRef]
- Reddy, P.; Shen, L.; Ren, C.; Boman, K.; Lundin, E.; Ottander, U.; Lindgren, P.; Liu, Y.X.; Sun, Q.Y.; Liu, K. Activation of Akt (PKB) and suppression of FKHRL1 in mouse and rat oocytes by stem cell factor during follicular activation and development. Dev. Biol. 2005, 281, 160–170. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Rajareddy, S.; Liu, L.; Jagarlamudi, K.; Boman, K.; Selstam, G.; Reddy, P. Control of mammalian oocyte growth and early follicular development by the oocyte PI3 kinase pathway: New roles for an old timer. Dev. Biol. 2006, 299, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Mayer, L.P.; Pearsall, N.A.; Christian, P.J.; Devine, P.J.; Payne, C.M.; McCuskey, M.K.; Marion, S.L.; Sipes, I.G.; Hoyer, P.B. Long-term effects of ovarian follicular depletion in rats by 4-vinylcyclohexene diepoxide. Reprod. Toxicol. 2002, 16, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Sahambi, S.K.; Visser, J.A.; Themmen, A.P.; Mayer, L.P.; Devine, P.J. Correlation of serum anti-Mullerian hormone with accelerated follicle loss following 4-vinylcyclohexene diepoxide-induced follicle loss in mice. Reprod. Toxicol. 2008, 26, 116–122. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, F.S.; Goode, A.K.; Kock, N.D.; Arifin, E.A.; Cline, J.M.; Adams, M.R.; Hoyer, P.B.; Christian, P.J.; Isom, S.; Kaplan, J.R.; et al. Effects of 4-vinylcyclohexene diepoxide on peripubertal and adult Sprague-Dawley rats: Ovarian, clinical, and pathologic outcomes. Comp. Med. 2009, 59, 46–59. [Google Scholar] [PubMed]
- Nelson, L.R.; Bulun, S.E. Estrogen production and action. J. Am. Acad. Dermatol. 2001, 45, S116–S124. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, D.; Griswold, J.; Erman, M.; Pangborn, W. Structural basis for androgen specificity and oestrogen synthesis in human aromatase. Nature 2009, 457, 219–223. [Google Scholar] [CrossRef]
- Sharma, N.; Tiwari, N.; Vyas, M.; Khurana, N.; Muthuraman, A.; Utreja, P. An overview of therapeutic effects of vanillic acid. Plant Arch. 2020, 20, 3053–3059. [Google Scholar]
- Zhou, Y.; Wang, F.; Hao, L.; Wang, N. Effects of magnoline on P-selectin’s expression in diabetic rats and its reno-protection. Kidney Blood Press. Res. 2013, 37, 211–220. [Google Scholar] [CrossRef]
- Wang, F.; Zhang, G.; Zhou, Y.; Gui, D.; Li, J.; Xing, T.; Wang, N. Magnolin protects against contrast-induced nephropathy in rats via antioxidation and antiapoptosis. Oxid. Med. Cell. Longev. 2014, 2014, 203458. [Google Scholar] [CrossRef]
- Castillo, C.; Bravo-Arrepol, G.; Wendt, A.; Saez-Orellana, F.; Millar, C.; Burgos, C.F.; Gavilan, J.; Pacheco, C.; Ahumada-Rudolph, R.; Napiorkowska, M.; et al. Neuroprotective Properties of Eudesmin on a Cellular Model of Amyloid-beta Peptide Toxicity. J. Alzheimers Dis. 2023, 94, S97–S108. [Google Scholar] [CrossRef] [PubMed]
- Park, J.-H.; Ahn, E.-K.; Park, K.; Hwang, M.H.; Cho, Y.-R.; Lee, D.-R.; Choi, B.-K.; Oh, J.S. 4-week dose range finding toxicity test of Magnoliae Flos extract in Sprague-Dawley rats. J. Appl. Biol. Chem. 2022, 65, 43–48. [Google Scholar] [CrossRef]
- Jeon, S.K.; Park, S.M.; Ahn, J.Y.; Kim, O.H.; Kim, Y.W.; Park, K.-I.; Park, S.-D.; Lee, J.-H. Anticancer activity of herbal formula Jisilhaebaekgyeji-Tang against human breast cancer cells and its mechanism. Mol. Cell. Toxicol. 2022, 19, 437–451. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, M.R.; Kim, D.-I.; Park, S.Y.; Kang, H.J.; Park, S.-D.; Lee, J.-H. The Protective Role of Magnoliae Flos in Preventing Ovotoxicity and Managing Ovarian Function: An In Vitro and In Vivo Study. Int. J. Mol. Sci. 2024, 25, 6456. https://doi.org/10.3390/ijms25126456
Kim MR, Kim D-I, Park SY, Kang HJ, Park S-D, Lee J-H. The Protective Role of Magnoliae Flos in Preventing Ovotoxicity and Managing Ovarian Function: An In Vitro and In Vivo Study. International Journal of Molecular Sciences. 2024; 25(12):6456. https://doi.org/10.3390/ijms25126456
Chicago/Turabian StyleKim, Mi Ra, Dong-Il Kim, Sung Yun Park, Hyo Jin Kang, Sun-Dong Park, and Ju-Hee Lee. 2024. "The Protective Role of Magnoliae Flos in Preventing Ovotoxicity and Managing Ovarian Function: An In Vitro and In Vivo Study" International Journal of Molecular Sciences 25, no. 12: 6456. https://doi.org/10.3390/ijms25126456